基于轉錄組測序的高原鼢鼠多態性微衛星標記篩選
2021-03-02 02:42:50劉倩倩譚宇塵姚寶輝康宇坤蘇軍虎
草業科學 2021年12期
劉倩倩,譚宇塵,姚寶輝,康宇坤,蘇軍虎,3
(1.甘肅農業大學草業學院 / 草業生態系統教育部重點實驗室 / 中—美草地畜牧業可持續發展研究中心, 甘肅 蘭州 730070;2.甘肅農業大學—新西蘭梅西大學草地生物多樣性研究中心, 甘肅 蘭州 730070;3.甘肅省祁連山草原生態系統野外科學觀測研究站, 甘肅 蘭州 730070)
微衛星DNA 也稱為簡單重復序列(simple sequence repeat, SSR),通常是由1~6 個核苷酸重復單元構成的串聯重復序列,在真核生物基因組中,微衛星DNA 絕大多數位于基因非編碼區,但也存在于編碼序列及外顯子中[1-2]。微衛星具有分布廣泛、高度多態性及共顯性遺傳等優點,已被廣泛應用于確定種群間的遺傳分化、估計種群結構和種群的遺傳多樣性研究中[3]。微衛星DNA 按照其來源可分為基因組SSR (genomic simple sequence repeat,gSSR)和表達序列標簽SSR (expressed sequence tagssimple sequence repeat, EST-SSR)。與gSSR 相 比,EST-SSR 來源于DNA 的轉錄區域,其開發成本低、速度快,具有良好的物種間通用性且數量大等特點[4],已成為對缺少基因組數據信息的生物進行分子標記開發的有效方法之一。
高原鼢鼠(Eospalax baileyi)隸屬于嚙齒目鼴形鼠科鼢鼠亞科凸顱鼢鼠屬,是青藏高原高寒草甸生態系統的特有種,它們的大部分活動均在地下洞穴系統中進行,主要依靠植物的地下貯存器官作為食物[5]。近年來,在自然因素和人為活動的影響下,高原鼢鼠的高密度種群加速了草地退化,危及青藏高原生態安全和區域經濟的發展[6]。隨著國家生態安全戰略的實施,高原鼢鼠在高寒草甸生態系統中的有效管理備受重視,然而有效的防控工作不僅需要了解高原鼢鼠的生物學特性,還需要了解其種群生態作用和種群生態學等方面的知識[7]。