蘋果MAX1基因的克隆和表達及啟動子分析
2021-03-10 01:40:38高馨怡常興敏孫怡婷謝玲玲馬娟娟
西北植物學報 2021年1期
關鍵詞:植物
高馨怡,陳 皓,程 方,常興敏,孫怡婷,謝玲玲,馬娟娟
(西北農林科技大學 園藝學院,陜西楊陵 712100)
獨角金內酯(strigolactones/SLs)因能誘導根寄生植物種子萌發和叢枝菌根真菌(AMF)菌絲分枝而受關注和研究[1],2008年被判定為第七大類植物激素,SLs作為植物激素首先被研究的功能與植物分枝相關[2-3]。分枝是植物株型重要構成方面之一,影響著植株對光照和養分等資源的獲取[4]。植物分枝受激素、營養、發育階段、環境因素及其之間復雜相互作用的調節[5-6]。生長素(吲哚-3-乙酸,IAAs)、細胞分裂素(CTKs)和SLs是控制分枝的重要激素,由類胡蘿卜素類物質產生的SLs可能作為生長素第二信使參與分枝生長[7]。
豌豆中的rms1/5(ramosus)突變體、水稻中d10/17(dwarf)突變體、擬南芥中的max3/4(moreaxillarygrowth)突變體均具有多分枝表型,液相串聯質譜測定后為SLs缺失型,外源施用SLs能抑制其分枝的產生[2-3],相互嫁接試驗證明SLs能從根部合成并向上運輸到芽進而抑制腋芽的生長[8]。
目前SLs的生物合成途徑尚不完全清楚,但擬南芥中至少有5種酶的參與。DWARF27(D27)、MAX3、MAX4、MAX1和LATERALBRANCHINGOXIDASE(LBO)分別編碼類胡蘿卜素異構酶、類胡蘿卜素裂解雙加氧酶 7(CCD7)、類胡蘿卜素裂解雙加氧酶 8(CCD8)、細胞色素P450單加氧酶和側枝雙加氧酶[9-12]。D27、CCD7和CCD8等3種酶依次將β-胡蘿卜素轉化為己內酯(carlactone,CL)。擬南芥MAX1/CYP711A1將CL轉化為己內酯酸(carlactonoic acid,CLA)[11],水稻中有5個MAX1的同源基因,其中編碼產物Os900/CYP711A2將CL轉化為 4- deoxyorobanchol (4DO),Os1400/CYP711A3將4DO轉化為列當醇(orobanchol)[13]。MAX1s根據催化產物的不同分為3種類型:A1型,催化CL產生CLA;A2型,催化CL產生4DO;A3型,催化CL產生列當醇;不同物種中MAX1保守功能是催化CL產生CLA[14]。
SLs在分枝調控中具有重要作用,MAX3/4已被證明對蘋果分枝有影響[15]。蘋果是一種世界各地廣泛栽培的果樹,具有重要的經濟價值。保持理想的分枝構型,以最大限度利用光能是提高產量的關鍵因素之一,因此,研究蘋果腋芽分枝生長相關基因對蘋果育種具有重要意義。擬南芥max1突變體具有多分枝表型,矮牽牛、水稻、楊樹、番茄和油菜的MAX1同源基因均被證明具有控制分枝功能[16-21]。為此,本研究鑒定了蘋果MAX1的同源基因,并進行了系統分析及研究,為將來進一步研究蘋果MAX1參與SLs合成調控蘋果分枝的功能提供依據。
1 材料和方法
1.1 實驗材料
‘長富2號’/T337/八棱海棠(MalusrobustaRehd.)嫁接苗于陜西省楊陵區西北農林科技大學武功站實驗苗圃(34°52′N,108°7′E)進行種植和管理,種植間距為50 cm×25 cm。基因克隆采樣部位為腋芽;不同節位的腋芽自頂芽以下依次編號1~6。組織特異性采樣部位為根(八棱海棠砧木所有根系混樣)、莖(靠近生長點處,粗度達到3~4 mm左右)、頂梢(生長點1~2 cm處)、葉和腋芽(不同節位腋芽混樣),開放花(盛花期)和幼果(直徑約1 cm左右)采集于5年生‘長富2號’/T337/八棱海棠蘋果樹。
5周齡本氏煙草(Nicotianabenthamiana)種植于西北農林科技大學園藝學院實驗室培養箱。
1.2 實驗方法
1.2.1 MdMAX1的鑒定與系統發育分析采用同源比對法,以擬南芥MAX1(AtMAX1)蛋白序列為參照通過GDR(www.rosaceae.org)blastp尋找蘋果同源序列。利用2個單子葉植物和7個雙子葉植物MAX1蛋白序列,使用MEGA 5.0軟件構建系統發育樹,構建參數為鄰接樹法,p-distance、Bootstrap=1 000。
1.2.2 保守motif分析將構建系統發育樹所用的MAX1蛋白質序列提交到MEME(http://meme-suite. org/tools/meme),并將motifs的參數設置為15。
