模擬空運過程中克氏原螯蝦生理生化指標的變化規律
2021-02-09 12:13:26郝晨光李軍濤張澤龍張秀霞冼健安王冬梅
南方農業學報 2021年10期
郝晨光 李軍濤 張澤龍 張秀霞 冼健安 王冬梅
摘要:【目的】明確空運過程中影響克氏原螯蝦存活率的關鍵環境因子,為有針對性地進行空運前預處理提供科學依據。【方法】模擬克氏原螯蝦空運條件,將克氏原螯蝦放入泡沫箱中,加入冰瓶后進行完全密封(密封組)或打孔通氣(打孔組)。模擬空運過程實時監測泡沫箱內的含氧量、溫度和相對濕度;模擬空運18 h后采集克氏原螯蝦的血清、肝胰腺、鰓絲及中腸等組織樣品,分別測定血清溶菌酶(LZM)活性、多酚氧化酶(PPO)活性、超氧化物歧化酶(SOD)活性及丙二醛(MDA)含量,肝胰腺SOD活性、總抗氧化能力(T-AOC)、MDA含量及蛋白質羰基含量,以及鰓絲Na+K+-ATP活性;并制作克氏原螯蝦肝胰腺、鰓絲及中腸組織石蠟切片,統計模擬空運結束后克氏原螯蝦的存活率。【結果】在模擬空運過程中,相對濕度變化不明顯,溫度和含氧量的變化引起克氏原螯蝦應激反應,是影響空運的主要環境因子。經模擬空運后,密封組克氏原螯蝦血清LZM活性、PPO活性及MDA含量顯著升高(P<0.05,下同),SOD活性顯著降低;肝胰腺蛋白質羰基含量、MDA含量和T-AOC水平均顯著高于打孔組及模擬空運前,而SOD活性變化不顯著;密封組和打孔組克氏原螯蝦經模擬空運18 h后,其鰓絲Na+K+-ATP活性均顯著高于模擬空運前。克氏原螯蝦經模擬空運18 h后其存活率較高,但模擬空運后暫養1周發現密封組和打孔組的克氏原螯蝦存活率均從第4 d開始顯著下降,至暫養第7 d密封組克氏原螯蝦的存活率僅為59.33%;且其鰓絲有大量附著物,微血管腔結構已完全消失,大部分呼吸上皮細胞脫落、壞死,鰓膜結構遭到破壞。【結論】溫度和含氧量是影響克氏原螯蝦空運存活率的主要環境因子。模擬空運對克氏原螯蝦造成嚴重損傷且不可逆,在模擬空運結束后克氏原螯蝦因自身缺乏修復能力而大量死亡。因此,實際生產中可通過在運輸前投喂免疫增強劑,或在運輸后優化其暫養環境等方式以提高克氏原螯蝦空運存活率,進而確得其引種成功。
關鍵詞:克氏原螯蝦;模擬空運;環境因子;生理生化指標;存活率
中圖分類號:S981.21? ? ? ? ? ? ? ? ? ? ? 文獻標志碼: A 文章編號:2095-1191(2021)10-2896-08
Abstract: 【Objective】 To identify the key environmental factors affecting Procambarus clarkii survival during air transportation and to provide the scientific basis for targeted pre- and post-transport treatments to improve survival rates. 【Method】 The conditions of air transportation were simulated by placing the crayfish in foam boxes under ice-cold conditions with (perforated group) or without (sealed group) ventilation. Real-time monitoring of oxygen content, temperature and relative humidity in the foam box was performed during simulated air transportation. After 18 h of simulation, the serum, hepatopancreas, gill filament and midgut samples were collected and the serum activities of lysozyme, polyphenol oxidase (PPO), superoxide dismutase (SOD) and malondialdehyde (MDA) were determined. Hepatopancreas SOD activity, total antioxidant capacity (T-AOC), MDA content, protein carbonyl content and gill filament Na+K+ -ATPASE activity were used, together with paraffin sections of the hepatopancreas, gill filaments and midgut, to calculate the survival rate of P. clarkii after simulated air transport. 【Result】 During simulated air transport, the relative humidity was not obviously altered, while the changes in temperature and oxygen content were significantly affected and observed to cause stress reactions in P. clarkii. After the simulation, the activities of lysozyme, PPO and MDA in the serum of the sealed group were significantly increased (P<0.05, the same below), while the activities of SOD were significantly decreased. Hepatopancreas protein carbonyl content, MDA content and T-AOC level were significantly higher than those in the perforated and control groups, while SOD activity showed no significant change. The activity of Na+K+-ATP in gill filaments of both the sealed and perforated groups was significantly higher than that of the control group. After simulated air transport for 18 h, the cray-fish survival rate was higher. However, the survival rate of both the sealing and perforated group decreased significantly from the 4th day, and only 59.33% of the sealed group was alive on the 7th day. Most of the respiratory epithelial cells were exfoliated and necrotic, and the branchial membrane structure was destroyed. 【Conclusion】 The changes of temperature and oxygen content are the main environmental factors affecting the survival rate of P. clarkii during air transportation. The simulated air transport caused serious and irreversible damage to P. clarkii and a large number of P. clarkii died after their return to control conditions. Therefore, the survival rate of air-transported P. clarkii can be improved by feeding immune enhancers before transportation or optimizing the environmental conditions following transport to improve recovery rates.
