基于COⅠ基因片段的中國南部沿海近親擬相手蟹的群體遺傳多樣性研究
2021-03-15 09:00:42楊明柳徐巖高霆煒潘紅平吳斌閻冰
海洋學報 2021年2期
關鍵詞:紅樹林
楊明柳,徐巖,,高霆煒,潘紅平,吳斌,閻冰*
(1.廣西科學院 廣西紅樹林研究中心 廣西紅樹林保護與利用重點實驗室,廣西 北海 536007;2.廣西大學 動物科學技術學院,廣西 南寧 530005)
1 引言
近親擬相手蟹(Parasesarma affine)隸屬于方蟹總科(Grapsoidea),相手蟹科(Sesarmidae),擬相手蟹屬(Parasesarma),廣泛分布于東海、南海、印度?西太平洋[1],棲息于紅樹林濕地中,以紅樹植物凋落物、沉積物、藻類等為食,是一種活動能力和適應能力均很強的常見種與優勢種[2-3]。近親擬相手蟹是紅樹林蟹類中重要種類之一,在紅樹林生態系統的結構和功能中扮演著非常重要的角色[4]。目前,國內外有關于近親擬相手蟹的研究已涉及生物學、攝食行為、形態分類、線粒體DNA 全基因組等方面[5-8],但未見其群體遺傳相關的研究報道。近年來,我國紅樹林遭到了不同程度的破壞,天然紅樹林植被的退化和生境的破碎化對紅樹林蟹類群落造成了一定影響,環境污染、圍塘養殖、港口建設等人為活動也使得蟹類資源面臨著巨大威脅[2,9-10]。因此,開展近親擬相手蟹群體遺傳的研究,對全面了解近親擬相手蟹群體遺傳多樣性現狀及加強紅樹林蟹類資源保護工作具有重要意義。
線粒體DNA 具有分子結構穩定、母系遺傳、進化速度快、核苷酸替代速率高等特征,是研究動物分子系統發育和群體遺傳的重要遺傳標記[11]。細胞色素氧化酶Ⅰ(COⅠ)基因是線粒體DNA 中最常用的標記之一,可用于揭示種群之間遺傳結構,目前已被廣泛地應用于蟹類的群體遺傳研究中[12?15]。本研究采用線粒體COⅠ基因對我國南部沿海12 個近親擬相手蟹群體進行群體遺傳學研究,分析近親擬相手蟹群體的遺傳結構和歷史動態,以期為紅樹林蟹類的資源保護與利用提供基礎資料。
