牛乳酪蛋白基因多態性研究進展
2021-02-25 09:29:38趙烜影劉振民雍靖怡穆海菠
乳業科學與技術 2021年1期
關鍵詞:檢測
趙烜影,劉振民,雍靖怡,穆海菠,李 楠*
(乳業生物技術國家重點實驗室,上海乳業生物工程技術研究中心,光明乳業股份有限公司乳業研究院,上海 200436)
牛乳是犢牛最重要的食物,也是人類補充蛋白質和微量元素的常見來源。牛乳中有2大類蛋白質:乳清蛋白和酪蛋白(casein,CN),乳清蛋白由β-乳球蛋白(β-lactoglobulin,β-Lg)、α-乳白蛋白(α-lactalbumin,α-La)、免疫球蛋白、糖肽、牛血清白蛋白和次要蛋白,如乳過氧化物酶、溶菌酶和乳鐵蛋白組成[1];CN約占牛乳蛋白的76%~86%,是由乳腺上皮細胞合成的一種含有大量鈣和磷的蛋白,以膠束形式存在于牛乳中。酪蛋白含有人體所必需的8 種氨基酸。牛乳含有4 種酪蛋白:αs1-CN(CSN1S1,占總酪蛋白的39%~46%)、αs2-CN(CSN1S2,占總酪蛋白的8%~11%)、β-CN(CSN2,占總酪蛋白的25%~35%)和κ-CN(CSN3,占總酪蛋白的8%~15%)[2]。這4 種酪蛋白通過磷酸化、糖基化和水解會產生一部分新的酪蛋白,如αs-CN通過磷酸化、κ-CN通過糖基化以及β-CN經過水解,會生成γ-酪蛋白和多肽等[3]。酪蛋白多態性被認為會影響乳產量、乳的理化特性和營養成分、乳制品加工特性以及人類的營養而得到學者們的重視。
1 酪蛋白基因多態性
超過95%反芻動物乳中的酪蛋白都是通過6 個結構基因編碼形成的,在6號染色體上4 個酪蛋白基因組成了一個250 kb的片段,如圖1所示。而水牛乳的4 種酪蛋白則同樣由4 個在水牛7號染色體上呈簇狀分布的酪蛋白基因編碼[5]。Aschaffernburg等[6]以β-Lg為對象,找出了2 種基因型,發現了乳蛋白的基因多態性。越來越多的學者開始研究乳蛋白基因多態性,乳蛋白基因多態性在不同牛種之間存在明顯差異,為了適應當地飼育條件而雜交培育出的本地特色奶牛則進一步豐富了牛乳酪蛋白的基因多態性。
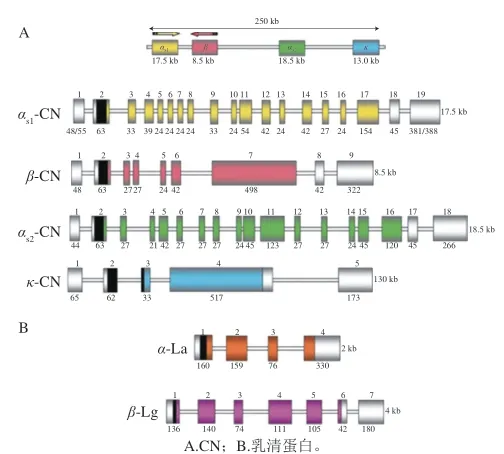
圖 1 主要乳蛋白編碼轉錄單位的結構組織[4]Fig. 1 Structural organization of major milk protein coding transcription units[4]
在眾多研究中發現會出現同一種等位基因在不同物種中命名不同的情況,這可能與檢測技術不同相關。盡管在DNA水平上的鑒定可以發現很多單核苷酸多態性(single nucleotide polymorphism,SNP)位點,但是這些突變位點可能是沒有更改氨基酸序列的同義突變位點,進而在蛋白質水平上無法檢測出新的蛋白質亞型。Farrell等[1]提出的對普通牛、瘤牛、牦牛、爪哇野牛乳蛋白基因型命名法更加適用于常見的乳蛋白等位基因。目前已被報道的牛乳蛋白基因有A、B、C、D、E、F、G、H、I、J 10 種CSN1S1等位基因;A、B、C、D、E 5 種CSN1S2等位基因;A1、A2、A3、B、C、D、E、F、G、H1、H2、I、J、K、L 15 種CSN2等位基因和A、B、B2、C、D、E、F1、F2、G1、G2、H、I、J 13 種CSN3等位基因[7-9]。
1.1 α-CN基因多態性
