青稞HvtAGO1基因的克隆及其在條紋病脅迫下的表達
2021-03-10 08:21:50姚曉華安立昆姚有華白羿雄吳昆侖
西北植物學報 2021年1期
姚曉華,王 越,安立昆,姚有華,楊 雪,白羿雄,吳昆侖*
(1 青海大學, 西寧 810016;2 青海省農林科學院, 西寧 810016;3 青海省青稞遺傳育種重點實驗室/國家麥類改良中心青海青稞分中心, 西寧 810016)
青稞(HordeumvulgareL. var.nudumHook. f.)為禾本科大麥屬,是栽培大麥的一個變種,因其成熟時籽粒內外稃與穎果分離,籽粒裸露,故又稱裸大麥[1]。公元5世紀以來,青稞是藏族同胞的主糧,在青藏高原被廣泛種植,為藏區的糧食安全提供了重要保障[2-3]。青稞籽粒含有豐富的營養成分和生理活性物質,因此具有豐富的營養價值和醫藥保健作用[4-5]。
條紋病是青稞生產中一種常見的種傳病害,主要由麥類核腔菌[Pyrenophoragraminea(Rabenh.) Ito et Kurib.]引起,屬于真菌性病害[6]。條紋病害一旦發生會使大麥產量大幅度降低,易感品種產量可降低92%[7];2012年青海省青稞條紋病發生面積5.2×104hm2,占播種面積的60%,發病率在15%~20%,重者達50%,條紋病嚴重影響著青稞的產量與品質[8]。目前條紋病的主要防治方法是種子包衣[9],但青藏高原地區生態環境脆弱,嚴控農藥化肥,種子包衣防治難以推行,因此抗病品種的選育是防控條紋病最有效、最環保的方式。大麥條紋病抗性是一個復雜的性狀,涉及許多抗病基因[10]。數量性狀位點(QTL)分析是大麥抗病育種中獲得關鍵性狀基因的有效方法[11-12]。例如,將QTL分析應用于抗條紋病和易感大麥品種雜交后代,鑒定出Rdg1a和Rdg2a兩個抗病基因,分別位于2HL和7HS染色體上[12-15],其過表達顯著增強了大麥對條紋病的抗性[16-19]。然而,通過QTL分析,只發現了少數抗條紋病基因。說明通過傳統的基因定位獲得抗條紋病基因可能遇到了瓶頸,因此需要新的方法和視角來探索與條紋病抗性相關的植物新基因。
Argonaute(AGO)蛋白是核糖核酸介導的轉錄后基因調控復合物的重要蛋白,最初在擬南芥中發現[20]。該蛋白主要由可變N端、PAZ(PIWI-Argonaute-Zwille)、PIWI(Pelement-induced wimpy testis)和MID(Middle)4個結構域組成,有的還有DUF1785(Domains of unknown function protein 1785)和AgoL2(Argonaute linker 2)兩個結構域,其中PAZ 結構域和C末端的PIWI 結構域為2個標志性的功能域[21-22]。研究表明,AGO蛋白可以與不同類型的小RNA結合,在植物的抗病中發揮重要作用。番茄miR403可以通過靶向AGO2基因調控葉縮短病[23];在水稻中,病毒誘導的OsAGO18能夠抑制miR168,從而減輕miR168對水稻OsAGO1的抑制,使感染RSV病毒(Ring Sopt Virus)的水稻具有抗病毒防御能力[24];在煙草中,AGO1的差異表達能夠改變植物對病毒感染的耐受能力,使遭受病毒侵染的煙草不僅恢復性狀,同時還增強了冷脅迫的耐受能力[25]。
本研究通過分析不同抗性青稞品種條紋病感染后的轉錄組測序結果,鑒定出一個差異表達的AGO基因,在Swiss-Prot中被注釋為AGO1基因。在抗病品種‘昆侖14號’和感病品種‘Z1141’中克隆了該基因,分析了其序列的結構、生理生化特性以及與其他禾本科植物的進化關系,利用psRNATarget在線工具預測了調控該基因的miRNA(hvu-miR168-5p),并利用降解組測序驗證了兩者的靶向關系。利用實時熒光定量PCR分析了HvtAGO1基因和調控其表達的hvu-miR168-5p在青稞條紋病脅迫下的響應模式,以期為進一步研究HvtAGO1基因的抗條紋病功能和抗病機理奠定理論基礎。
1 材料和方法
1.1 材 料
抗病青稞品種‘昆侖14號’和感病青稞品種‘Z1141’由青海大學農林科學院作物栽培與育種研究所青稞研究室提供。‘昆侖14號’條紋病發病率<10%,‘Z1141’條紋病發病率>90%。
1.2 方 法