Key words: Procambarus clarkii; simulated air transportation; environmental factors; biochemical indicators; survival rate
Foundation item:Hainan Key Research and Development Project(ZDEF2018027);Central Public-interest Scientific Institution Basal Research Fund for Chinese Academy of Tropical Agricultural Sciences(1630052019013)
0 引言
【研究意義】克氏原螯蝦(Procambarus clarkii)又名小龍蝦,原產于北美洲,現已發展成為我國淡水蝦類中的重要資源(姜愛蘭等,2020)。長江中下游地區是目前我國克氏原螯蝦養殖的主要集中區,屬于亞熱帶季風性氣候,每年10月天氣轉涼后,克氏原螯蝦開始冬眠,直至翌年3月均無新鮮的活蝦上市。海南省地處熱帶地區,其氣候條件非常適合克氏原螯蝦生長,若能引種成功可有效填補冬春季節克氏原螯蝦市場的空白。長途運輸是生物活體引種的必經環節,其中以空運所需時間最短、運輸應激最小,對引種對象損傷低,是一種理想的引種運輸方法。因此,掌握空運過程中克氏原螯蝦各項生理生化指標的變化規律,對有效提高克氏原螯蝦引種成功率具有重要意義。【前人研究進展】至今,針對甲殼類動物空氣暴露及低氧脅迫的研究已有較多報道,且對離水條件下甲殼類動物體內生理生化指標的變化規律有所了解。Rabin等(1996)發現無水干運會引起斑節對蝦(Penaeus monodon)肝胰腺的氧化應激及損傷,且影響HSP70蛋白和鐵蛋白的表達。Paterson等(1997)研究發現,在將捕撈龍蝦干運至母艦的過程中由于水流量下降會迅速降低溶解氧含量,致使龍蝦血淋巴當中的乳酸濃度也隨之下降。Oliveira等(2001)對三疣梭子蟹(Portunus trituberculatus)進行干露脅迫,結果發現缺氧導致密封組三疣梭子蟹的葡萄糖攝取和糖原合成明顯減少,而肝胰腺糖原動員和血淋巴葡萄糖濃度呈上升趨勢。de Oliveira等(2005)對中華絨螯蟹(Eriocheir sinensis)進行低氧脅迫時發現,缺氧狀態下其體內的過氧化氫酶(CAT)和谷胱甘肽轉移酶(GST)活性增強,而超氧化物歧化酶(SOD)活性降低。Zenteno-Savín等(2006)在凡納濱對蝦(P. vannamei)的低氧脅迫研究中發現,缺氧后凡納濱對蝦肝胰腺的脂質過氧化(TBARS)水平高于鰓絲和肌肉,且缺氧恢復會造成其抗氧化能力降低。姜娜等(2014)在三疣梭子蟹的干露脅迫研究中發現,干露脅迫時間對其SOD活性和丙二醛(MDA)含量均無顯著影響,但顯著影響肝胰臟總抗氧化能力(T-AOC)。段亞飛等(2015)研究表明,經干露脅迫后日本囊對蝦(P. japonicus)的谷胱甘肽過氧化物酶(GPx)活性、CAT活性、SOD活性及MDA含量變化顯著,可作為日本囊對蝦干露脅迫的監測指標。Lu等(2016)在中華絨螯蟹模擬干運過程中發現,干露的中華絨螯蟹在受熱應激刺激后,其血淋巴葡萄糖和脂質水平均發生顯著變化。連春盈等(2017)通過對脊尾白蝦(Exopalaemon carinicauda Holthuis)進行干露脅迫,結果發現密封組脊尾白蝦肝胰腺、鰓絲、肌肉的琥珀酸脫氫酶(SDH)和細胞色素氧化酶(CCO)活性均隨脅迫時間的延長而降低,而延胡索酸還原酶(FRD)和乳酸脫氫酶(LDH)活性隨脅迫時間的延長呈先升高后降低的變化趨勢。