αS1-CN由214 個氨基酸組成的肽鏈和1 個含有15 個氨基酸的前導鏈組成[10]。編碼αS1-CN的CSN1S1基因全長17 508 bp,有18 個內含子及19 個外顯子[11]。αS1-CN在磷酸鈣的運輸、酪蛋白膠束的形成以及酪蛋白從內質網傳遞到高爾基體中起到關鍵作用。CSN1S1基因多態性由于與牛乳的營養價值和加工特性相關而得到廣泛關注[12]。CSN1S1的B等位基因最為常見;在DNA序列上的26 139 bp處由C突變為T的A等位基因在埃及、意大利等地水牛中被多次報導;C等位基因在利木贊牛、瑞士褐牛、弗萊維赫牛等雜交品種中發生頻率最高(>10%)[2]。Fan Xinyang等[11]在沼澤水牛中發現了由c.609T>G產生的D等位基因。I等位基因被證實是由c.296A>T產生[1]。2013年,在吉爾牛和短角牛中發現了c.543G>T,并將其命名為J等位基因[9],至此CSN1S1基因增加為10 種。除B、C等位基因外,其余等位基因僅在少數品種中出現。
編碼αS2-CN的CSN1S2基因全長18 480 bp,有18 個外顯子和17 個內含子[13]。CSN1S2基因多態性被認為與泌乳性能有關。目前已報道的CSN1S2等位基因共有A、B、C、D、E 5 種,大都集中在第3、第6及第8個外顯子中。等位基因B、C、D、E均被認為是由A等位基因突變而來,A等位基因外顯子3的第17位堿基發生了C→T的替換,產生了B等位基因;外顯子6的第2位堿基發生了A→G的替換,產生了C等位基因;外顯子8的第26位堿基發生了G→T的替換,產生了D等位基因;外顯子3的第13位堿基發生了C→T的替換,產生了E等位基因[13]。
1.2 β-CN基因多態性
β-CN是一種疏水性球狀蛋白,由209 個氨基酸殘基組成,分子質量為25.382 kDa,牛乳中的β-CN以完全磷酸化形式存在[14]。編碼β-CN的CSN2基因全長8.5 kb,由9 個外顯子和8 個內含子組成[15]。β-CN的基因多態性被認為與Ⅰ型糖尿病、心血管疾病、自閉癥、精神分裂和免疫缺陷等多種疾病相關[16]。目前已被報道的CSN2等位基因共有15 種(A1、A2、A3、B、C、D、E、F、G、H1、H2、I、J、K、L)[17]。CSN2等位基因及其發生頻率因檢測品種、樣本量及地域的差異而存在較大差異。意大利北部荷斯坦牛的檢測結果顯示,A1發生頻率為0.371,A2發生頻率為0.546,而美國對于荷斯坦牛的檢測結果與上述結果相反,A1的發生頻率(0.31~0.66)超過了A2(0.24~0.62)[16-17]。但總的來說,在現有研究結果中,A1和A2等位基因以超過0.4的發生頻率最為常見[18]。B等位基因的發生頻率為0.01~0.10[17]。I等位基因發生頻率在大部分品種中均低于0.03,但在意大利紅白花牛(0.14)和德國荷斯坦-弗里森雜交牛(0.192)中顯示出較高的頻率[19]。有學者在韓國本地牛類中檢測到A4等位基因,但尚未發現其SNP位點,D等位基因只在瘤牛中被發現,E等位基因只在皮埃蒙特品種中被發現[4]。A3、C、F、G等位基因較為罕見,發生頻率均低于0.01[20]。目前尚未有關于H1和H2等位基因發生頻率數據。
1.3 κ-CN基因多態性
κ-CN多存在于酪蛋白膠束的表面,對維持膠束的穩定性起到重要作用[21]。κ-CN含有半胱氨酸和蛋氨酸2 種含硫氨基酸,是一種糖基化酪蛋白[1],由編碼190 個氨基酸的肽鏈組成,其中21 個氨基酸形成信號肽前導鏈,編碼κ-CN的CSN3基因全長573 pb,含有5 個外顯子和4 個內含子,大多數編碼序列包含在第4個外顯子中[22]。CSN3基因首次被證實是在1964年,迄今為止已發現13 個等位基因(A、B、B2、C、D、E、F1、F2、G1、G2、H、I和J)及1 個同義變體(A1)[23]。目前已知的等位基因都應該是A等位基因由于氨基酸的缺失或置換等突變引起的,其中A、B等位基因因與牛乳產量及凝乳特性有關而被研究人員重視[11]。在荷斯坦牛中A等位基因的發生頻率為0.736,遠高于B等位基因的0.186,但是在娟姍牛中B等位基因的發生頻率(0.692)超過了A等位基因(0.308)[9]。A等位基因具有136Thr(ACC)/148Asp(GAT),而B等位基因具有136Ile(ATC)/148Ala(GCT),E等位基因將氨基酸155Ser(AGC)更改為155Gly(GGC)[4]。G等位基因比A、B等位基因少見,僅在德國黑白花牛、荷斯坦-弗里斯蘭牛、捷克弗萊維赫牛、艾爾郡牛和瑞典紅牛等少數品種中發現過[24]。