1.2.3 基因克隆和實時熒光定量PCR將‘長富2號’嫁接苗不同組織(根、莖、開放花、幼果、頂芽、葉、腋芽)及不同節位腋芽分別提取總RNA。使用SYBR Prime Script RT-PCR Kit Ⅱ (TaKaRa, 中國上海)進行反轉錄。利用腋芽cDNA進行MAX1的克隆,利用Primer 6.0軟件設計引物,CDS克隆引物為MdMAX1F(ATGAAGAGCTTGGAAGA-AGAG)和MdMAX1R(TGTCCTCTTTATGAC-TCTAAGCTT);定量引物為MdMAX1r-F(TCT-CGGACTTCATCAACCAACACA)和MdMAX1r-R(GCATTCGCTTCGCCACAATACC),以蘋果Actin為內參基因,使用2×Fast qPCR Master Mixture Green(DiNing, 中國北京)進行實時熒光定量PCR。
1.2.4 RNA-seq表達分析蘋果‘長富2號’嫁接苗 6-BA處理腋芽和海棠‘bly114’(Malusspectabilis)(WT)及其多分枝突變體(MB)腋芽的RNA-seq數據庫來自本實驗室已有數據[22-23],其中海棠‘bly114’及其多分枝突變體引種自青島市農業科學院。RNA-seq表達量基于RPKM(reads per kilo bases per million reads)數值。
1.2.5MdMAX1啟動子的克隆與序列分析利用GDR網站的contig MDP0000130133序列設計引物,以‘長富2號’葉片DNA為模板克隆MdMAX1 ATG前的1 500 bp序列,并將克隆得到的啟動子序列提交至PlantCARE進行順式作用元件的分析。啟動子區段克隆引物為MdMAX1p-F(TAGTTTTAAGAACACCATCCATAA)和MdMAX1p-R(ATGAATGATTTCAATTGTGGAA-AT)。
1.2.6MdMAX1啟動子活性分析采用同源重組法將MdMAX1 1 500 bp啟動子序列連接到pCAMBIA1381-GUS載體。通過農桿菌EHA105介導,侵染5周齡本氏煙草,GUS染色后進行啟動子活性分析[24]。為了解啟動子活性是否受激素和光照等處理影響,用20 μmol/L GR24(獨腳金內酯類似物)、100 μmol/L IAA(生長素)、100 μmol/L 6-BA(細胞分裂素)、100 μmol/L Lovastatin(細胞分裂素合成抑制劑)、100 μmol/L GA3(赤霉素)及光照處理48 h后,分別收集葉片進行GUS活性定量分析和組織化學染色。
1.3 數據分析
運用Excel 2019整理統計數據,使用軟件SPSS 22.0分析數據,采用單因素ANOVA的Duncan法和獨立樣本T檢驗進行比較分析, 差異顯著水平均為0.05(P<0.05),使用軟件Origin 2017 作圖。
2 結果與分析
2.1 MdMAX1的克隆和鑒定
在GDR網站以擬南芥MAX1為模板進行同源比對,蘋果中位于LG15的MDP0000130133相似度最高。以MDP0000130133 CDS序列為參考,克隆得到MdMAX1,其開放閱讀框1 620 bp,比參考序列增加了6個堿基,相似度99.44%;編碼539個氨基酸,與參考基因氨基酸序列的相似度99.07%(圖1)。系統發育樹分析顯示,MdMAX1與雙子葉植物MAX1聚類距離較近(圖2,A)。MAX1根據作用產物的不同可分為3種類型(A1~A3型),與A2類型水稻(Os900)和A3類型水稻、玉米(Os1400、ZmMAX1b)相比,MdMAX1與A1類型矮牽牛、番茄、擬南芥MAX1(PtMAX1、SlMAX1和AtMAX1)聚類關系更近。
進一步的基序分析表明,雙子葉植物MAX1都包含13個motifs,而單子葉植物motifs數目不固定(12~14個)(圖2,B)。雙子葉植物MAX1比單子葉植物MAX1都少1個固定的motif,序列為RKDFLSVVLDARESNKSSREL。此外,所有的A1型MAX1都包含13個motifs。
2.2 MdMAX1在蘋果中的表達模式
利用qRT-PCR分析‘長富2號’嫁接苗7個組織中MAX1的表達。MAX1在莖中高表達,在腋芽本身也有表達,而在開放的花、頂梢和葉片中的表達水平可以忽略不計(圖3)。MAX1在腋芽3、5中的表達水平高于其他節位的腋芽并存在顯著差異(圖3)。