2 材料與方法
2.1 樣品采集
本研究所用的近親擬相手蟹于2012 年7 月至2013 年6 月采集于福建龍海、福建云霄、廣東珠海、廣東陽江、廣東雷州、廣東徐聞、廣西合浦山口、廣西北海、廣西防城港、海南洋浦、海南東方等12 個采樣地,共計222 個個體(表1,圖1)。所有個體置于95%酒精中固定,并保存于?20℃冰箱。
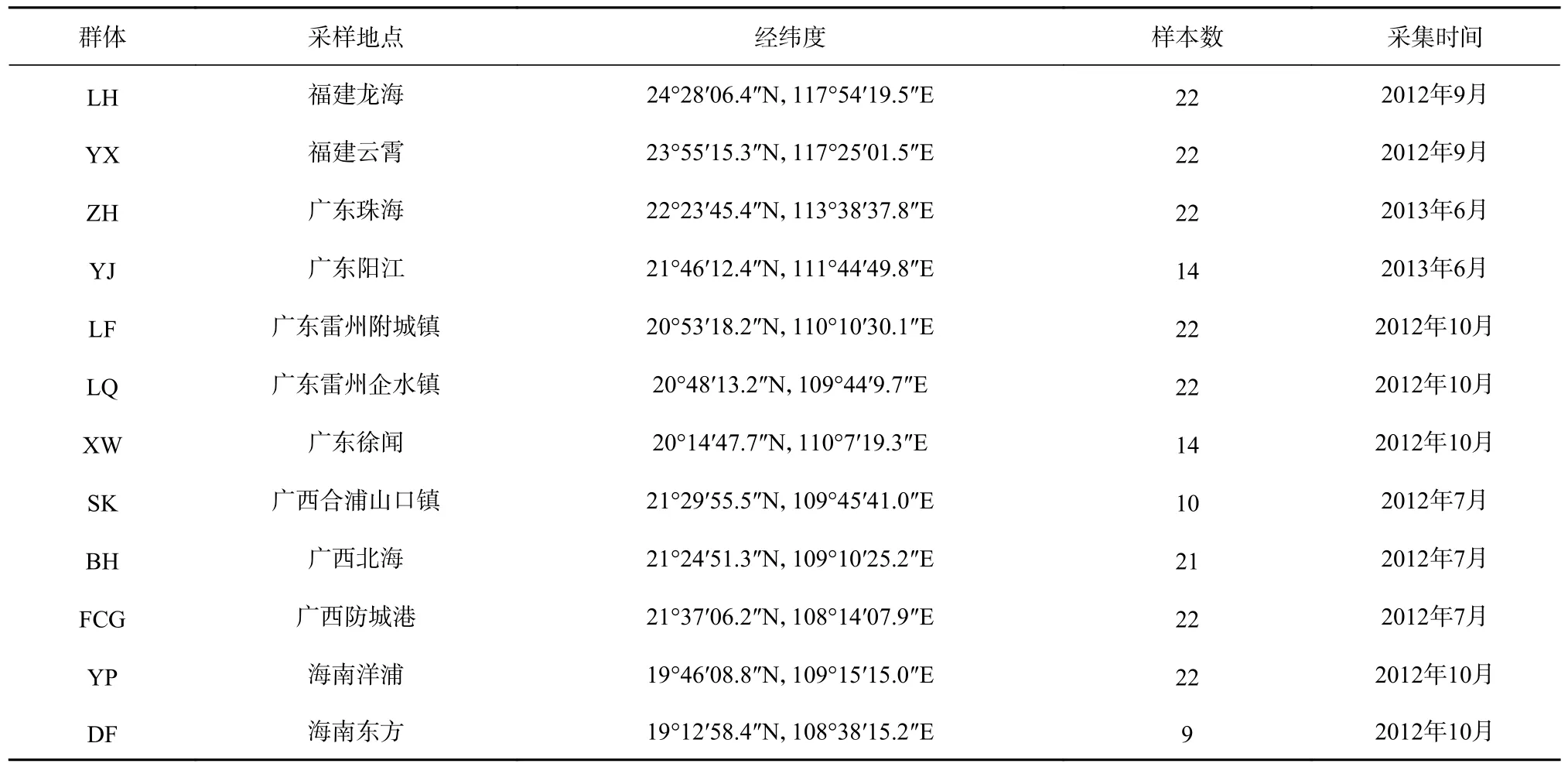
表1 中國南部沿海近親擬相手蟹的采樣信息Table 1 The sampling information of P.affine from the South China Sea
2.2 實驗方法
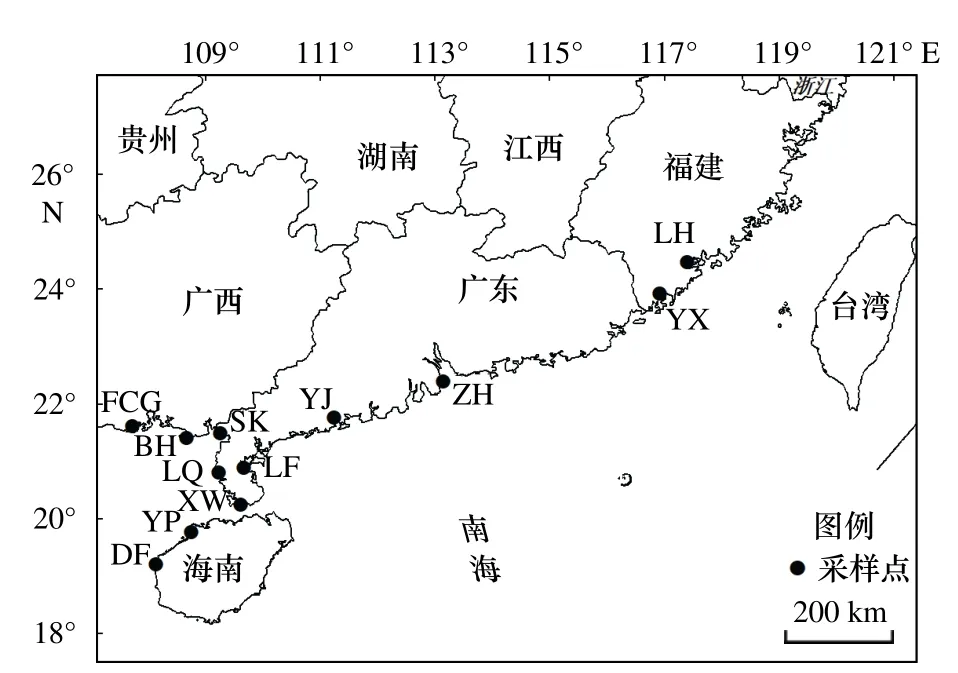
圖1 近親擬相手蟹的采樣點地理位置Fig.1 Locations of the sampling stations of P.affine
取近親擬相手蟹步足肌肉10 mg 左右,采用E.Z.N.A.TMSQ Tissue DNA Kit 試劑盒(Omega 公司) 提取總DNA,于?20℃分管保存備用。使用無脊椎動物通用引物COILl490:5’-GGTCAACAAATCATAAAGATATTGG-3’;COIH2198:5’-TAAACTTCAGGGTGACCAAAAAATCA-3’[16]擴增COⅠ基因片段,引物由廣州英濰捷基測序公司合成。PCR 反應體系為50 μL,其中包含2×Taq10 MasterMix (北京奧賽博公司)25 μL,模板 DNA 2 μL,引物各2 μL,其余用 Milli-Q-Water 補足到50 μL。PCR 反應條件為94℃預變性2 min,94℃變性45 s,47℃退火50 s,72℃延伸1 min,35 個循環,最后72℃再延伸8 min。PCR 產物經1%的瓊脂糖凝膠電泳檢測確認為目的片段后,將樣品送至廣州英濰捷基測測公司進行純化,并用ABI3730XL 測序儀進行正反鏈雙向測序。