2 基因多態性檢測方法
乳蛋白的基因多態性主要是由氨基酸的缺失或置換、糖基化、磷酸化位點的差異引起的,目前,關于基因多態性的檢測大都是在蛋白水平和DNA水平上進行。蛋白水平檢測常見的方法包括毛細管區帶電泳、反相高效液相色譜(reversed-phase high-performance liquid chromatography,RP-HPLC)、等電聚焦電泳或尿素-聚丙烯酰胺凝膠電泳[25-28]。最近,液相色譜-串聯質譜或液相色譜-高分辨率質譜等更強大的技術已被成功地用于蛋白質變體的鑒定和定量[29-30],主要包括等位基因特異性聚合酶鏈式反應(allele specific polymerase chain reaction,AS-PCR)、限制性片段長度多態性PCR(restriction fragment length polymorphism PCR,RFLP-PCR)、單鏈構象多態性PCR(single strand conformation polymorphism PCR,SSCP-PCR)、創造酶切位點PCR(amplification created restriction site PCR,ACRS-PCR)等方法,近年來還出現了TaqMan法及焦磷酸測序等技術[31-32]。
2.1 蛋白質水平檢測方法
HPLC法雖然不是最常見的蛋白質組學分析技術,但適用于初始階段分離多種蛋白質混合物。RP-HPLC法通常采用十八烷基-硅膠固定相和極性強于固定相的低離子強度酸性有機流動相。隨著流動相有機溶劑濃度的改變,蛋白質親硅醇基效應和疏水性會影響到蛋白質的保留時間。不同亞型的酪蛋白由于-NH2和-COOH等極性和氫鍵作用較強的基團含量及位置的不同,保留時間不同,而被分離。C8色譜柱屬于短鏈且不具有支鏈的鍵合固定相,其保留性弱于C18,因而更適用于出峰時間差異較小的酪蛋白亞型之間的測定。劉亞楠等[33]通過RP-HPLC法成功檢測到中國水牛4 種酪蛋白的不同亞型,發現κ-CN出現了3 種不同的峰,即A、B、C 3 種等位基因,αS1-CN、αS2-CN及β-CN各出現了2 個不同的峰。陳晨等[34]利用RP-HPLC法在雜交水牛的κ-CN中發現了新的D等位基因。操作簡便、耗時短、回收率高、可以檢測出沉默變異體等優點使HPLC法成為檢測酪蛋白多態性的有效方法。
液相色譜-質譜聯用方法是將液相色譜分離后的目標蛋白質通過電離的方式引入質譜,得到蛋白質不同亞型特異性肽段的質譜圖。將測定出的單同位素質量與已知的蛋白質亞型單同位素質量進行對比,來鑒定蛋白質的多態性。Nguyen等[31]通過使用與Q-Exactive質譜儀偶聯的Ultimate 3000 UPLC成功檢測到越南黑白花荷斯坦F2(HF2)乳樣中β-CN的3 個亞型,在85 頭HF2乳樣中檢測到3 個單同位素質量分別為24 008.143 0、23 968.167 0、23 950.226 0 Da的亞型,分別與β-CN亞型中A1、A2和I的理論單同位素質量24 008.161 6、23 968.151 6、23 950.191 6 Da匹配。