王海鋒等(2019)在對克氏原螯蝦進行干露脅迫后發現,密封組克氏原螯蝦的T-AOC無顯著變化,但CAT活性隨脅迫時間的延長逐漸升高,SOD活性顯著降低,MDA含量在干露脅迫第24 h達峰值。【本研究切入點】目前,關于水產動物長途運輸的研究以魚類較多,蝦蟹類的研究相對較少,要確保存活率必須綜合考慮各項生理生化指標參數,包括應激程度及健康狀況等因素(Pan et al.,2007;吳佳靜等,2016)。克氏原螯蝦巨大的市場需求促使其養殖面積不斷擴大,而克氏原螯蝦引種需通過無水空運來實現,因此探究克氏原螯蝦在空運過程中各項生理生化指標的變化規律及運輸后的存活率,對有效優化運輸環境參數具有重要意義。【擬解決的關鍵問題】通過監測模擬空運過程中的各項環境因子,以及運輸前后克氏原螯蝦各項生理生化指標的變化,明確空運過程中影響克氏原螯蝦存活率的關鍵環境因子,為有針對性進行空運前預處理提供科學依據。
1 材料與方法
1. 1 模擬空運
在海南省文昌市苑頭鎮克氏原螯蝦養殖基地,選取規格相近、活力較好的克氏原螯蝦(平均體重24.0±0.5 g)。將克氏原螯蝦分別裝入6個泡沫箱中,每箱100只,泡沫箱內提前放好冰瓶。其中,試驗組3個泡沫箱完全密封,打孔組3個泡沫箱打孔透氣。泡沫箱中插入氧氣測定儀和相對濕度測定儀,進行環境因子實時監測。將泡沫箱置于室內(提前打開空調使室內溫度保持在24 ℃,模擬空運貨倉環境)進行18 h模擬空運。
1. 2 樣品采集
模擬空運前進行0 h取樣,以無菌注射器吸入抗凝劑后抽取克氏原螯蝦血淋巴液,注入1.5 mL無菌離心管中,2500 r/min離心10 min,收集血清置于新的無菌離心管中并迅速放入液氮,-80 ℃保存備用。同時用無菌手術剪摘取肝胰腺、鰓絲及中腸等組織,裝入1.5 mL無菌離心管中放入液氮,-80 ℃保存;取部分肝胰腺、鰓絲和中腸組織放入甲醛固定液中固定,用于制作石蠟切片。模擬空運結束后,打開泡沫箱,統計克氏原螯蝦存活率并進行取樣(同0 h取樣),將剩余克氏原螯蝦放入玻璃缸暫養,統計暫養1周后的存活率。
1. 3 測定指標及方法
通過手機實時傳送溫度、相對濕度及含氧量隨時間的變化情況,試驗結束后導出數據并繪制折線圖。采用南京建成生物工程研究所生產的溶菌酶(LZM)測試盒(A050-1-1)、多酚氧化酶(PPO)測試盒(A136-1-1)、總超氧化物歧化酶(SOD)試劑盒(A001-3-2)、總抗氧化能力(T-AOC)檢測試劑盒(比色法,A015-1-2)、微量丙二醛(MDA)測定試劑盒(TBA法,A003-2-2)、蛋白質羰基含量測定試劑盒(紫外比色法,A087-1-2)、超微量Na+K+-ATP酶試劑盒(A070-2-2)分別測定克氏原螯蝦血清LZM活性、PPO活性、SOD活性及MDA含量,肝胰腺SOD活性、T-AOC水平、MDA含量及蛋白質羰基含量,以及鰓絲Na+K+-ATP活性。同時,參照溫久福等(2019)的方法制作克氏原螯蝦肝胰腺、鰓絲及中腸組織石蠟切片,在顯微鏡下進行觀察和拍照。所有試驗數據以SPSS 18.0進行整理及單因素方差分析(One-way ANVOA)。
2 結果與分析
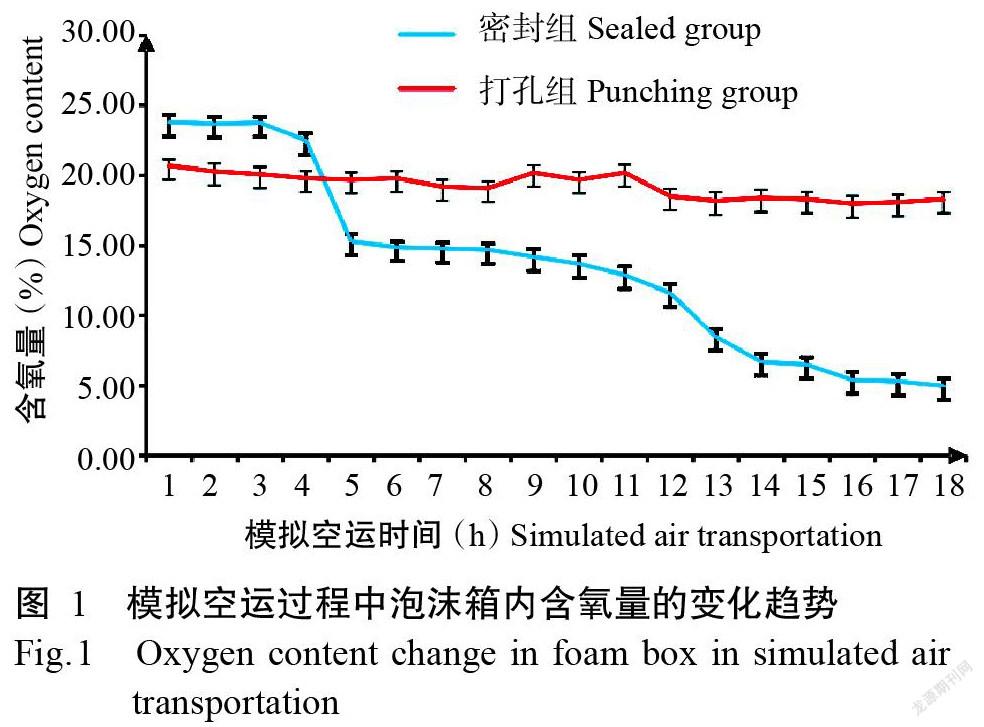
2. 1 模擬空運過程中泡沫箱內含氧量的變化趨勢
