根域限制對葡萄根中IAA含量及其代謝相關基因表達的影響
2021-03-10 08:21:52糾松濤王繼源許文平張才喜趙麗萍王世平
西北植物學報 2021年1期
徐 巖,糾松濤,王繼源,王 磊,許文平,張才喜,趙麗萍,王世平
(上海交通大學 農業與生物學院,上海 200240)
根系作為植物重要的營養器官,負責從土壤中吸收水分和礦質元素,并合成氨基酸、激素等物質[1]。20世紀90年代初期,研究人員從盆栽實踐中獲得啟發,開始探索限制植物根系的栽培方式,即根域限制栽培(root restriction cultivation)。根域限制栽培是利用物理或生態方式將植物的根系控制在一定的容積內,通過控制根系生長來調節地上部的營養生長和生殖生長平衡的一種非傳統栽培方式[2]。先前研究表明,根域限制栽培下葡萄的根系構型發生顯著變化[3],葡萄果實品質也得到顯著提升[4-5]。
內源激素作為重要的植物生長調節物質,在植物的生長發育過程中發揮著重要的調節作用,尤其是對根系發生的影響。其中,吲哚-3-乙酸(IAA)是生長素(auxin)的主要活性成分,參與植物根系發育的諸多方面,其中包括調控主根生長、根毛的發育、側根原基的形成以及不定根的生長[6-8]。植物根系的生長受到根系組織中IAA濃度的影響,適當的IAA濃度有助于根系的生長,但過高或過低的濃度均不利于根系的生長[9]。生長素在地上部生命活動旺盛的部位合成,如莖尖、幼葉、胚芽鞘等器官,合成的生長素會暫時儲存在“庫”器官或者直接通過極性運輸到根尖等作用部位[10],該過程涉及生長素的合成、極性運輸等過程。
植物體內的生長素主要以色氨酸為前體,在氨基轉移酶(tryptophan aminotransferase, TAA)的作用下生成吲哚-3-丙酮酸(indole-3-pyruvate, IPA),IPA又在黃素單加氧酶(flavin monooxygenases, YUC)的作用下形成IAA[11-12]。生長素的極性運輸主要由向胞內運輸的AUXIN蛋白和向胞外運輸的PIN蛋白來實現[13-14]。在植物根中有2種不同的極性運輸方式,即向基性和向頂性運輸,主要通過AUX/LAX、PIN和ABCB/MDR/PGP等運輸載體來完成[15]。通過AUX和PIN等運輸載體的協同作用,IAA在根中形成濃度梯度,這是IAA對植物根系發育調控的關鍵。生長素的信號傳遞需要與受體結合來完成,目前發現的生長素受體有生長素結合蛋白1(auxin binding protein 1, ABP1)和運輸抑制劑響應蛋白1(transport inhibitor response 1, TIR1)[16-17]。此外,根系的生長不僅需要根尖分生區細胞不斷分裂增殖,還需要根伸長區細胞的伸長。CycA1、cycD3、cdc2、cycB1均屬于細胞周期調控蛋白,在有絲分裂過程中發揮著關鍵作用[18-19]。LRP1(lateral root primordia 1)在不定根及側根原基中特異表達,是一個調控根系伸長的轉錄因子,具有較強的組織特異性。SWP1(struwwelpeter 1)能夠結合到LRP1基因啟動子序列,對其表達起抑制作用。
近年來,根域限制栽培技術在葡萄生產過程中得到了廣泛應用,關于根域限制栽培影響葡萄根系發育和果實品質的研究也日漸深入[20-21],但是根域限制是如何調控葡萄根中生長素代謝、根發育相關基因的表達水平來影響內源IAA的合成和分配,進而影響其根系構型的還不清晰。因此,本研究以一年生‘玫瑰香’葡萄為試驗材料,對根域限制和傳統栽培下的葡萄根系中生長素的含量、生物合成、轉運以及信號轉導相關基因(VvTAA1、VvTAR2、VvYUC2、VvYUC4、VvYUC6、VvYUC8、VvABP1、VvTIR1、VvAUX1、VvLAX1、VvLAX2、VvLAX3、VvPIN1、VvPIN4、VvABCB1和VvABCB19)、細胞周期相關基因(VvcycA1、VvcycB1、VvcycD3和Vvcdc2)和根系伸長相關基因(VvLRP1和VvSWP1)表達豐度進行檢測,以期闡明根域限制栽培影響葡萄根系構型變化的內在機制,從而進一步完善根域限制栽培理論。