已有研究表明高原鼢鼠在不同地理種群間存在著限制性基因流,會導致種群分化與種群遺傳結構改變[8-9],影響到群體遺傳多樣性和繁殖策略,進而影響高原鼢鼠的種群動態及其防控決策[10]。但對于高原鼢鼠種群自身的近交回避、親權識別和繁殖策略等問題至今仍然知之甚少,解決這些問題對于高原鼢鼠的有效管理至關重要,因此迫切需要有效的分子生物學技術和方法得以應用。
近年來,學者們利用線粒體基因細胞色素b[11]和D-loop 區[12]等測序方法研究了不同群體高原鼢鼠的遺傳多樣性和遺傳結構。關于高原鼢鼠核基因組微衛星位點開發也有零星的報道,Su 等[13]基于454 高通量測序,獲得了12 個微衛星位點。康宇坤等[14]分析并驗證了跨種的通用性,但相較于模式生物和其他生物具有上千個微衛星位點,高原鼢鼠現有的40 多個微衛星位點仍無法滿足不同地理群體的分析需求,還需更多的微衛星位點進行婚配方式、遺傳結構和基因流等檢測,而迄今對高原鼢鼠的EST-SSR 標記篩選未見報道。鑒于此,本研究基于Illumina HiSeq 測序平臺,測定高原鼢鼠的特異性微衛星標記,分析其分布特征,并對這些SSR 位點在高原鼢鼠中的多態性進行驗證,以期篩選出具有高多態性的高原鼢鼠EST-SSR 引物,旨在為其種群遺傳學和分子生態學等相關研究提供技術基礎。
1 材料與方法
1.1 材料
選用高原鼢鼠63 只,其中雌性46 只,雄性17只,樣本采集于甘肅省甘南州碌曲縣境內(100°46′—104°44′ E,33°06′—36°10′ N),用活捕籠非損傷性捕捉,麻醉處死,迅速剖取其大腦組織,存入液氮速凍后置于-80 ℃冰箱中保存備用,采集腿部肌肉保存于95%酒精中,用于基因組DNA 的提取。
1.2 試驗方法
1.2.1 RNA 提取及EST-SSR 序列的獲得