1.2.1 青稞葉片總RNA的提取及cDNA合成利用植物總RNA提取試劑盒(天根)提取青稞品種‘昆侖14號’和‘Z1141’的總RNA,用超微量核酸蛋白測量儀(NanoPhotometer)測定RNA的濃度和純度,以1.5%瓊脂糖凝膠電泳檢測質量。參照第一鏈cDNA合成試劑盒PrimeScript 1st Strand cDNA Synthesis Kit(TaKaRa)合成cDNA,-80 ℃保存。
1.2.2 青稞HvtAGO1基因的克隆從‘昆侖14號’和 ‘Z1141’ 青稞品種條紋病感染前后差異表達的轉錄組測序(RNA-seq)結果中獲得1個差異表達的AGO基因(Gene IDHORVU7Hr1G007000),利用Primer5.0設計該基因引物(表1)。以青稞‘昆侖14號’和‘Z1141’葉片cDNA 為模板進行PCR擴增(東洋紡試劑盒)。擴增體系為25 μL,其中2×PCR Buffer for KOD Fx緩沖液12.5 μL、上下游引物各0.5 μL(10 μmol·L-1)、dNTPs 5 μL(2 mmol·L-1)、KOD酶0.5 μL(1.0 U·μL-1)、cDNA模板1 μL、ddH2O補足至25 μL。反應程序:94 ℃預變性 2 min;98 ℃變性 10 s,56 ℃退火 30 s,68 ℃延伸40 min 30 s,35個循環;72 ℃延伸10 min,最后4 ℃ 保持5 min。乙醇沉淀法回收目的條帶,用超微量核酸蛋白測量儀測定回收產物的濃度和純度,再將其連接到pEasy-Blunt載體(全式金生物)上,轉化至大腸桿菌Trans-T1感受態細胞。挑取5個白斑單菌落進行陽性克隆鑒定,至少3個陽性克隆送至上海生物工程股份有限公司,使用通用引物M13測序。
1.2.3 青稞HvtAGO1基因生物信息學分析利用Expasy Protparma(http://www.expasy.org/tools/protp aram.html)和Protscale(https://web.expasy.org/protscale/)預測蛋白質的理化性質和親/疏水性。利用SignalP4.1(http://www. Detaibio.com/tools/signal-peptide.html)和TMHMM-2.0(http://www.cbs.dtu.dk/services/TMHMM-2.0/)預測蛋白質的信號肽和跨膜結構。利用CELLO 2.5 (http://cello.life.nctu.edu.tw)對HvtAGO1蛋白進行亞細胞定位預測。利用SMART(http://smart.embl-heidelberg.de/)預測結構域。利用SPOMA(https:// npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_sopma.html)和SWISS-MODEL(https://swissmodel.expasy.org/)預測HvtAGO1蛋白的二級結構和三級結構。利用SignalP 4.1(http://www.detaibio. com/tools/signal-peptide. html)和TMHMM-2.0(http://www. cbs. dtu. dk/services/TMHMM-2.0/)預測蛋白質的信號肽和跨膜結構。利用NCBI中的Blastp功能,查詢與HvtAGO1蛋白同源的禾本科其他物種AGO1蛋白序列,用DNAMAN6.0軟件進行多序列比對,用Mega5.1軟件構建系統進化樹。
1.2.4 調控HvtAGO1基因的miRNA鑒定利用psRNATarget(http://plantgrn.noble.org/psRNATarget/analysis?function=1)預測調控HvtAGO1基因的miRNA。利用降解組測序驗證miRNA與HvtAGO1基因的靶向關系。

表1 引物序列
1.2.5HvtAGO1基因及其調控miRNA在青稞條紋病脅迫下的表達分析采用Primer5.0設計hvu-miR168-5p和HvtAGO1的qRT-PCR引物,利用實驗室建立的青稞條紋病侵染體系,獲得條紋病侵染20 d后‘昆侖14號’和‘Z1141’正常葉片和感病葉片[26]。提取葉片總RNA,反轉錄后以cDNA(200 ng·μL-1)為模板,分別以5SrRNA和TC139057為內參進行實時熒光定量PCR(qRT-PCR)分析,引物序列見表1。反應體系為20 μL,其中TB Green premix Ex Taq Ⅱ(Tli RNaseH Plus)(2×)10 μL,cDNA模板2 μL、(10 μmol·L-1)各0.8 μL和ddH2O 6.4 μL。qRT-PCR擴增條件:95 ℃預變性5 min; 95 ℃變性5 s, 60 ℃退火30 s,72 ℃延伸30 s,循環40次;溶解曲線,95℃ 5 s,70 ℃ 1 min。應用2-ΔΔCt法計算基因相對表達量[27]。每個樣品設3次生物學重復。