由圖1可知,密封組和打孔組泡沫箱內的含氧量均隨模擬空運時間的延長而下降。密封組由于完全密封,含氧量下降速率明顯高于打孔組;至模擬空運結束后,密封組泡沫箱內的含氧量為5.00%,遠低于打孔組的18.30%。由此可見,密封環境很容易造成運輸過程中克氏原螯蝦缺氧。
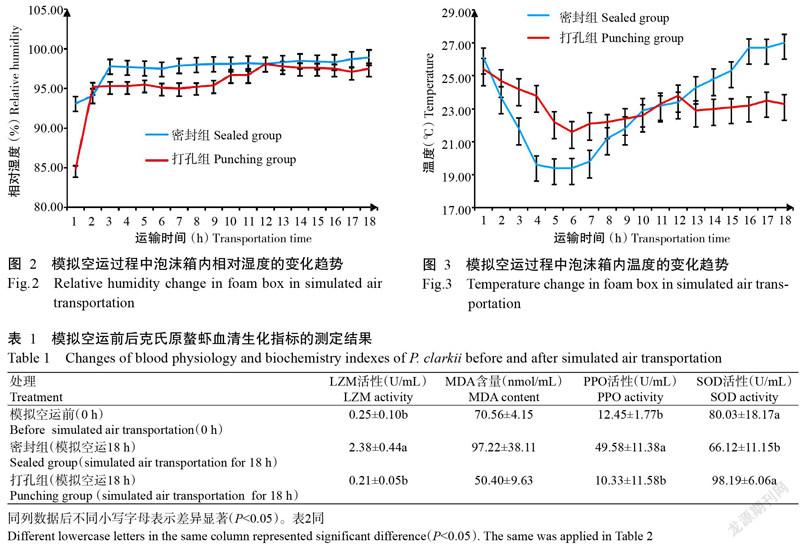
2. 2 模擬空運過程中泡沫箱內相對濕度的變化趨勢
由圖2可知,密封組和打孔組泡沫箱內的相對濕度均隨模擬空運時間的延長而上升,且最終的相對濕度達到100.00%左右,二者差異不明顯。相對濕度上升可能是由于冰瓶的蒸發及克氏原螯蝦的呼吸作用所致。
2. 3 模擬空運過程中泡沫箱內溫度的變化趨勢
由圖3可知,密封組和打孔組泡沫箱內的溫度均隨模擬空運時間的延長呈先下降后上升的變化趨勢,可能與泡沫箱內放置冰瓶有關。前期冰瓶起到降溫的作用,但隨著模擬空運時間的推移,冰瓶的降溫作用逐漸不明顯,而導致泡沫箱內溫度升高。密封組泡沫箱屬于密封環境,其溫度變化較劇烈,運輸期間溫差達7.6 ℃;而打孔組由于打孔透氣,受外界環境影響,其總體溫差相對不明顯,僅為3.4 ℃。
2. 4 模擬空運對克氏原螯蝦各項生物酶活性的影響
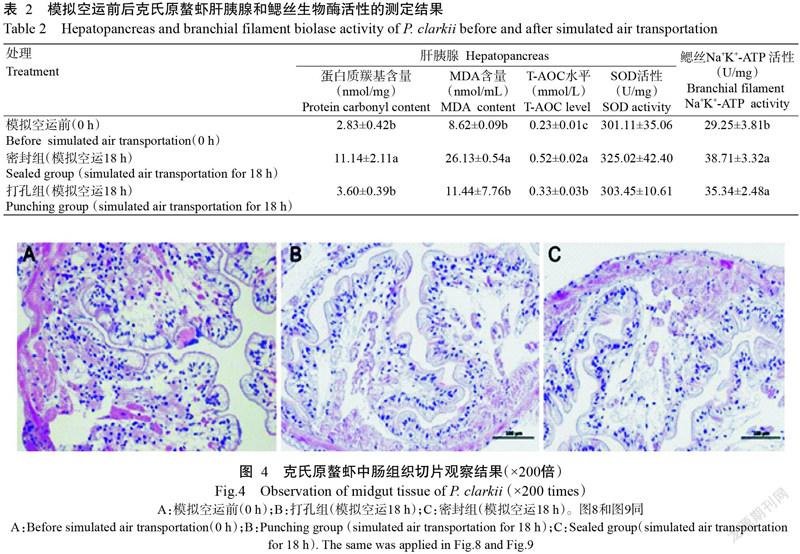
模擬空運結束后,密封組克氏原螯蝦血清LZM活性顯著高于打孔組及模擬空運前(P<0.05,下同),而打孔組克氏原螯蝦血清LZM活性與模擬空運前相比差異不顯著(P>0.05,下同);在血清MDA含量方面,模擬空運前后及密封組與打孔組間的差異均不顯著,但以密封組克氏原螯蝦的含量最高;密封組克氏原螯蝦血清PPO活性顯著高于打孔組及模擬空運前,但其SOD活性顯著低于打孔組及模擬空運前;打孔組克氏原螯蝦血清PPO活性和SOD活性與模擬空運前相比,其差異均不顯著(表1)。
模擬空運結束后,密封組克氏原螯蝦肝胰腺蛋白質羰基含量和MDA含量均顯著高于打孔組及模擬空運前,而打孔組與模擬空運前相比,差異均不顯著(表2);密封組克氏原螯蝦肝胰腺T-AOC水平顯著高于打孔組,且二者又顯著高于模擬空運前;克氏原螯蝦肝胰腺SOD活性在模擬空運前后的差異不顯著,且密封組與打孔組間的差異也不顯著。密封組和打孔組克氏原螯蝦經模擬空運后,其鰓絲Na+K+-ATP活性均顯著高于模擬空運前,但二者差異不顯著。
2. 5 模擬空運對克氏原螯蝦器官組織結構的影響
2. 5. 1 中腸組織切片觀察結果 模擬空運前克氏原螯蝦中腸組織上皮層、基膜、結締組織層、肌肉層和外膜結構完整,上皮被覆單層矮柱狀上皮,排列整齊,部分細胞染色較淺;疏松結締組織層未見明顯的細胞壞死和血細胞滲出;基膜、肌層和外膜未見明顯病理變化(圖4-A)。打孔組克氏原螯蝦中腸組織上皮層、基膜、結締組織層、肌肉層和外膜結構較完整,上皮被覆單層矮柱狀上皮,排列較整齊,部分細胞空泡變性,細胞腫大,排列不整齊,總體受損程度低于密封組克氏原螯蝦;結締組織層、基膜、肌層和外膜未見明顯病理變化(圖4-B)。密封組克氏原螯蝦中腸組織上皮層、基膜、結締組織層、肌肉層和外膜結構較完整,上皮被覆單層矮柱狀上皮,部分細胞脫落,略顯腫脹,排列紊亂;結締組織層、基膜、肌層和外膜也未見明顯壞死及血細胞滲出等病理變化(圖4-C)。
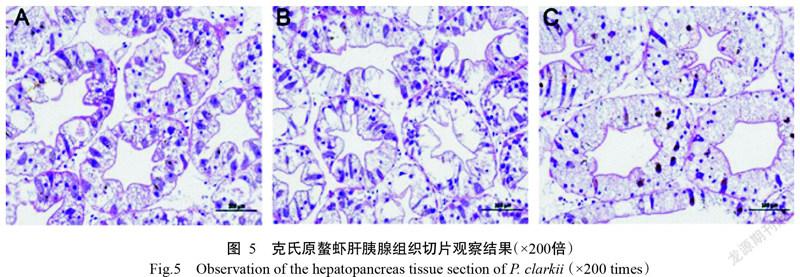