1 材料和方法
1.1 材料及處理
本研究以300株一年生‘玫瑰香’葡萄扦插苗為試驗材料,定植于上海交通大學果樹研究室玻璃溫室(31°11′N,121°29′E)內。其中,150株采取根域限制栽培(簡稱限根組),定植在直徑30 cm、高40 cm的限根器中,限根器的四周分布著透水孔,下部用塑料托盤與地面隔開;將土壤、有機肥、河沙按照1∶1∶1混合作為栽培基質。其余150株采取傳統地栽方式(非根域限制栽培,簡稱對照組),將溫室中的土壤深翻20 cm后,在其上堆積相同成分的栽培基質,形成厚度為40 cm、寬300 cm的栽培床。對照組初始栽培株行距為60 cm ×60 cm,為保證不同植株間根系發育有足夠的空間,我們采取間伐的策略進行樣品采集。2種栽培模式下地上部管理方式一致,均保持單蔓生長,每周對副梢進行一次修剪,所有試驗材料均配置統一的滴灌設備。肥水管理參照婁玉穗等[5]的方法進行。
1.2 樣品采集
實驗材料定植于2019年3月29日,經過10 d的緩苗,分別于定植后10、25、40、55、70、85、100、115、130、145、160和205 d采集限根組和對照組植株的根系樣品,限根組和對照組分別記為RR和nR。由于定植后10和25 d新生白根數量少,分別選取18株樹進行取樣,6株為一個重復,共設置3組重復;隨后白根數量增加,每個取樣點分別選取6株樹,2株為一個重復,設置3組重復。為保證根系完整性,采用流水沖洗根部周圍土壤,挖取植株時,以葡萄的主干為中心,以冠幅為參考標準確定根部挖取范圍;限根組葡萄根系采樣時,用流水將限根器內的土壤淋濕,后將限根器拆開,同樣用流水將土壤沖散,取出葡萄全根系。清洗干凈取出的葡萄全根系,用吸水紙吸干,將葡萄根系均勻攤開、置于黑色背景布上拍照。同時取下幼嫩的白根,迅速用液氮處理,并保存于-80 ℃超低溫冰箱中備用。
1.3 生長素含量測定
葡萄根中生長素(IAA)含量利用固相萃取反相高效液相色譜熒光檢測法,參照Xin等[22]和符繼紅等[23]的方法。色譜柱采用Waters sunfire C18(150 mm×4.6 mm,5 μm),流動相為乙腈-10 mmol/L乙酸鈉(20∶80,V/V,乙酸調節至pH 3.5)。在流速1 mL/min,進樣量20 μL,柱溫20 ℃,樣品溫度10 ℃,熒光激發和發射波長分別為275和345 nm條件下,分離測定IAA含量。
1.4 根系細胞結構觀察
選取限根組和對照組中定植70 d的葡萄根系進行細胞結構觀察。觀察的根系樣品包括老根從距離根莖連接處2 cm處橫剖、幼根從距離根尖2 cm處橫剖、幼根根尖縱剖和毛細根橫剖。石蠟切片的制作過程參照安吉翠[24]的方法。制片完成后使用Olympus BX61顯微鏡進行觀察攝影。
1.5 基因表達分析
1.5.1 總RNA提取總RNA的提取使用Tiangen公司的RNA plant plus Reagent植物總RNA提取試劑盒,按照說明書步驟操作。
1.5.2 引物設計利用Primer 5軟件進行特異性引物設計,要求上游引物和下游引物的Tm值相差在1 ℃以內,GC含量保持在40%~60%;引物長度保持在17~25個堿基對(base pair,bp);引物合成后,稀釋成100 μmol/L的母液,-20 ℃保存備用,用時可稀釋成10 μmol/L的工作液使用。引物序列見表1,委托上海睿勉生物進行引物合成。