2 結果與分析
2.1 青稞HvtAGO1基因的克隆與序列分析
以‘昆侖14號’和‘Z1141’的葉片總RNA反轉錄的cDNA為模板,HvtAGO1-F/R為引物,擴增到一條約3 675 bp的目的條帶(圖1)。其完整開放閱讀框為3 651 bp,編碼1 217個氨基酸(GenBank 登錄號 MW387021)。測序后比對,‘昆侖14號’和‘Z1141’的堿基序列及其編碼的氨基酸序列相似性均為100%。利用SMART對其氨基酸序列進行保守結構域預測,發現該基因具有典型的 PIWI、PAZ和DUF1785結構域,同時具有Gly-rich-ArgoN(Glycine-rich region of argonaut)、ArgoN(N-terminal domain of argonaute)、ArgoL2 (Argonaute Linker 2)和ArgoMid(Mid domain of argonaute)結構域,屬于AGOs 基因家族(圖2)。
蛋白質理化性質分析表明,HvtAGO1蛋白分子式為C5861H9195N1785O1746S37,分子量為133.79 kD,不穩定指數為50.90,脂溶指數是65.90,理論等電點為9.49,其中負電荷殘基(Asp + Glu) 為112個,正電荷殘基(Arg + Lys)為149個。平均疏水性(GRAVY)為-0.694。預測結果表明,HvtAGO1蛋白是一個親水性的不穩定堿性蛋白。對HvtAGO1蛋白跨膜結構與信號肽預測發現,該蛋白不存在跨膜結構,且無信號肽。CELLO V.2.5分析表明,HvtAGO1蛋白可能定位在細胞周質中,得分為1.492。該蛋白定位到細胞結構的可能性:細胞周質(1.492)>細胞外(1.165)>外膜(0.818)> 細胞質(0.802)>內膜(0.722)。

圖2 HvtAGO1基因結構域預測Fig.2 Domain prediction of HvtAGO1 gene

藍色表示α-螺旋;紅色表示延伸鏈;綠色表示β-轉角;橙色表示無規則卷曲; 紫色表示HvtAGO1蛋白氨基酸長度圖3 HvtAGO1蛋白二級結構預測Blue stands for α-helix; Red stands for extended chain; Green stands for β- turn; Orange stands for randon coil; Purple stands for the amino acid length of HVTAGO1 proteinFig.3 Secondary structure prediction of HvtAGO1
對HvtAGO1蛋白的二級結構預測結果表明:HvtAGO1蛋白的二級結構主要是由無規則卷曲、α-螺旋、延伸鏈、β-轉角組成,其中無規則卷曲占56.41%,α-螺旋占25.74%,延伸鏈占13.40%,β-轉角占4.44%(圖3),可見該基因是以無規則卷曲和α-螺旋為主,這2個二級結構在蛋白行使功能時可能發揮重要作用。用SWISS-MODEL對HvtAGO1蛋白進行三級結構預測,由圖4可知,HvtAGO1蛋白的功能結構域折疊成錘子狀,主要由無規則卷曲和α-螺旋構成,錘身包含了PIWI特征結構域;錘柄主要包含PAZ特征結構域。Unipro基因注釋結果表明該蛋白為AGO1,因此將該基因命名為HvtAGO1。

圖4 HvtAGO1蛋白三級結構預測Fig.4 Tertiary structure prediction of HvtAGO1