2. 5. 2 肝胰腺組織切片觀察結果 模擬空運前克氏原螯蝦肝胰腺小管結構正常,基膜完整,管腔清晰,呈星形,B細胞和R細胞分布較均勻,肝細胞內有較多空泡;肝胰腺小管周圍間質內主要為疏松結締組織及豐富的血竇,血竇內主要為顆粒細胞及半顆粒細胞,可見少量血細胞,未見明顯血細胞滲出及其他病理變化(圖5-A)。打孔組克氏原螯蝦肝胰腺小管結構較完整,部分肝細胞腫脹,致使管腔不規則,部分細胞輪廓模糊不清,變性細胞腫大變形,胞質內空泡體積增大,且空泡內出現顆粒樣內容物;肝胰腺小管周圍間質內可見少量均質紅染的蛋白樣物質沉積,未見明顯血細胞滲出及其他病理變化(圖5-B)。密封組克氏原螯蝦肝胰腺小管結構較完整,大量肝細胞明顯腫大變形,空泡體積顯著增大;肝胰腺小管周圍間質結構較正常,未見明顯血細胞滲出及其他病理變化(圖5-C)。
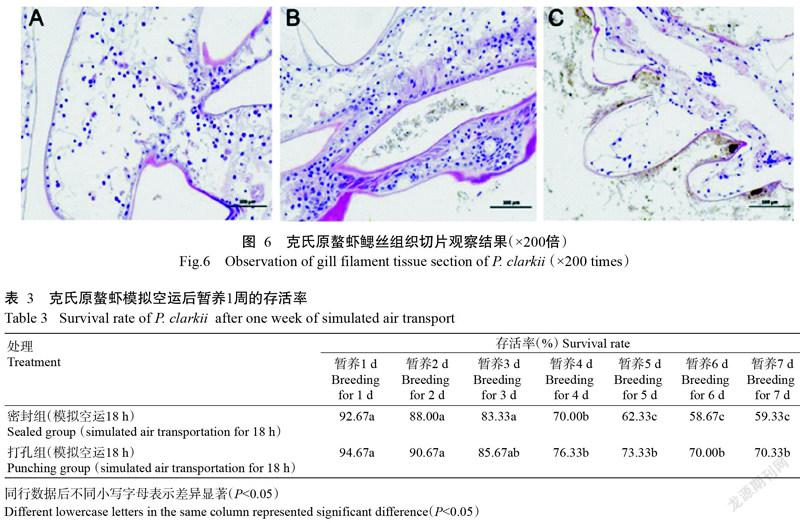
2. 5. 3 鰓絲組織切片觀察結果 模擬空運前克氏原螯蝦鰓膜結構完整,呼吸細胞排列整齊,能觀察到微血管,未見任何病理變化(圖6-A)。打孔組克氏原螯蝦鰓膜結構較完整,鰓絲出現部分附著物,微血管腔不明顯,呼吸上皮細胞排列較整齊,未出現明顯的變性壞死現象(圖6-B)。密封組克氏原螯蝦鰓絲有大量附著物,微血管腔結構已完全消失,大部分呼吸上皮細胞脫落、壞死,鰓膜結構遭到破壞(圖6-B)。
2. 6 模擬空運對克氏原螯蝦存活率的影響
經模擬空運18 h后,密封組克氏原螯蝦的存活率為82.00%,打孔組的存活率為89.00%,二者間的存活率差異不顯著。模擬空運后暫養1周發現,無論是密封組還是打孔組,克氏原螯蝦存活率均從第4 d開始顯著下降,至暫養第7 d密封組克氏原螯蝦的存活率顯著低于打孔組(表3)。
3 討論
水產品鮮活運輸包括有水保活運輸和無水保活運輸2種方式(Skudlarek et al.,2011;吳佳靜等,2016)。其中,有水保活運輸方式包括凈水法、充氧保活法、麻醉保活法和低溫保活法;無水保活運輸方式包括化學麻醉法及無水生態冰溫保活法。目前,關于水產動物長途運輸的研究以魚類較多,而蝦蟹類相對較少。宮春光等(2006)研究發現,無水充氧運輸過程中將日本對蝦密度控制在33.3 kg/m2,其13 h以內的運輸存活率均在90.0%以上。邊會喜(2010)研究發現,溫度在25.0 ℃以下時丁香油麻醉劑對克氏原螯蝦的保活效果較好,能有效提高長途運輸的存活率。謝慧明等(2010)通過對比幾種麻醉劑對克氏原螯蝦無水干運的影響,結果發現在25.0~30.0 ℃下存放時經60 mg/L丁香—乙醇水溶液麻醉的克氏原螯蝦保活效果最佳,與非麻醉組相比,20 h后的存活率提高9%。任紅梅等(2015)在對青蝦的模擬無水運輸研究中發現,6.0 ℃下運輸青蝦6 h以內其存活率顯著高于相同運輸時間其他溫度的存活率,均在80.0%以上。梁敏(2018)對凡納濱對蝦進行無水保活研究,結果發現保活時間在8 h內的最佳無水保活處理條件為溫度18.2 ℃和pH 6.08,該條件下凡納濱對蝦的存活率高達86.4%。徐子涵(2018)研究表明,將凡納濱對蝦事先在常溫充氧海水中暫養6 h,13.0 ℃水浴中冷擊3 min,充純氧包裝后15.0 ℃模擬空運12 h,結果發現水低溫脅迫會引起凡納濱對蝦的氧化應激反應,進而影響其抗氧化狀態,但并未引起難以逆轉的損傷。因此,在確保運輸存活率的同時應綜合考慮各項生理生化指標的變化(Pan et al.,2007;吳佳靜等,2016)。