取24 只高原鼢鼠,雌雄各12 只,每只取0.3 g 新鮮的大腦組織,用MiniBEST Universal RNA Extraction Kit (Takara,日本)提取總RNA,通過1.0%瓊脂糖凝膠電泳和Nanodrop-2000 紫外分光光度儀檢測總RNA的濃度和純度,送樣至北京諾禾致源科技股份有限公司構建cDNA 文庫,利用Illumina HiSeq 平臺進行測序,對其得到的數據進行評估、過濾(去除接頭、低質量區域等),用Trinity-v2.5.1 軟件進行拼接組裝[15],共得到46 858 條unigenes,利用SSR 分析軟件MISA 1.0(http://pgrc.ipkgatersleben.de/misa/)搜索高原鼢鼠組織轉錄組數據中潛在的SSR 位點[16]。
1.2.2 DNA 的提取及檢測
選擇63 只高原鼢鼠腿部肌肉組織(保存于95%酒精中)為試驗材料,運用常規的酚氯仿抽提法提取基因組DNA。通過紫外分光光度計檢測DNA 的濃度和純度,用1%瓊脂糖凝膠電泳檢測DNA 的質量,-20 ℃保存。
1.2.3 引物的合成及PCR 擴增
基于轉錄組測序得到的29 090 條序列,用Primer 3.0 軟件設計引物,共隨機選取104 對引物由上海生工生物工程股份有限公司合成。PCR 反應體系總體積25 μL,其組成為2 × Taq PCR Master Mix 12.5 μL,上下游引物各1 μL,模板DNA 1 μL,ddH2O 補足至25 μL。反應條件為95 ℃預變性4 min,95 ℃變性45 s,53 ℃退火45 s,72 ℃延伸30 s,共30 個循環,72 ℃延伸10 min,4 ℃保存。
1.2.4 微衛星基因型分型
擴增產物經1%瓊脂糖凝膠電泳擴增出清晰條帶后,利用8%聚丙烯酰胺凝膠電泳進行初步多態性篩選,篩選出12 對引物進行毛細管電泳檢測,在正向引物5’末端用FAM、HEX 及TAMR 3 種熒光染料標記,送至上海生工生物工程股份有限公司,用ABI 3 730 分析儀進行毛細管電泳分型。
1.2.5 微衛星數據分析