2.3 數據處理
所獲得的序列均由DNAstar 7.1[17]軟件包中的SeqMan、MegAlign 進行校對、編輯、排序。Dnasp v5.0[18]統計單倍型數目、多態位點數、簡約信息位點數,Arlequin3.1[19]計算單倍型多樣性和核苷酸多樣性。采用MEGA7.0[20]軟件計算近親擬相手蟹群體內和群體間平均遺傳距離,并基于Kimura 2-parameter 模型構建單倍型鄰接樹(1 000 次自展重復)。使用Network 4.51[21]軟件包中介鄰接網絡法(Median Joining,MJ)構建單倍型中介網絡圖。利用Arlequin 3.1[19]軟件進行分子方差分析(Analysis of Molecular Variance,AMOVA),來評估近親擬相手蟹群體間的遺傳變異,計算群體間的遺傳分化指數Fst值,并用Tajima'sD和Fu'sFs中性檢驗及核苷酸不配對分布,來進行群體的歷史動態評估[22?23]。采用公式τ=2ut估算群體擴張發生的大致時間,其中t為群體發生擴張的時間,u為所研究的整個序列的進化速率,u采用2.3% 每百萬年的核苷酸分歧速率[24?25]。
3 結果
3.1 序列分析
本研究的近親擬相手蟹12 個群體共222 條序列經比對分析后,獲得長度為612 bp 的COⅠ基因片段,其A、T、C、G 的平均含量分別為28.93%、37.25%、17.17%、16.65%,A+T 含量(66.18%)明顯大于C+G 含量(33.82%),堿基存在明顯的偏向性(表2)。共檢測到34 個變異位點,包括19 個單核苷酸變異位點,15 個簡約信息位點,無插入或缺失位點。34 個變異位點共發生了35 次核苷酸替換,其中有30 次轉換,5 次顛換,轉換和顛換之比為6.0。