Hvt. 青稞;Hv. 大麥;Bd. 二穗短柄草;Si. 粟;At. 山羊草; Tu. 烏拉爾圖小麥;Zm.玉米;黑線表示DUF1785結構域;紅線表示PAZ結構域;綠線表示PIWI結構域圖5 HvtAGO1與其他禾本科植物蛋白和DUF1785、PAZ和PIWI結構域序列比對Hvt. Hordeum vulgare(Tibetan hulless barley);Hv. Hordeum vulgare;Bd. Brachypodium distachyon;Si. Setaria italica;At. Aegilops tauschii;Tu.Triticum urartu;Zm. Zea mays;The bold black frame represents NB-ARC conserved functional region of N/A protein;The black line represents DUF1785 conserved domain; The red line represents PAZ conserved domain; The green line represents PIWI conserved domainFig.5 Multiple alignment of HvtAGO1 and three domains (DUF1785, PAZ and PIWI) compared with other Gramineae plants
2.2 HvtAGO1蛋白的同源比較及系統進化分析
圖5顯示,青稞HvtAGO1蛋白與山羊草(Aegilopstauschii)、二穗短柄草 (Brachypodiumdistachyon)和烏拉爾圖小麥(Triticumurartu)等的AGO1蛋白序列相似性分別為96.64%、79.34%和80.07%,這些序列都具有高度保守的DUF1785、PAZ和PIWI結構域。系統進化樹表明,在7種禾本科植物種中,青稞HvtAGO1蛋白與山羊草的親緣關系最近,與玉米和粟的親緣關系較遠(圖6)。

圖6 HvtAGO1蛋白與禾本科其他植物系統進化樹Fig.6 Phylogenetic tree analysis of the HvtAGO1 protein and other Gramineae plants

A. hvu-miR168-5p剪切靶基因HvtAGO1的位置;B.hvu-miR168-5p剪切靶基因HvtAGO1的靶點鑒定;在A中HvtAGO1下方箭頭和B中圓點表示hvu-miR168-5p指向的裂解位點圖7 hvu-miR168-5p剪切靶基因HvtAGO1的位置和靶點鑒定1.Position of hvu-miR168-5p shearing target gene HvtAGO1; B. Target identification of hvu-miR168-5p shearing target gene HvtAGO1; The arrow below HvtAGO1 in A and the dot in B indicated hvu-miR168-5p-directed cleavage siteFig.7 Position and target identification of hvu-miR168-5p shearing target gene HvtAGO1
2.3 調控HvtAGO1基因的miRNA的鑒定
利用在線工具預測調控HvtAGO1基因的miRNA為hvu-miR168-5p;同時降解組測序結果證明,HvtAGO1確實為hvu-miR168-5p的靶基因,其剪切位置為HvtAGO1基因的832 bp處(圖7)。由此說明hvu-miR168-5p和靶基因HvtAGO1存在靶向作用關系。
2.4 hvu-miR168-5p和靶基因HvtAGO1在條紋病脅迫下的表達
利用實時定量PCR檢測了HvtAGO1和調控其表達的hvu-miR168-5p在條紋病脅迫下的表達模式。結果(圖8)表明,抗病青稞品種‘昆侖14號’和感病青稞品種‘Z1141’在條紋病菌感染后,hvu-miR168-5p和其靶基因HvtAGO1表達量均極顯著上調(P<0.01)。其中,病菌侵染后hvu-miR168-5p在‘昆侖14號’葉片表達量為未侵染的42.24倍,而‘Z1141’為未侵染的7.21倍;病菌侵染后HvtAGO1在‘昆侖14號’葉片的表達量為未侵染的14.37倍,‘Z1141’為未侵染的5.08倍。且病菌侵染后,hvu-miR168-5p和靶基因HvtAGO1在‘昆侖14號’葉片的表達量均極顯著高于‘Z1141’(P<0.01)。推測hvu-miR168-5p和靶基因HvtAGO1在青稞抗條紋病過程中發揮重要作用。
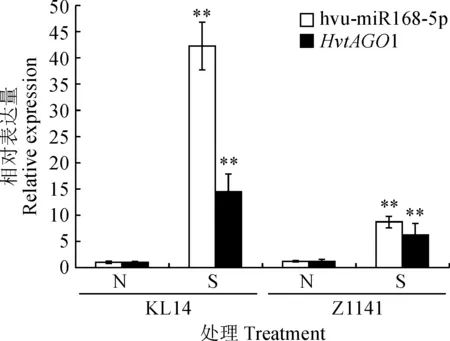