用GeneMapper 4.0 對SSR 分型后得到的熒光電泳圖譜進行判讀,并進一步人工校正,獲取每個微衛星位點在不同個體上的等位基因片段大小;用POPGENE 1.32 統計等位基因數(number of alleles,Na),計算觀測雜合度(observed heterozygosity, Ho)和期望雜合度(expected heterozygosity, He);用Cervus 3.0計算多態信息含量(polymorphism information content,PIC)并檢驗哈迪—溫伯格平衡(Hardy—Weinberg equilibrium, HWE)。
2 結果與分析
2.1 轉錄組SSR 重復類型分析
對高原鼢鼠的轉錄組微衛星數據進行篩選,完美型微衛星是最常見的類型,共有29 090 個,其次是復合型微衛星,不完美型微衛星數量最少,只有1 001 個。本研究僅對高原鼢鼠轉錄組中1~6 堿基重復完美型SSR 進行分析,結果顯示,對高原鼢鼠轉錄組中46 858 條unigenes 的數據進行搜索,共找到29 090 個微衛星位點,分布在7 306 條unigenes中,發生頻率(含有SSR 的unigenes 數量與總unigenes數量之比)為25.12%。在高原鼢鼠SSR 重復基序中,以單核苷酸重復單元的SSR 含量最多,所占比例為56.57%,其次是二核苷酸及三核苷酸重復,所占比例分別為28.65%和10.17%,四核苷酸、五核苷酸、六核苷酸重復的SSR 類型較少,共占4.61% (表1)。
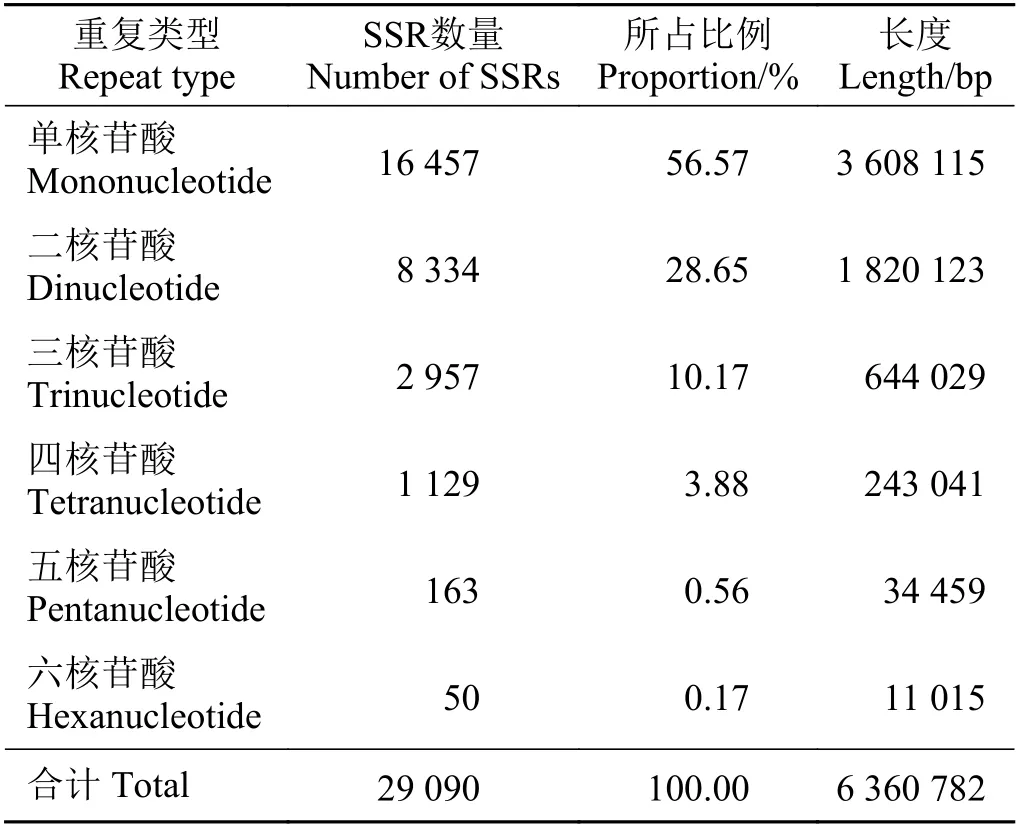
表1 高原鼢鼠轉錄組微衛星位點重復次數及長度Table 1 Repeats and length analysis of SSRS microsatellite loci in the transcriptome of the plateau zokor
2.2 轉錄組微衛星優勢重復單元堿基組成