不同的電泳技術則是通過蛋白質亞型在電泳或等電聚焦過程中遷移率的差異進行檢測,可以對多種乳蛋白進行同時檢測。Caroli等[27]開發了一種將等電聚焦電泳后膠片浸泡在三氯乙酸中,將β-CN亞型固定化的定量方法,避免了考馬斯亮藍染色過程β-CN與β-Lg重疊,該方法不僅成功檢測出β-CN的A1和A22 種亞型,還能夠對樣品中的β-CN亞型進行定量分析。馮慿等[35]建立的毛細管區帶電泳方法可以對液態乳和乳粉中β-CN亞型進行定性及定量分析,成功檢測出A1及A22 種亞型。Duarte-Vázquez等[28]通過尿素-聚丙烯酰胺電泳成功檢測出荷斯坦及娟姍牛β-CN的A1、A22 種亞型。
蛋白質水平上的檢測方法由于能夠更加直觀驗證酪蛋白的基因型而在實際中應用更加廣泛。
2.2 DNA水平檢測方法
焦磷酸測序作為一種新型的酶聯級聯測序技術適用于對已知短序列的測序分析,其通過引物與模板DNA退火后,在DNA聚合酶、三磷酸腺苷(adenosine triphosphate,ATP)硫酸化酶、熒光素酶和ATP雙磷酸酶4 種酶的協同作用下,將引物上dNTP的聚合與熒光信號的釋放偶聯,檢測熒光的釋放和強度,達到實時測定DNA序列的目的[36]。可以大通量、低成本、實時、快速、直觀地進行SNP測定。任大喜等[37]采用焦磷酸測序法分析中國荷斯坦牛、娟姍牛和水牛的乳蛋白基因多態性,并找出了水牛κ-CN的特異性多態位點。
AS-PCR方法的原理是將突變堿基設計在引物3’末端,在PCR中若突變堿基與模板互補,獲得特異擴增條帶,則表明被測基因含有該種突變[38]。Gholami等[39]使用AS-PCR技術擴增了1 個854 bp的片段,成功鑒定出伊朗荷斯坦牛、娟姍牛、當地雜交奶牛β-CN的A1、A2等位基因,并測定出A1等位基因的發生頻率為荷斯坦牛0.5、娟姍牛0.51、西斯塔尼牛0.54、塔萊西牛0.49、馬贊達拉尼牛0.46。Firouzamandi等[40]同樣利用AS-PCR技術在荷斯坦Sarabi牛中檢測到β-CN的A1、A2等位基因。
RFLP-PCR技術是將PCR擴增后的DNA用特異性內切酶切分成長短不一的片段,并在凝膠電泳上分辨。因為不同等位基因的限制性酶切位點分布不同,會產生不同長度的DNA片段條帶。Cinar等[41]利用該方法設計正向引物5’-ATCATTTATGGCCATTCCACCAAAG-3’、反向引物5’-GGCCATTTCGCCTTCTCTGTAACAGA-3’,成功測定了安那托利亞水牛κ-CN的B等位基因。Jawane等[42]使用CASB67R和CASB122L引物擴增印度瘤牛β-CN基因外顯子VII的一部分,并用TaqI限制酶對251 bp的PCR擴增序列進行限制性酶切,得到3 種基因型:A1A1(213、38 bp)、A1A2(251、213、38 bp)和A2A2(251 bp)。
SSCP-PCR技術的基本原理是將PCR擴增后的DNA片段經變性形成單鏈DNA,單個堿基的差異會在中性聚丙烯酰胺凝膠電泳時形成不同的立體構象,改變泳動速率,產生不同的條帶。Chiatti等[43]運用此方法成功鑒定出山羊乳αS2-CN的A、B、C、E共4 個等位基因,并通過等電聚焦在蛋白質水平上驗證,他們認為SSCP-PCR技術相比RFLP-PCR技術在DNA水平上檢測CSN1S2等位基因會更加快捷、平價。Hamza等[44]利用SSCP-PCR技術設計正向引物5’-CTAAATCTGGCATAAAAGTA-3’、反向引物5’-AATCACGGACTAAATAA-3’的引物對,成功檢測出中國荷斯坦奶牛κ-CN的C、T等位基因。