本研究結果表明,經模擬空運后密封組克氏原螯蝦血清LZM活性、PPO活性及MDA含量顯著升高,而SOD活性顯著降低。MDA是反映細胞受損傷程度的重要指標,MDA積累越多,表明細胞受損傷程度越嚴重(姜冬梅和康波,2020);PPO是氧化酶源激活系統的重要組成部分,在機體應對抗氧化的過程中發揮重要作用;LZM具有抗菌消炎、抗病毒及增強機體免疫力等生理功能,還可激活血小板,改善組織局部血液循環障礙,增強局部防衛功能,具有止血消腫等作用;SOD是生物體內重要的抗氧化酶,能將有害的超氧自由基轉化為H2O2,再通過過氧化氫酶(CAT)將H2O2轉化為H2O(劉雨等,2019)。本研究的模擬空運也對克氏原螯蝦產生一定應激反應,克氏原螯蝦通過調節LZM活性和SOD活性以緩解應激帶來的影響,其體內MDA含量未出現明顯變化可能與這一調節有關。PPO在模擬空運中并未發揮明顯的作用,可能是因為應激時間不夠長,血清免疫指標尚未發生變化。此外,模擬空運結束后,密封組克氏原螯蝦肝胰腺蛋白質羰基含量、MDA含量和T-AOC水平均顯著高于打孔組及模擬空運前,而SOD活性變化不顯著。蛋白質羰基化是指氨基酸殘基側鏈中的氨基或亞氨基受到氧自由基攻擊,最后轉變成醛基并釋放NH3+的過程,羰基含量增加是蛋白喪失正常功能甚至被降解的重要原因(周家華等,2020)。說明模擬空運對克氏原螯蝦肝胰腺已造成損傷,與組織切片觀察到的肝胰腺細胞空泡化相互印證。T-AOC主要包括機體內的抗氧化酶體系和抗氧化物質體系,是衡量機體抗氧化系統功能狀況的綜合性指標(徐文杰等,2020)。模擬空運結束后,克氏原螯蝦肝胰腺T-AOC水平顯著提升,而SOD活性變化不顯著,說明克氏原螯蝦在模擬空運過程中受到脅迫后,其機體通過自身的免疫系統來抵抗應激反應,但SOD并不是主要的調節方式,而是通過別的途徑進行調節,因此后續研究宜選擇其他抗氧化指標進行檢測,如酸性磷酸酶或堿性磷酸酶等。Na+K+-ATP廣泛存在于生物體內,可催化腺嘌呤核苷三磷酸(ATP)水解生成二磷酸腺苷(ADP)和無機磷,在離子調節中發揮核心作用,對細胞滲透調節、物質吸收(葡萄糖和氨基酸等)及跨細胞離子運動等至關重要。鰓是克氏原螯蝦重要的呼吸器官,直接與外界環境接觸,因此其Na+K+-ATP活性能反映細胞主動運輸的能力,對判斷鰓絲的活力具有重要意義。密封組和打孔組克氏原螯蝦經模擬空運18 h后,其鰓絲Na+K+-ATP活性均顯著高于模擬空運前,說明克氏原螯蝦通過提升Na+K+-ATP活性以應對缺氧脅迫,從而增強鰓組織的工作效率,獲取更多氧氣。
經18 h模擬空運后,無論是打孔組還是密封組的克氏原螯蝦存活率均較高,與王海鋒等(2019)對克氏原螯蝦空氣暴露18 h的存活率相近。說明克氏原螯蝦具有較強的抗逆性,能通過自身免疫系統調節應對模擬空運應激,與Demers等(2006)對歐洲原產克氏原螯蝦進行低氧脅迫的研究結果相似。但模擬空運后暫養1周發現,密封組和打孔組的克氏原螯蝦存活率均從第4 d開始顯著下降,至暫養第7 d密封組克氏原螯蝦的存活率僅為59.33%,顯著低于打孔組克氏原螯蝦,未能滿足引種對存活率的要求,也說明模擬空運已對克氏原螯蝦造成嚴重損傷且不可逆轉。
4 結論
溫度和含氧量是影響克氏原螯蝦空運存活率的主要環境因子。模擬空運對克氏原螯蝦造成嚴重損傷且不可逆轉,在模擬空運結束后克氏原螯蝦因自身缺乏修復能力而大量死亡。因此,實際生產中可通過在運輸前投喂免疫增強劑,或在運輸后優化其暫養環境等方式以提高克氏原螯蝦空運存活率,進而確保其引種成功。
參考文獻:
邊會喜. 2010. 克氏原螯蝦遠程運輸保活技術研究[D]. 合肥:合肥工業大學. [Bian H X. 2010. Study on keep-alive technology of long-tange transportation of crayfish[D]. Hefei:Hefei University of Technology.] doi:10.7666/d.y1700626.