在高原鼢鼠轉錄組29 090 個微衛星位點中發現重復堿基類型共有227 種,其中單堿基、二堿基、三堿基、四堿基、五堿基、六堿基重復分別有2、8、29、91、55 和42 種。其中四堿基重復類型最多。轉錄組SSR重復次數在5~64 次,單核苷酸重復次數種類最多,為10~64 次,二核苷酸重復次數分布在6~37 次。從合計看,在高原鼢鼠SSR 重復單元中,重復次數最多的是10 次,有5 780 個,占19.87% (表2)。單堿基重復A/T 為重復次數最多的SSR 類型,占總數的54.31%,CA/TG 是二堿基重復的主要重復單元,共2 629 個,8 種類型的二堿基重復類型占微衛星重復的36.53%,GCC/GGC 和CAAA/TTTG 分別是三堿基、四堿基的主要重復單元,分別有343 和103 個,占總數的1.18%和0.35% (圖1),而五堿基、六堿基重復單元由于數量較少、分布松散,分別只有163、50 個,各重復類型分布較為均勻,由于其相應的SSR 總數較少,且每種基元中均只有1 個SSR,故不作比較。
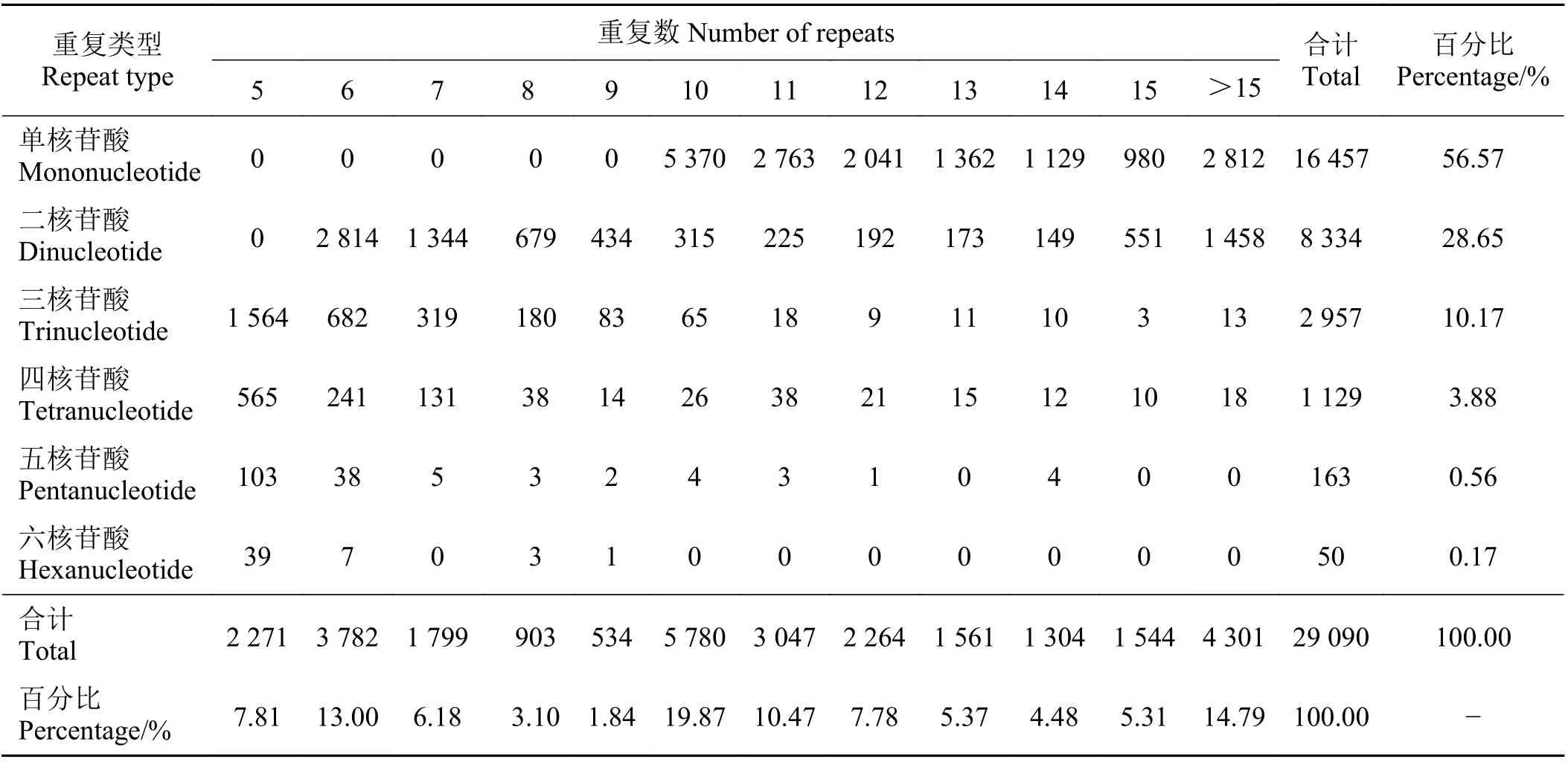
表2 基于高原鼢鼠轉錄組數據的不同類型微衛星重復數量統計Table 2 Repeated number statistics of different types of SSRS based on the transcriptome data of the plateau zokor
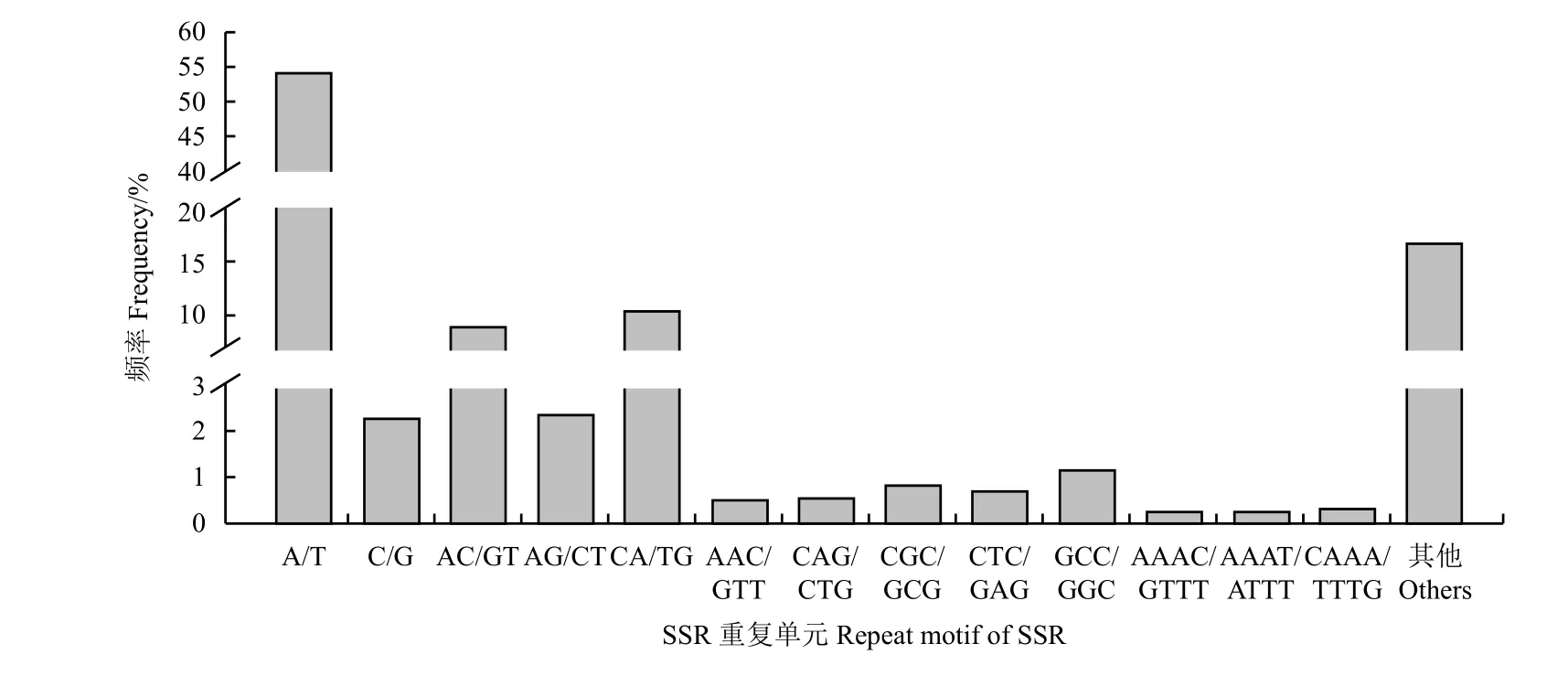
圖1 基于重復類型的微衛星分布Figure 1 Distribution of microsatellites based on repeat types
2.3 多態性微衛星位點篩選
從高原鼢鼠轉錄組中隨機篩選出104 個符合引物設計要求的EST—SSR 位點進行引物合成,其中重復微衛星數量以四堿基最多,有52 個,二堿基為43 個,三堿基為 18 個。以63 個高原鼢鼠基因組DNA 為模板進行PCR 擴增和毛細管電泳檢測,結果顯示有12 對引物能夠在高原鼢鼠個體肌肉組織DNA 中成功擴增,表現出特異性擴增產物且大小符合,擴增成功且在這些個體的擴增產物中均可找到2 個以上的等位基因,即呈現出多態性。