RNA-seq轉錄組數據分析發現6-BA處理‘長富2號’腋芽96 h后MAX1的表達量下降至對照組的一半,達到顯著差異(圖4,A)。另外,分析‘bly114’海棠腋芽(WT)及它的多分枝突變體(MB) 轉錄組數據發現MAX1在WT和MB腋芽中表達量有顯著差異(圖4,B)。
2.3 MdMAX1基因啟動子的克隆與序列分析
以‘長富2號’葉片基因組 DNA 為模板,對MdMAX1 ATG前的1 500 bp啟動子區段進行克隆(圖5),得到的序列與GDR預測序列完全相同。使用PlantCARE在線軟件進行順式作用元件預測,MdMAX1啟動子片段上含有多個光響應原件(TCT-motif、ATC-motif、Box4和MRE),多個激素相關作用元件(涉及到的激素有茉莉酸甲酯、赤霉素和水楊酸),分別與低溫、抗逆和創傷響應相關的LTR、TC-rich repeats和WUN-motif元件,另外,還有響應厭氧誘導、分生組織特異性激活和生物節律的作用元件(表1)。
2.4 不同處理對MdMAX1啟動子活性的影響
GUS活性檢測顯示MdMAX1 ATG上游1 500 bp啟動子片段具有啟動活性,且GR24和Lovastatin處理后,MdMAX1啟動子活性增強;光照處理后,MdMAX1啟動子活性減弱;IAA、GA3和6-BA處理后,MdMAX1啟動子活性無顯著變化(圖 6)。
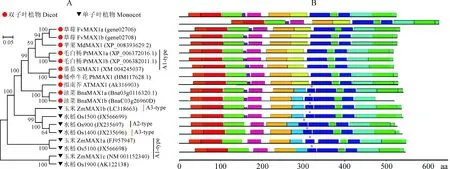
A1型MAX1將己內酯轉化為已內酯酸;A2型通過已內酯酸將己內酯轉化為4-deoxyorobancho(4DO);A3類型,將己內酯轉換為列當醇;紅色星號.單子葉植物MAX1特有的motif圖2 不同植物MAX1蛋白的系統發育(A)及motif分析(B)A1-type, converting carlactone (CL) to carlactonoic acid(CLA); A2-type, converting CL to 4-deoxyorobanchol(4DO) via CLA; and A3-type, converting CL to CLA and 4DO to orobanchol. Monocots MAX1 unique motif is marked with a red asteriskFig.2 A phylogenetic (A) and motif analysis (B) of MAX1 protein from different plant species

不同小寫字母代表不同組織間(虛線左)和不同節位腋芽間(虛線右)的顯著差異(P < 0.05)。數據為平均值±標準差(n = 3)圖3 MdMAX1在‘長富2號’嫁接苗不同組織中的表達Different lowercase letters represent significant differences between different tissues (the left part of the dashed line) and between axillary buds at different nodes (the left part of the dashed line). Data are means ± SE (n = 3)Fig.3 The expression of MdMAX1 in different tissues of ‘Nagafu 2’

A.6-BA處理后‘長富2號’腋芽中MAX1的表達;B.‘bly114’海棠(WT)和它的多分枝突變體(MB)腋芽中MAX1的表達。* 表示在0.05 水平上有顯著性差異(P <0.05),RNA-seq表達量基于RPKM(Reads per kilo bases per million reads)數值圖4 基于RNA-seq的蘋果腋芽MAX1基因表達譜分析A. Expression of MAX1 in lateral buds of ‘Nagafu 2’with 6-BA treatment; B. Expression of MAX1 in lateral buds Malus spectabilis ‘bly114’ (WT) and its more branching mutant(MB).* indicates significant difference at the 0.05 level (P <0.05). RNA-seq expression is based on RPKM (Reads per kilo bases per million reads) valueFig.4 The expression patterns of MAX1 in lateral buds of apple by RNA-seq-based transcriptome