3.2 序列多態性分析
近親擬相手蟹總群體的單倍型多樣性指數Hd為0.508 9,核苷酸多樣性指數Pi為0.001 126。各群體間的單倍型多樣性指數Hd在0.222 2~0.710 0 之間,核苷酸多樣性指數Pi在0.000 362~0.001 726 之間,其中雷州企水群體的單倍型多樣性和核苷酸多樣性最高,海南東方群體的單倍型多樣性和核苷酸多樣性相對較低。
在222 個個體中,COⅠ基因片段檢測到的34 個變異位點共定義了40 個單倍型,由表3 可知,有29 個獨有單倍型和11 個共享單倍型。其中單倍型Hap2在12 個群體中均有分布,在群體中所占比例高達69.82%,為主體單倍型。Hap3 為10 個群體共享,在群體中所占 比例為6.31%;Hap22 為4 個群體共享,Hap30 為3 個群體共享,Hap7、Hap12、Hap18、Hap20、Hap29、Hap34 各為2 個群體共享。在12 個群體中,雷州企水群體的單倍型數量最多(10 個),其次為廣西北海群體(9 個),海南東方群體的單倍型最少(2 個)。由于部分群體的樣本個體數量較少,這可能會在一定程度上影響單倍型數量、單倍型多樣性和核苷酸多樣性的統計結果。
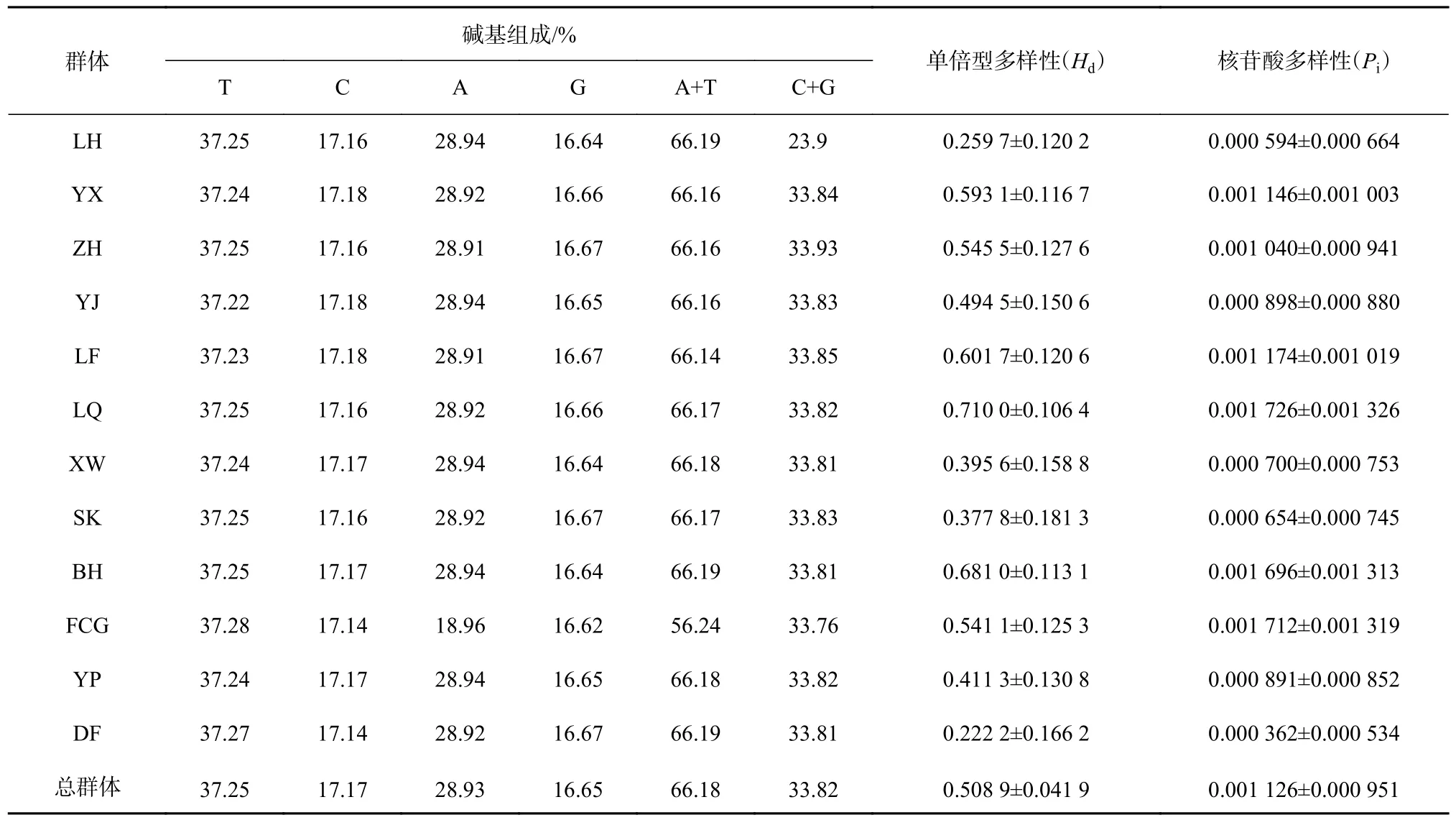
表2 近親擬相手蟹不同群體COⅠ基因片段的堿基組成和遺傳多樣性Table 2 The base composition and genetic diversity parameters of COⅠ gene fragments among different population of P.affine