TaqMan熒光探針是一種寡核苷酸探針,其5’末端攜帶熒光基團,如FAM、TET、VIC、HEX等,3’端攜帶猝滅基團,如TAMRA、BHQ等。其原理為:用1 對雙標記的TaqMan熒光探針分別針對雙等位SNP的不同等位基因,只有完全匹配的探針可以擴增出其對應的等位基因。若只有VIC熒光信號明顯增強表明為等位基因1,若只有FAM熒光信號明顯增強表明為等位基因2,當VIC與FAM熒光信號均明顯增強則為等位基因1與2的雜合型。而新型TaqMan-MGB(minor groove binder)探針使該技術既可進行基因定量分析,又可分析基因突變(SNP)[45]。Kusza等[46]利用該技術發現羅馬尼亞本土扎克爾綿羊CSN2基因的A等位基因發生頻率為0.009 8。
3 酪蛋白基因多態性的應用
3.1 酪蛋白基因多態性在生產加工中的應用
作為奶牛的重要經濟指標,泌乳性能由多個基因控制,乳蛋白基因則是影響泌乳性能的主要基因,乳蛋白中編碼酪蛋白和β-Lg的基因對奶牛的泌乳性能影響最大。梁艷等[47]對智利進口的荷斯坦牛酪蛋白多態性進行研究發現,β-CN多態性對日產奶量有極顯著影響,AB亞型個體的日產奶量極顯著高于AA亞型(P<0.01)。楊雪瑤等[48]研究寧夏地區荷斯坦奶牛發現,含κ-CN的AA基因型奶牛的產奶量顯著高于CA基因型(P<0.05),極顯著高于CC基因型(P<0.01),說明κ-CN的A等位基因有利于提高日產奶量。Morkuniene等[49]對立陶宛牛群κ-CN多態性進行研究同樣發現,A等位基因有利于提高日產奶量。
除了影響奶牛泌乳性能以外,酪蛋白多態性對牛乳營養成分的影響更為顯著。Gurses等[23]研究κ-CN發現,含AB亞型的荷斯坦牛乳蛋白質及乳糖含量顯著高于AA及AE亞型;含A等位基因的娟姍牛乳脂含量顯著低于含B等位基因。也有研究顯示,水牛β-CN的B等位基因與乳蛋白含量成正比,而A等位基因則與乳脂含量成正比[50]。黃改奶牛αs2-CN的A等位基因與乳脂及乳蛋白含量成正相關[13]。Li Shanshan等[51]研究中國水牛的酪蛋白遺傳多態性與乳蛋白組成之間的關聯,發現κ-αs1-β復合基因型中,BB-AB-AA型和AB-AB-AA型分別對κ-CN和αs2-CN含量有顯著影響。Day等[52]研究酪蛋白多態性對乳蛋白膠束大小的影響,發現κ-CN的AA亞型酪蛋白膠束顯著大于AB及BB亞型,并且小膠束酪蛋白((151.0±2.2) nm)的糖基化比例更高,即含B等位基因的酪蛋白更容易糖基化;同時還發現小膠束酪蛋白中β-CN出現基因突變的頻率更高。陳晨等[34]研究中國水牛的酪蛋白多態性發現,水牛αs1-β-κ-CN復合基因為ABBB-AB型,κ-CN的C等位基因均與乳蛋白粒度存在顯著相關性,αs1-CN和κ-CN基因型為A等位基因的蛋白粒度大于基因型為其他等位基因的蛋白。
酪蛋白多態性對牛乳加工性能的影響也是學者們的研究重點。黃麗[53]、李玲[54]等研究水牛αs1-CN對馬蘇里拉干酪及類蒙特利杰克半硬質干酪加工及品質的影響,發現水牛αs1-CN多態性對馬蘇里拉干酪組成影響顯著,αs1-CN為AB亞型的水牛乳制作的馬蘇里拉干酪在脂肪、蛋白質含量以及干酪硬度、彈性、膠黏性、咀嚼性等指標方面都優于BB亞型;但是αs1-CN為BB亞型的水牛乳制作的類蒙特利杰克半硬質干酪在各項指標上均顯著優于AB亞型。Zicarelli等[55]研究發現,含有意大利地中海水牛αs1-CN的B等位基因以及κ-CN的F1等位基因的牛乳凝乳效率顯著高于含其他等位基因的牛乳。Nguyen等[56]研究奧克蘭當地牧場奶牛β-CN的A1與A2等位基因對牛乳中鈣分布、酸凝固、起泡特性及酸乳凝膠微觀結構的影響,發現含有β-CN A2等位基因的牛乳中游離Ca2+含量顯著高于含A1等位基因的牛乳,泡沫形成能力增強,需要更長的時間才能凝固;A2A2亞型酸乳的凝膠強度較弱,微觀結構較多孔,表明該凝膠更細膩,在外部機械力的作用下容易破裂和變形,因此,在處理和運輸A2A2亞型酸乳產品期間可能需要格外小心,然而從健康和營養的角度來看,A2A2亞型酸乳較軟,而較弱和較多孔的凝膠有助于蛋白質更快地分解,在胃中酸性條件下更好被消化。在對中國水牛β-CN基因多態性的研究過程中發現,體外消化實驗表明,AB亞型消化最快,其次為AA亞型,BB亞型最難消化[50]。