1.5.3 cDNA的合成和熒光定量PCR以總RNA為模板,參照TaKaRa的PrimeScriptTMRT reagent Kit with gDNA Eraser(Perfect Real Time)試劑盒說明書反轉錄合成cDNA。以cDNA為模板,使用1.5.2中合成的引物序列,參考TB GreenTMPremix Ex TaqTMⅡ試劑盒的使用步驟,利用Bio-Rad iQ5(Hercules, CA, USA)熒光定量PCR儀檢測目的基因的表達水平。優化后的反應條件為:95 ℃預變性60 s;95 ℃變性 5 s,60 ℃退火 15 s,40個循環;72 ℃ 20 s。試驗設置3次重復。參考Jiu等[25-26]的方法,以葡萄KyActin1基因作為內參基因,各基因的定量水平采用2-ΔΔCt方法計算。
1.6 數據處理
用Excel 2010和SAS 9.2軟件(SAS Institute, Cary, North Carolina)對獲取的數據進行統計分析,并進行單因素方差分析(One-way ANOVA)及最小顯著性差異法(LSD法)檢驗(P<0.01),使用Origin 9軟件制圖。

表1 用于熒光定量PCR 的 引物序列
2 結果與分析
2.1 根域限制對葡萄根系構型和細胞結構的影響
首先,與對照組相比,限根組葡萄根系構型發生顯著變化,主要表現在根域限制后根尖處出現大量集群根、不定根持續發生、根系出現螺旋卷曲(圖1,A)。其次,根域限制栽培后,由于葡萄的根系構型發生了極顯著的改變,與之密切相關的根系細胞結構和細胞大小也會相應發生改變(圖1,B)。其中,葡萄老根的細胞在根域限制處理后大部分呈圓形或橢圓,相比較而言,對照組老根的細胞大部分呈長方形,細胞排列更緊密(圖1,B、a);葡萄具有吸收能力的毛細根結構在根域限制后也出現了顯著的變化,具體表現在表皮細胞分布更均勻,中柱鞘和皮層細胞更大,內皮層細胞更厚(圖1,B、b);葡萄幼根根尖的根冠區對根尖主要起到保護作用,根域限制處理后,其根冠區明顯更厚(圖1,B、c);而且根域限制栽培下幼根細胞排列更疏松,細胞更長、更大(圖1,B、d)。
2.2 根域限制對葡萄根中IAA含量的影響
為進一步解析根域限制影響葡萄根系構型的機理,本研究采用液質聯用法(HPLC-MS/MS)分析了根域限制對一年生‘玫瑰香’葡萄幼嫩根系中IAA含量的影響(圖2)。其中,在定植后40、100、145和205 d,限根組葡萄幼根中IAA含量極顯著低于對照組(P<0.01),尤其在定植后205 d時,對照組根系中IAA含量高達255.2 ng/g,約是對照組的19.0倍。整體來看,限根組葡萄根系中IAA含量在4個采樣時期呈現先降低后上升的趨勢,而在對照組中IAA的含量呈現先升高后降低然后又快速上升的趨勢。由此可見,根域限制處理不僅對葡萄根中IAA的含量產生影響,還使得IAA含量在不同時期發生顯著動態變化。

不同大寫字母表示相同時期nR和RR之間在0.01 水平差異顯著(P<0.01);下同圖2 根域限制條件下“玫瑰香”葡萄的根系IAA含量的變化Different capital letters indicate significant differences between nR and RR on the same growth days at the 0.01 level (P<0.01); the same for the following figuresFig.2 The root IAA content of grapevine (Vitis vinifera L. cv. Muscat Hamburg) under root restriction treatment
2.3 根域限制對葡萄細胞周期和根伸長相關基因表達的影響