段亞飛,董宏標,王蕓,張真真,李卓佳,張家松. 2015. 干露脅迫對日本囊對蝦抗氧化酶活性的影響[J]. 南方水產科學,11(4):102-108. [Duan Y F,Dong H B,Wang Y,Zhang Z Z,Li Z J,Zhang J S. 2015. Effect of desiccation on antioxidant enzyme activities of kuruma shrimp(Marsupenaeus japonicus)[J]. South China Fisheries Science,11(4):102-108.] doi:10.3969/j.issn.2095-0780.2015. 04.015.
宮春光,李鳳臣,蘇利. 2006. 日本對蝦活蝦冬季運輸技術研究[J]. 水產科學,25(6):310-311. [Gong C G,Li F C,Su L. 2006. Transportation of live tiger shrimp(Marsupe-naeus japonicus) in winter[J]. Fisheries Science,25(6):310-311.] doi:10.3969/j.issn.1003-1111.2006.06.011.
姜愛蘭,王信海,金倩,王飛飛,王夏雯,張智慧,吳學軍,藺玉華. 2020. 不同底棲餌料對克氏原螯蝦生長、消化酶活性、血清生化指標及肌肉營養成分的影響[J]. 江西農業學報,32(1):97-101. [Jiang A L,Wang X H,Jin Q,Wang F F,Wang X W,Zhang Z H,Wu X J,Lin Y H. 2020. Effects of different benthos baits on growth,activities of digestive enzyme,serum biochemical indicators and muscle compositions of Procambarus clarkii[J]. Acta Agriculturae Jiangxi,32(1):97-101.] doi:10.19386/j.cnki.jxnyxb. 2020.01.16.
姜冬梅,康波. 2020. 亞精胺誘導小鼠卵巢組織氧化損傷的作用[J]. 甘肅農業大學學報,55(1):1-6. [Jiang D M,Kang B. 2020. Spermidine induced oxidative damage in ovary of female mouse[J]. Journal of Gansu Agricultural University,55(1):1-6.] doi:10.13432/j.cnki.jgsau.2020.01.001.
姜娜,王芳,路允良,董雙林. 2014. 干露對三疣梭子蟹抗氧化和應激能力的影響[J]. 水產學報,38(12):1996-2004. [Jiang N,Wang F,Lu Y L,Dong S L. 2014. Effects of air-exposure stress on antioxidant capacity and stress response of swimming crab (Portunus trituberculatus)[J]. Journal of Fisheries of China,38(12):1996-2004.] doi:10.3724/SP.J.1231.2014.49441.
連春盎,李健,李吉濤,馮艷艷. 2017. 干露脅迫對脊尾白蝦(Exopalaemon carinicauda)呼吸代謝相關酶的影響[J]. 漁業科學進展,38(4):53-60. [Lian C A,Li J,Li J T,Feng Y Y. 2017. Effects of dry exposure on the respira-tory metabolism-related enzymes of Exopalaemon carinicauda[J]. Progress in Fishery Sciences,38(4):53-60.] doi:10.11758/yykxjz.20160325001.
梁敏. 2018. 低溫與二氧化碳聯合脅迫凡納濱對蝦無水保活技術研究[D]. 湛江:廣東海洋大學. [Liang M. 2018. Study on the waterless transportation of Litopenaeus vannamei by combined stress of low temperature and carbon dioxide[D]. Zhanjiang:Guangdong Ocean University.]
劉雨,丁煒東,曹哲明,邴旭文,徐暢,楊帆,張晨光,谷心池,孫阿君. 2019. 急性氨氮脅迫對翹嘴鱖幼魚抗氧化酶活性及炎癥反應相關基因表達的影響[J]. 南方農業學報,50(8):1860-1868. [Liu Y,Ding W D,Cao Z M,Bing X W,Xu C,Yang F,Zhang C G,Gu X C,Sun A J. 2019. Effects of acute ammonia nitrogen stress on antioxidant enzymes activity and gene expression involved in inflammation of juvenile Siniperca chuatsi[J]. Journal of Sou-thern Agriculture,50(8):1860-1868.] doi:10.3969/j.issn. 2095-1191.2019.08.29.
任紅梅,萬玉芳,李雪梅. 2015. 青蝦無水低溫保活運輸初探[J]. 科學養魚,(5):50-51. [Ren H M,Wan Y F,Li X M. 2015. Preliminary study on anhydrous low temperature transportation of prawns[J]. Scientific Fish Farming,(5):50-51.]
王海鋒,成永旭,李京昊,奚業文,李嘉堯. 2019. 干露和再入水對克氏原螯蝦抗氧化應激能力的影響[J]. 南方水產科學,15(5):69-76. [Wang H F,Cheng Y X,Li J H,Xi Y W,Li J Y. 2019. Effects of desiccation and resubmersion on oxidative stress response of crayfish(Procambarus clarkii)[J]. South China Fisheries Science,15(5):69-76.] doi:10.12131/20190059.