3.2 酪蛋白基因多態性對人體健康的潛在影響
關于酪蛋白多態性對人體健康的影響,學者們大都聚焦在β-CN的A1與A2等位基因,這可能與β-CN含量及A1和A2等位基因發生頻率高有關。β-CN的A1等位基因在人體內經過胃蛋白酶的消化會產生一種阿片肽——β-酪啡肽7(β-casomorphin 7,BCM7),其產生機制見圖2。BCM7被認為與Ⅰ型糖尿病、缺血型心臟病、精神分裂、自閉癥、嬰兒猝死綜合征、動脈粥樣硬化、冠狀動脈性心臟病、孤獨癥譜系障礙等疾病有關[16,32,57]。
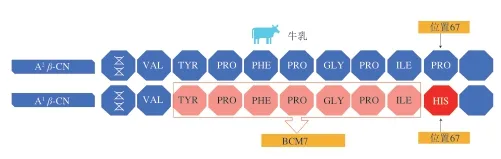
圖 2 BCM7產生機制[57]Fig. 2 BCM7 generation mechanism[57]
多位學者研究發現A1等位基因的攝入量與Ⅰ型糖尿病的發病率有顯著相關性;對BioBreeding(BB)大鼠的動物實驗也顯示,A1等位基因有誘發Ⅰ型糖尿病的作用[58]。關于誘發機制目前存在2 種假說:1)含A1等位基因的β-CN會刺激T免疫細胞產生免疫反應及抗體免疫反應,這會促進Ⅰ型糖尿病的發病;2)類似于BCM-7的阿片類藥物可能會干擾代謝過程,包括血糖水平的調節和胰島素的產生,進而導致Ⅰ型糖尿病,動物實驗也證實了阿片肽類藥物的共同給藥受體拮抗劑納洛酮有效減弱了A1等位基因的作用[58]。
對食物攝入的不良反應具有非常多樣的病因和癥狀,在牛乳中主要表現為乳蛋白過敏和乳糖不耐癥。乳蛋白過敏是最常見的食物過敏癥狀,對兒童的影響較大,尤其是新生兒。一般認為BCM7會刺激T免疫細胞,進而引起乳蛋白過敏,β-CN的A2等位基因則會降低致敏乳蛋白的表達[59]。當然,除了β-CN以外,β-Lg與α-La也是乳蛋白過敏的重要過敏原。乳糖不耐癥則是因為體內缺乏乳糖酶,乳糖在腸道中分解為乳酸,破壞了腸道酸堿平衡,進而引起腹瀉,是一種多發于亞洲地區的先天性遺傳疾病,部分人體內的乳糖酶活力會隨著年齡的增加而降低。He Mei等[60]通過臨床實驗研究發現,含A1β-CN牛乳會降低乳糖酶活性,并增加胃腸道癥狀,而含A2β-CN牛乳可減輕急性乳糜瀉引發的胃腸道不適癥狀。目前尚未有直接的證據解釋β-CN的A1等位基因對腸胃影響的機理。
氧化代謝是維持細胞生命活動的重要功能之一,新陳代謝失調會產生許多活性氧和自由基,引起氧化應激。過氧化物自由基、羥自由基、超氧自由基和過氧化氫等自由基不僅會引起蛋白變質,還會損害生物系統。由于蛋白質和多肽具有特定的或非特異性的生物學機制,如使活性氧失活、清除促氧化的金屬離子、降低氫過氧化物水平以及改變系統的內部特性,因此蛋白質和多肽具有抗氧化潛力;對中國水牛酪蛋白多態性與其抗氧化性的研究發現,AA亞型β-CN的1,1-二苯基-2-三硝基苯肼自由基清除能力優于BB亞型,BB亞型的羥自由基清除能力和還原能力優于AA亞型[50]。
4 結 語
酪蛋白作為牛乳中的重要組成部分,具有極高的營養價值。4 種酪蛋白均存在基因多態性,這些多態性對牛乳的組成成分、牛乳品質、牛乳的生產加工特性以及乳制品的營養、口感、品質具有重要影響。加強對酪蛋白多態性的基礎研究可以為保護牛的基因多態性、奶牛的定向育種以及精準設計功能性乳制品提供理論基礎,引導中國乳業向高附加值、高收益多樣化發展。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48