通過形態學觀察和解剖結構分析發現,根域限制抑制葡萄不定根的伸長、致使不定根的根尖簇生大量集群根,根尖膨大且伴有明顯的增生。因此,本研究對幾個與細胞周期調控和根伸長相關的基因進行了qRT-PCR分析。結果(圖3)顯示,在大多數取樣時期,根域限制處理對葡萄細胞周期調控和根伸長相關基因的表達水平產生了顯著的影響。首先,根域限制對細胞周期調控基因VvcycA1、VvcycB1、Vvcdc2、Vvcycd3表達量的影響存在相似性,大部分時間都顯著高于同期對照(圖3,Ⅰ~Ⅳ)。其中,限根組VvcycA1基因表達量除在定植后10 d顯著低于對照,在定植40和55 d與對照相近外,其余時間與對照組相比均顯著上調表達;而根域限制顯著誘導VvcycB1基因在定植后70、85、100、130和205 d時比對照組上調表達,在定植10、140和160 d時下調表達;限根組Vvcdc2基因的表達量除定植后25和55 d顯著低于對照組外,其余10個采樣時期均顯著高于對照組;同樣,限根組Vvcycd3的表達量在定植后160 d比對照組顯著下調,在定植10、25和145 d與對照組相近,其余取樣時期均顯著高于對照組。由此可知,根域限制可能通過誘導VvcycA1、VvcycB1、Vvcycd3和Vvcdc2基因的上調表達來加快細胞分裂的進程。其次,根域限制也顯著影響根伸長相關的基因VvLRP1和VvSWP1的表達(圖3,Ⅴ、Ⅵ)。其中,與對照組相比,根域限制在定植后的7個采樣時期均顯著抑制VvLRP1的表達水平,而在定植后的9個采樣時期顯著促進VvSWP1的表達水平,說明這兩個基因在根域限制誘導下發揮相互拮抗的作用。然而,根域限制處理后VvLRP1基因上調表達的5個時期,VvSWP1基因同樣上調表達,又說明在有些時期二者發揮著協同的作用。由此可以看出,根域限制處理后,VvLRP1和VvSWP1在根系不同發育階段發揮的作用存在較大差異。
2.4 根域限制對葡萄生長素代謝相關基因表達的影響
2.4.1 生長素合成相關基因表達前人研究表明,IPA途徑對擬南芥中的生長素生物合成至關重要,并且YUC家族是該途徑中的限速步驟[27]。本研究結果顯示,葡萄根中IAA合成相關基因VvTAA1、VvTAR2、VvYUC2、VvYUC4、VvYUC6和VvYUC8的表達在根域限制條件下受到了不同程度的影響(圖4)。其中,根域限制葡萄根系VvTAA1基因在定植10~40 d下調表達,從定植55 d開始受到顯著誘導并上調表達,其在定植后115 d時表現尤為明顯,限根組表達量約是對照的26.2倍;隨后除定植160 d顯著下調表達外,其余時期仍表現為上調表達(圖4,Ⅰ)。與VvTAA1基因表現相反,VvTAR2基因表達量在采樣前期受根域限制的上調誘導更為明顯,尤其在定植55 d時表現最為突出,限根組表達量約是對照的31.2倍;之后,短暫顯著下調表達(定植70 d),再持續上調表達(定植85~130 d);隨之,在定植145~205 d顯著下調表達(圖4,Ⅱ)。同時,與對照組相比,葡萄根系VvYUC2和VvYUC4基因的表達水平在根域限制誘導下多數時期均上調,而VvYUC6和VvYUC8基因多數時期表達量均下調(圖4,Ⅲ~Ⅵ)。整體來看,在定植后100、145和205 d時,葡萄根系中VvYUC6的表達量與IAA含量變化一致,而在定植后40、145和205 d時,根中VvYUC8的表達量與IAA含量變化一致,說明這兩個基因可能在葡萄根域限制調控根系IAA合成中發揮著重要的作用。

圖3 根域限制條件下葡萄幼根中細胞周期和根伸長基因表達的變化Fig.3 Changes in expression levels of cell cycle and root growth regulatory genes of grapevine (Vitis vinifera L. cv. Muscat Hamburg) roots under root restriction treatment
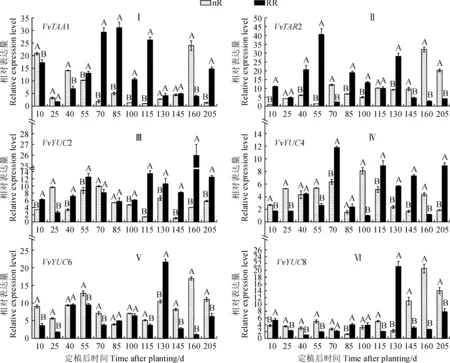
圖4 根域限制條件下葡萄根系IAA合成相關基因表達的變化Fig.4 Changes in expression levels of IAA biosynthesis genes of grapevine (Vitis vinifera L. cv. Muscat Hamburg) under root restriction treatment