對12 對SSR 引物在63 個高原鼢鼠個體中的擴增片段進行統計(表3)得出,共獲得76 個等位基因, 每個位點產生等位基因數Na 為3~12 個,Ho 為0.263~0.937,平均值為0.651。He 為0.285~0.844,平均值為0.600。PIC 在0.251~0.816,其中有7 個微衛星位點表現為高度多態性,5 個位點為中度多態性。使用Bonferroni校正后,有2 個微衛星位點顯著偏離哈迪—溫伯格平衡(P< 0.01),10 個微衛星位點未觀察到顯著的連鎖不平衡(P> 0.01)。
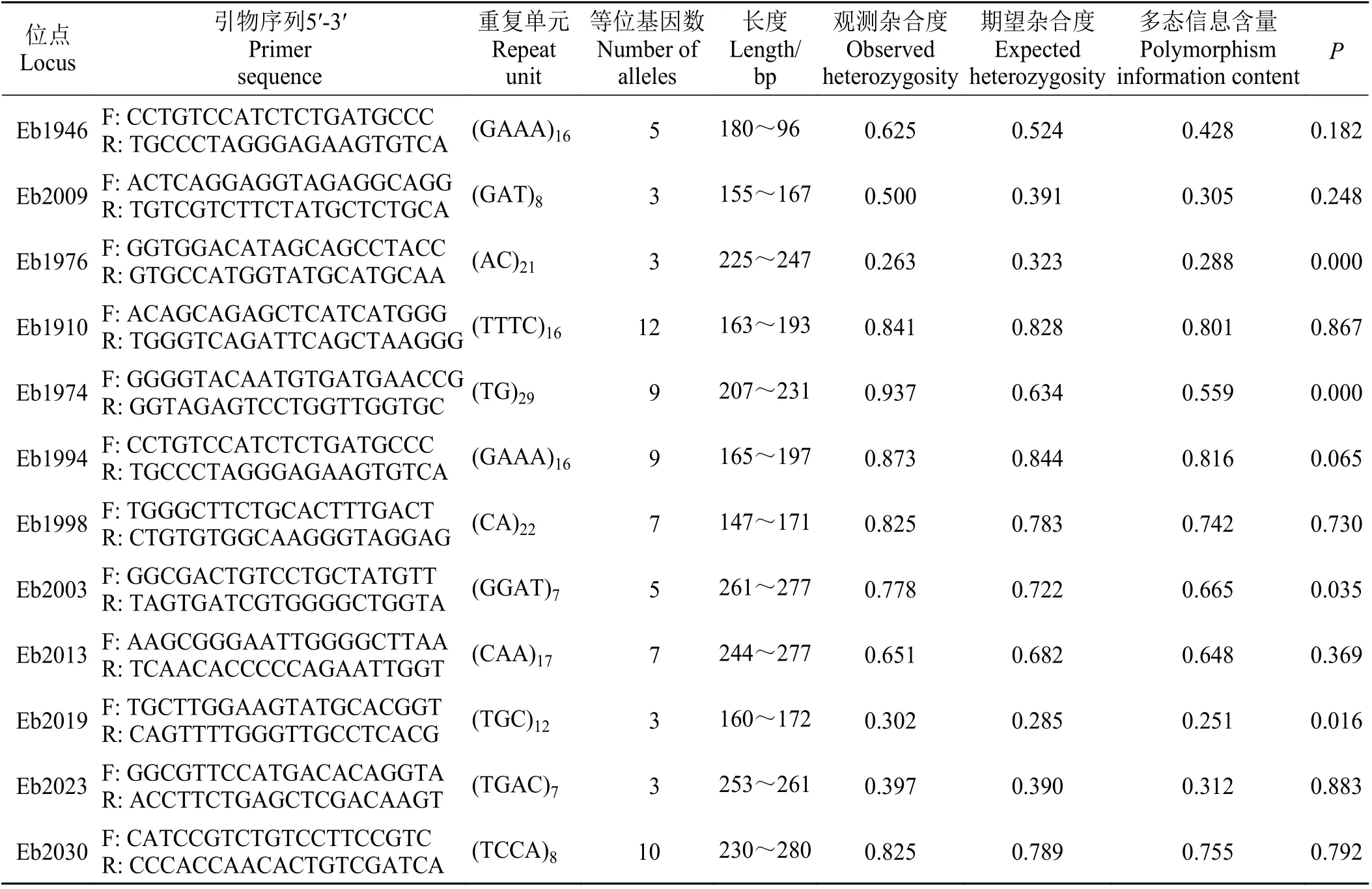
表3 12 對高原鼢鼠微衛星位點的遺傳特征Table 3 Analysis of genetic characteristics of 12 pairs of Eospalax baileyi microsatellite loci
3 討論
高通量測序技術的快速發展使得SSR 標記的開發變得簡單快捷,利用微衛星分子標記技術進行物種種群遺傳學研究已得到廣泛應用[17]。目前,微衛星位點開發主要是基于基因組文庫和轉錄組文庫進行篩選的[18],通過基因組數據可有效地分析SSR 的序列特征和功能,如涂飛云等[19]對大鼠(Rattus norvegicus)基因組各部分微衛星位點的分布特點進行了分析,為大鼠全基因微衛星研究積累了寶貴的資料;馮丹丹等[20]測定了長爪沙鼠(Meriones unguiculatus)全基因組數據,從中篩選出了357 個符合標準的微衛星位點,結果有135 個擴增成功。轉錄組測序是第二代高通量測序技術一個發展較為迅速的重要應用[21],對于缺失基因組數據的高原鼢鼠來說,利用篩選效率高、工作量較小的轉錄組數據進行微衛星位點的篩選是一種有效的方法。本研究共發現29 090 個高原鼢鼠的微衛星,其發生頻率為25.12%,與以往學者關于密斑刺鲀(Diodon hystrix)轉錄組微衛星位點的發生頻率結果相近[22],但遠遠高于已報道的斑點叉尾鮰(Ictalurus punctatus)[23]、黃姑魚(Nibea albiflora)[24]等物種,因此微衛星位點的分布頻率還因物種和組織不同而不同。