N.正常葉片;S.感病葉片;**表示同一品種同一基因的感病葉片與正常葉片差異極顯著 (P <0.01)圖8 不同抗性青稞品種hvu-miR168-5p和靶基因HvtAGO1在條紋病脅迫下的表達N. Normal leaves; S. Leaves with barley leaf stripe; ** stands for significant difference between leaves with barley leaf stripe and normal leaves within same variety and gene at 0.01 levelFig.8 Expression levels of hvu-miR168-5p and target gene HvtAGO1 in different resistant Tibetan hulless barley infection with barley leaf stripe
3 討 論
近年來,AGOs家族基因已經從擬南芥、蘋果、玉米、水稻等多種高等植物中被分離鑒定出來[28-31]。RNA干擾在植物的生長發育過程中發揮重要的作用,在水稻和玉米中已經證實,AGO蛋白是已知的小RNA調控通路的核心部分,通過抑制靶基因的表達來調控發育過程[30-31]。然而,青稞HvtAGOs相關基因序列和抗病功能目前還沒有相關研究。本研究從青稞抗病品種‘昆侖14號’和 感病品種‘Z1141’條紋病感染前后轉錄組差異表達的基因注釋中獲得的一個HvtAGO1基因,進一步對其進行了克隆和序列分析。已有研究表明,AGO1 參與的通路包括miRNA 通路、轉錄后基因沉默(PTGS)通路、病毒誘導的基因沉默(VIGS)或抗病毒沉默通路和ta-siRNA 通路[28]。AGO1主要結合miRNA在細胞質中通過切割靶mRNA或介導翻譯抑制在轉錄后水平抑制靶基因表達[32]。在擬南芥中,AtAGO1 能與病毒siRNA結合形成RNA沉默復合體,在抗病中發揮重要作用[33]。因此推測,HvtAGO1編碼的蛋白在青稞中的作用與同類AGO1蛋白在其他植物中的作用相似。
前人研究表明,AGO蛋白包含高度保守的結構域,如DUF1785、PAZ和PIWI,這些結構域在結合sRNA雙鏈和裂解靶RNA中起重要作用[34-36]。本研究從青稞分離的與條紋病相關的HvtAGO1蛋白與山羊草、二穗短柄草、小麥等6個禾本科植物的AGO1蛋白序列相似性為87.05%,其中DUF1875、PAZ和PIWI結構域相似性分別為97.57%,98.44%和96.02%。可見,7個AGO1蛋白雖然相似性不太高,但3個結構域相似性均大于95%,結構域的較高保守性也說明了他們在特異地與小RNA 結合進行目標靶基因切割時,可能存在基因功能的一致性[37]。另外發現,青稞與大麥的AGO1蛋白序列相似性僅為82.69%,青稞與大麥相比,在70~137 bp和185~234 bp產生片段插入,這2個片段的插入導致兩者在進化上產生距離,是否會導致功能差異,還有待進一步研究。
2010年Kantar等[38]首次從大麥中分離出miR168,之后又有學者從干旱、鎘、鋁和鹽脅迫下的水稻[39]和黃瓜[40]中分離出miR168。利用轉基因擬南芥進行的啟動子分析顯示,真菌可以激活miR168,而且能夠上調控制葉片形態和植物生長[41]。在番茄卷曲葉病毒感染期間也發揮抗病作用[24]。本研究中,青稞條紋病感染前后的降解組測序結果中,HvtAGO1被miR168-5p剪切。研究表明,miR168能夠調控AGO1基因的表達,AGO1作為RNA誘導沉默復合物(RNA induced silencing complex, RISC)中轉錄調控或/和轉錄后調控的核心元件,負責轉錄后基因沉默,參與植物發育過程和病毒防御[41]。本研究在證明了HvtAGO1為hvu-miR168-5p的靶基因后,利用qRT-PCR表達分析發現,青稞葉片被條紋病菌侵染后hvu-miR168-5p被激活,且抗病品種表達量顯著高于感病品種,推測該miRNA在青稞抗條紋病過程中發揮重要作用,能夠正向調控條紋病抗性。同時,靶基因HvtAGO1在受青稞條紋病侵染后,葉片中的表達與hvu-miR168-5p呈相同的表達趨勢,這與被RMV、CMV和TCV感染的擬南芥中miR168和AtAGO1的表達量總是與病毒感染過程的進展同時增加的結果類似[42]。HvtAGO1和hvu-miR168-5p表達水平的同時升高可能是兩者均具有協同表達調節機制[43]。但兩者在青稞抗條紋病過程中的具體調控機制有待進一步研究。