圖5 根域限制條件下葡萄根系IAA受體基因表達的變化Fig.5 Changes in expression levels of IAA receptor genes of grapevine (Vitis vinifera L. cv. Muscat Hamburg) roots under root restriction treatment
2.4.2 生長素受體基因表達生長素與受體結合后才能實現信號的順利傳遞,從而發揮調控作用[16]。目前已經明確的IAA受體蛋白有ABP1和TIR1[17,28-29]。qPCR分析結果也顯示,根域限制下葡萄根系生長素受體基因VvABP1和VvTIR1的表達量均發生了顯著變化(圖5)。其中,根域限制處理根系VvABP1基因的表達水平在定植后25、85、100和205 d時均顯著高于對照組,而在定植后10、55、70、115、130和160 d時均顯著低于相應對照組,尤其在定植后160 d時約是對照組的1/14(圖5,Ⅰ);同時,根域限制處理在定植后70、100、130和205 d時均顯著誘導根系中VvTIR1基因的上調表達,特別在定植后130 d時表達豐度最高,根域限制的誘導效果最為顯著;而對照組根系中VvTIR1基因在定植后160 d時表達豐度最高(圖5,Ⅱ)。整體來看,葡萄根系VvABP1和VvTIR1基因在整個采樣時期的表達變化趨勢較為一致。

圖6 根域限制條件下葡萄根系中IAA運輸相關基因表達的變化Fig.6 Changes in expression levels of IAA transport-related genes of grapevine (Vitis vinifera L. cv. Muscat Hamburg) roots under root restriction treatment
2.4.3 生長素運輸相關基因表達生長素主要依賴細胞膜上的載體蛋白實現向細胞內外的運輸,AUX/LAX家族是IAA向細胞內運輸的載體蛋白[30],而ABCB/PGP/MDR家族和PIN蛋白家族是IAA向細胞外運輸的載體[31]。圖6顯示,葡萄根系VvAUX1等8個生長素運輸相關基因在根域限制后的表達模式存在一定的相似性,它們在定植后70、85和130 d時均受到顯著誘導而上調表達,這與上述根域限制下根系大量發生的時期相吻合,而它們在定植后145和160 d時的表達豐度與其他時期相比處于較低的水平。其中,在定植后70 d,根域限制誘導VvAUX1(圖6,Ⅰ)和VvLAX2(圖6,Ⅲ)上調表達效果最為顯著;根域限制誘導VvLAX1(圖6,Ⅱ)和VvLAX3(圖6,Ⅳ)的表達量均在定植后130 d達到峰值,分別為同期對照組的2.7和5.1倍;根系VvPIN1和VvPIN4的表達模式較為相似,它們在限根組定植后55~130 d以及205 d時的表達水平均顯著高于對照組,并以定植后85 d和130 d時受到根域限制的誘導作用最為顯著,且兩基因均在定植后85 d時增長幅度最大,此時限根組分別為對照組的15.1和23.7倍(圖6,Ⅴ、Ⅵ);在定植后70 d、85 d、100 d、130 d和205 d,根域限制組中VvABCB1的表達量顯著高于對照組,而在其他時期均低于對照組(圖6,Ⅶ);在定植后25、55、85、100、115、130和205 d時,根域限制組中VvABCB19的表達量均顯著高于對照組,而在其他時期與對照組相比沒有顯著性差異(圖6,Ⅷ)。整體來看,在多數時期,根域限制處理均能顯著誘導生長素運輸相關基因的表達水平,也表明根域限制加快了葡萄根中生長素的運輸過程。
3 討 論
3.1 根域限制對葡萄根系細胞結構、細胞周期和根系伸長的影響