轉錄后的unigene 是通過剪接產生的,有些微衛星其重復單元可能是內含子剪除后,兩側外顯子拼接而成的,其本身在基因組DNA 上并不是微衛星[25],其多態性可能是落后的分型技術產生的。現階段最常用于微衛星分型的方法有聚丙烯酰胺凝膠(polyacylamide gel electrophoresis,PAGE)和毛細管熒光電泳(capillary electrophoresis,CE),與聚丙烯酰胺凝膠電泳相比較,毛細管電泳準確性更高、速度更快且儀器操作可自動化[26]。因此本研究采用高靈敏度的毛細管熒光電泳來檢測每個位點的多態性。根據轉錄組微衛星測序結果,在高原鼢鼠中微衛星重復單元以單核苷酸重復最多,原因可能是A/T 含量高的序列退火溫度較低,易導致DNA 解鏈造成微衛星序列滑動錯配的可能性增加;其次為二核苷酸重復及三核苷酸重復類型,呈現隨著堿基數增加微衛星數量逐漸減少的現象,該結果與大熊貓(Ailuropodamelanoleuca)、興國紅鯉(Cyprinus carpiovar.singuonensis)等的轉錄組微衛星組成情況相似[27-28],但不同物種的微衛星堿基重復類型仍存在種屬間差異。
高原鼢鼠營地下獨居生活,穩定的生活環境和有限的遷移能力使得該物種的變異性較小,進化潛力較弱,擴散能力有限[29]。與地面活動的嚙齒類動物相比,高原鼢鼠種群結構的特殊性在生態系統中占據著不可替代的地位,引起了國內外學者的廣泛關注,迫切需要有效的技術手段對其遺傳資源進行開發利用。目前對于高原鼢鼠種群生態學方面的研究,大多數均從生物學特性入手,研究其在高寒草甸生態系統中的作用及其分布與防治措施,而對于研究高原鼢鼠種群遺傳分化、種群間的擴散特點等眾多問題還需要有效的分子生物學手段[30]。唐利洲等[8]以線粒體細胞色素b基因作為分子標記,對不同地理種群間的高原鼢鼠個體進行研究,結果顯示其種群間存在嚴重的限制性基因流。