溫久福,藍軍南,周慧,王鵬飛,區又君,李加兒. 2019. 不同鹽度對花鱸幼魚消化器官組織結構的影響[J]. 南方農業學報,50(12):2826-2832. [Wen J F,Lan J N,Zhou H,Wang P F,Ou Y J,Li J E. 2019. Effects of different salinities on histological structure of digestive organs of juvenile Lateolabrax maculatus[J]. Journal of Southern Agriculture,50(12):2826-2832.] doi:10.3969/j.issn.2095- 1191.2019.12.27.
吳佳靜,楊悅,許啟軍,黃寶生,聶小寶. 2016. 水產品保活運輸技術研究進展[J]. 農產品加工,(8):55-56. [Wu J J,Yang Y,Xu Q J,Huang B S,Nie X B. 2016. Research progress of aquatic products keep alive transportation technolog[J]. Farm Products Processing,(8):55-56.] doi: 10.16693/j.cnki.1671-9646(X).2016.08.045.
謝慧明,邊會喜,楊毅,張文成,林春銘,高香蘭. 2010. 克氏原螯蝦麻醉保活技術研究[J]. 食品科學,31(12):247-250. [Xie H M,Bian H X,Yang Y,Zhang W C,Lin C M,Gao X L. 2010. Clove oil anesthesia for the improvement of survival rate of crayfish[J]. Food Science,31(12):247-250.]
徐文杰,韓士群,周慶,巫佳杰,陳婷. 2020. 飼料中添加抗氧化劑對鯽魚抵抗藻毒素作用的影響[J]. 江蘇農業學報,36(2):417-422. [Xu W J,Han S Q,Zhou Q,Wu J J,Chen T. 2020. Effect of dietary antioxidant on microcystin-induced toxicosis in crucian[J]. Jiangsu Journal of Agricultural Sciences,36(2):417-422.] doi:10.3969/j.issn. 1000-4440.2020.02.023.
徐子涵. 2018. 南美白對蝦的無水低溫脅迫響應和無水保活運輸裝置[D]. 杭州:浙江大學. [Xu Z H. 2018. The response of Penaeus vannamei to waterless and low temperature stress and equipment for waterless transportation[D]. Hangzhou:Zhejiang University.]
周家華,秦洪強,葉明亮. 2020. 羰基化蛋白質組學分析進展[J]. 分析測試學報,39(1):82-88. [Zhou J H,Qin H Q,Ye M L. 2020. Progress in proteomic analysis of protein carbonylation[J]. Journal of Instrumental Analysis,39(1):82-88.] doi:10.3969/j.issn.1004-4957.2020.01.011.
de Oliveira U O,da Rosa Araújo A S,Belló-Klein A,da Silva R S M,Kucharski L C. 2005. Effects of environmental anoxia and different periods of reoxygenation on oxidative balance in gills of the estuarine crab Chasmagnathus granulata[J]. Comparative Biochemistry & Physiology. Part B:Biochemistry & Molecular Biology,140(1):51-57. doi:10.1016/j.cbpc.2004.09.026.
Demers A,Souty-Grosset C,Trouilhé M C,Füreder L,Renai B,Gherardi F. 2006. Tolerance of three European native species of crayfish to hypoxia[J]. Hydrobiologia,560:425-432. doi:10.1007/s10750-005-1466-9.
Lu Y L,Wang F,Li L,Dong S L. 2016. Responses of metabolism and haemolymph ions of swimming crab Portunus trituberculatus to thermal stresses:A comparative study between air and water[J]. Aquaculture Research,47(9):2989-3000. doi:10.1111/are.12752.
Oliveira G,Rossi I,da Silva R. 2001. Carbohydrate metabolism during anoxia and post-anoxia recovery in Chasmagnathus granulata crabs maintained on high-protein or carbohydrate-rich diets[J]. Marine Biology,139:335-342. doi:10.1007/s002270100569.
Pan L Q,Zhang L J,Liu H Y. 2007. Effects of salinity and pH on ion-transport enzyme activities,survival and growth of Litopenaeus vannamei postlarvae[J]. Aquaculture,273(4):711-720. doi:10.1016/j.aquaculture.2007.07.218.
Paterson B D,Grauf S G,Smith R A. 1997. Haemolymph chemistry of tropical rock lobsters (Panulirus ornatus) brought onto a mother ship from a catching dinghy in Torres Strait[J]. Marine and Freshwater Research,48(8):835-838. doi:10.1071/MF97069.
Rabin O,Piciotti M,Drieu K,Bourre J M,Roux F. 1996. Effect of anoxia and reoxygenation on antioxidant enzyme activities in immortalized brain endothelial cells[J]. In Vitro Cellular & Developmental Biology,32(4):221-224. doi:10.1007/BF02722949.
Skudlarek J G,Coyle S D,Bright L A,Tidwell J H. 2011. Effect of holding and packing conditions on hemolymph parameters of freshwater prawns,Macrobrachium rosenbergii,during simulated waterless transport[J]. Journal of the World Aquaculture Society,42(5):603-617. doi:10. 1111/j.1749-7345.2011.00508.x.
Zenteno-Savín T,Saldierna R,Ahuejote-Sandoval M. 2006. Superoxide radical production in response to environmental hypoxia in cultured shrimp[J]. Comparative Bioche-mistry and Physiology. Part C:Toxicology & Pharmaco-logy,142(3-4):301-308. doi:10.1016/j.cbpc.2005.11.001.
(責任編輯 蘭宗寶)