與傳統地栽組相比,葡萄根系構型在根域限制后發生極顯著變化,主要表現在根尖處出現大量集群根、不定根持續發生、根系出現螺旋卷曲,其表型改變與根系細胞結構和細胞大小的改變密切相關。根域限制處理后,根系細胞排列更加疏松,說明細胞需要不斷脫落更新,這與根域限制后根系更新加快的表型吻合。其次,可能由于限根器對根尖存在壓力脅迫,導致幼根根尖的根冠區更厚,從而實現對根尖的保護。另外,由于根系被局限在一定的容積內,導致吸收根更多、更粗壯,吸收根的內皮層細胞則更厚,中柱鞘和皮層細胞都更大。
根系的伸長得益于根尖分生區細胞的分裂和伸長區細胞的延伸。本研究篩選了細胞周期調控相關基因VvcycA1、VvcycB1、VvcycD3和Vvcdc2以及根系伸長相關的基因VvLRP1和VvSWP1進行表達分析。在8月中旬到11月初,葡萄細胞周期調控相關基因VvcycA1、VvcycB1、VvcycD3和Vvcdc2的表達量受到根域限制誘導呈現顯著上調表達,這也與該時期根系快速更新的變化相一致。然而,在4月初到5月中下旬,根域限制組和對照組中上述基因的表達量幾乎沒有顯著差異,可能是由于在根域限制處理的前期,限根器對根系伸長生長的阻礙作用尚不明顯,使得根系中細胞周期相關基因的表達未發生顯著變化。LRP1基因在不定根及側根原基中特異表達,對根系伸長和側根發生起到了促進作用[32]。SWP1基因位于LRP1基因的上游,會抑制LRP1基因發揮作用。前人研究表明,敲除SWP1基因使得LRP1的表達量上調,進而促進擬南芥根系的伸長生長[32-33]。本研究結果表明,在定植后10、70、85、100和205 d,葡萄根域限制顯著誘導VvLRP1基因上調表達,潛在表明上述時期側根發生和根系伸長的速度快于對照組。然而,本項研究也發現,根域限制處理后,VvLRP1和VvSWP1在葡萄根系不同發育階段發揮的作用存在較大差異,二者在部分時期協同調控根系生長,而在有些時期VvSWP1的上調表達會抑制VvLRP發揮作用,這與Krichevsky等[32]的研究結果有差異。
3.2 根域限制對葡萄根系生長素代謝相關基因表達的影響
植物根中的IAA主要依賴于根尖自身合成和地上部幼嫩組織合成后轉運而來。因此,要明確根域限制對根中IAA含量的影響,不僅要分析根域限制處理對IAA合成基因的影響,還要分析根域限制對IAA運輸相關基因的影響。本研究結果表明,在定植后40、100、145和205 d時,根域限制組葡萄根中IAA的含量均顯著低于對照組。同時,6個IAA合成相關基因中僅有VvYUC6和VvYUC8的表達模式與IAA含量變化趨勢較為一致,表明這2個基因可能對于根域限制下葡萄根中IAA合成發揮著重要的作用;而其他幾個基因,諸如VvYUC2、VvTAA1等,在多數時期卻受到根域限制處理的誘導上調表達,這與根域限制下根中IAA含量下降的趨勢不一致。這可能從側面反映出,根域限制可能對葡萄根中IAA的極性運輸產生了更大的影響。植物根中生長素的極性運輸包括向基性和向頂性兩個不同方向,其中多種生長素轉運蛋白參與該過程[15]。本研究發現,在葡萄定植后的多個時間點,根系向基性轉運蛋白基因VvAUX1、VvPIN1和VvPIN4的表達水平明顯上調,同時向頂性轉運蛋白基因VvLAX1/2/3、VvABCB1和VvABCB1的表達也受到根域限制的誘導呈現上調表達。表明葡萄根域限制致使生長素的極性運輸加劇,這可能是因為根域限制下根域空間有限,根系生長受限,幼嫩部位合成的生長素只有通過加快極性運輸進程才能使葡萄抵御這一根域逆境脅迫。因此,根域限制通過調控生長素合成和極性運輸相關的基因表達豐度來影響根系中生長素的合成和運輸。
綜上所述,根域限制下葡萄根系構型、細胞結構以及內源IAA均發生顯著變化。此外,根域限制可能通過改變了根系中生長素合成和極性運輸相關基因的表達豐度,從而影響根系中生長素的合成和運輸,進而改變了葡萄的根系構型以適應根域限制的栽培環境。該研究有助于闡明根域限制栽培影響葡萄根系構型變化的內在機制,為進一步完善根域限制栽培理論奠定了堅實的基礎。