表1 MdMAX1基因啟動子的順式作用元件

1和2為MdMAX1啟動子; M為DNA Marker圖5 蘋果MdMAX1基因啟動子的克隆1 and 2 indicated the cloned products of the MdMAX1 promoter;M indicated the DNA markerFig.5 Promoter cloning of MdMAX1 in apple
3 討 論
SLs是近年來發現的抑制腋芽生長的新型激素,有關SLs和分枝的關系許多問題還需深入研究[25]。MAX1是SLs合成途徑中的關鍵基因,MAX1在‘長富2號’和 ‘Bly114’海棠的腋芽中均有表達,‘長富2號’不同節位的腋芽中,MAX1的表達水平不同。不同節位的腋芽具有不同的分枝能力[26];SLs在豌豆腋芽內自身合成并直接作用于腋芽[27]。根是SLs合成的主要部位,但嫁接實驗表明,在莖中直接合成的SLs向上移動以抑制芽的生長可能更為重要[15,27]。蘋果腋芽中MAX1表達的豐富性是否使腋芽中合成的SLs扮演更為重要的角色有待深入研究。
MAX1編碼細胞色素P450單加氧酶,MAX1主要負責 SL 的結構修飾。植物中已發現三十多種天然的 SL,這些 SL 結構的差異化歸功于MAX1同源基因的存在。MAX1s可以分為3種類型:A1~A3型,A1型是保守的,其將CL轉化為CLA[14]。通過系統發育分析,MdMAX1與A1型MAX1s聚類接近,MdMAX1 基序數與A1型MAX1相同,因此MdMAX1可能也參與了蘋果中CL向CLA的轉變。
在植物分枝發育的復雜調控網絡中,激素和環境因子發揮著極其重要的作用。但是,內源激素和環境因子之間的相互調控關系尚不明確。光作為重要的環境因素之一,通過光強、光質和光周期等影響植物側枝伸長。高粱腋芽中 SL 信號轉導基因SbMAX2 響應遠紅光處理上調表達,抑制phyB突變體的生長會誘導SbMAX2基因的表達[28];SL調控腋芽萌發依賴于BRC1,較低的紅光/遠紅光比率處理能誘導腋芽中BRC1的表達,BRC1可以抑制與光合作用相關基因的轉錄活性,誘導與脫落酸相關基因的表達[29];在遮蔭條件下,擬南芥phyB突變體分枝減少,莖中生長素響應相關基因的轉錄水平顯著提高[30],由此推測,光可能通過植物激素(如生長素、SL 和脫落酸等)相關基因的表達調控植物的分枝發育。對MdMAX1 啟動子片段順式作用元件分析發現,蘋果MdMAX1啟動子含有多個光響應作用元件,推測光同樣調控MdMAX1的表達;另外,啟動子GUS活性分析結果顯示光照處理能顯著抑制蘋果MdMAX1啟動子的活性,表明蘋果MdMAX1 基因與光信號之間有著不可忽視的關聯,但其中的分子機制還需進行深入研究。

A.PMAX1-GUS 融合表達載體構建示意圖; B.不同處理后煙草葉片中GUS含量的定量測定(n=3),小寫字母代表不同處理間的顯著差異(P < 0.05);C.不同處理后煙草葉片的 GUS組織染色,N b.本氏煙草圖6 不同處理后蘋果MdMAX1啟動子瞬時轉化煙草的 GUS 活性分析A. Schematic diagram of PMAX1-GUS fusion expression vector; B. GUS activity of MdMAX1a promoters in tobacco leaves in response to different treatments (n = 3), Lowercase letters represent significant differences among different treatments (P < 0.05); C. GUS tissue staining of tobacco leaves after different treatments, Nb. Nicotiana benthamianaFig.6 GUS activity of apple MdMAX1 promoter in tobacco leaves after different treatments
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