隨后蔡振媛等[11]測定了采自青藏高原東部不同地理種群高原鼢鼠個體的線粒體基因組序列變異情況,發現主要是地理隔離所導致的種群間遺傳分化,并且地理隔離對高原鼢鼠的作用大于甘肅鼢鼠(Eospalax cansus)。目前,微衛星標記已成功應用于高原鼢鼠的種群遺傳多樣性分析中,劉麗等[31]利用10 個中度多態性的微衛星位點,探討不同地理種群高原鼢鼠的遺傳結構,研究發現對種群個體信息交流起到阻礙作用的主要是河流和公路。在Su 等[13]和康宇坤等[14]的研究中,二者利用了高原鼢鼠近緣種鼴形鼠科和甘肅鼢鼠中已開發出的微衛星位點,在高原鼢鼠中進行了跨種擴增,有部分微衛星位點呈現出多態性,為高原鼢鼠微衛星標記的運用奠定了基礎。但高原鼢鼠僅有上述的微衛星標記還遠遠不夠,本研究篩選出的104 個轉錄組微衛星位點中僅成功擴增出12 個條帶清晰且具有多態性的微衛星位點,大部分微衛星位點表現為單態性,原因可能是轉錄組微衛星主要分布于外顯子上,其多態性受到選擇壓力及突變等的影響要比其他區域的微衛星低,因而多態性較低[32]。群體的遺傳多樣性可以通過等位基因數、雜合度和多態信息含量來反映[33]。本研究篩選出12 對引物能夠產生特異性擴增產物且均呈現出中度及高度多態性,表明該群體處于中高度多態性,與Li 等[34]在高原鼠兔(Ochotona curzoniae)及Ingram等[35]在常年營地下生活的裸鼴鼠(Heterocephalus glaber)中篩選的微衛星位點相比,大多數位點的等位基因數量較少,雜合度水平較低,且有2 個位點顯著偏離哈迪—溫伯格平衡,原因可能是所選高原鼢鼠群體較小,大多數等位基因差異存在于群體間和區域間,特殊地下生境使得群體內變異較小,導致近交,也有可能是由于無效等位基因的存在,被視為純合子,增加了純合子比例[36]。
本研究基于高原鼢鼠轉錄組測序數據開發出的12 對微衛星標記為進一步尋找更多、更有效的高原鼢鼠高度多態性微衛星位點提供了合適的工具,為進一步研究高原鼢鼠種群內部親緣關系及遷移擴散等生態學問題奠定了基礎,為高原鼢鼠種群遺傳學和分子生態學等相關研究提供了基礎數據。