表3 近親擬相手蟹12 個群體單倍型分布情況Table 3 Haplotype distribution of P.affine in twelve populations
3.3 系統發育分析與單倍型中介網絡圖
為了更好地了解近親擬相手蟹各單倍型之間的關系,以弧邊招潮(Uca arcua,基因登陸號:LC053370)為外群,構建了單倍型鄰接發育樹(圖2)。從系統發育樹可以看出,40 個單倍型沒有分成明顯的支系,不同地理群體的單倍型并沒有出現明顯的地域分群。利用中介網絡鄰接法構建的單倍型網絡中介圖呈星狀散射分布,主體單倍型為Hap2,其他單倍型與主體單倍型Hap2 僅保留一步或兩步突變。中介網絡圖中近親擬相手蟹各群體的單倍型呈零散分布,沒有明顯的地理系譜結構,其結果進一步支持了系統發育樹的分析(圖3)。

圖2 基于近親擬相手蟹COⅠ基因構建的40 個單倍型鄰接系統發育樹Fig.2 Neighbor-joining tree for 40 haplotypes of P.affine based on COⅠ gene
3.4 遺傳距離分析
從表4 可以看出,12 個近親擬相手蟹群體內的遺傳距離在0.000 36~0.001 73 之間,海南東方群體內的遺傳距離最小,為0.000 36;雷州企水群體內的遺傳距離最大,為0.001 73。在不同地理群體間的遺傳距離范圍為0.000 48~0.001 72,其中雷州企水群體與廣西北海群體遺傳距離最大,為0.001 72,而海南東方群體與福建龍海群體之間的遺傳距離最小,為0.000 48。這表明群體內和群體間的遺傳分化程度均很低,各群體間沒有形成地理隔離現象。遺傳分化系數Fst結果顯示,12 個群體間的Fst值均較小,范圍為?0.055 04~0.021 51,且統計學差異不顯著(p>0.05),表明各群體間沒有發生明顯的遺傳分化。絕大多數群體間的Fst值為負值,表明大部分群體內的遺傳差異水平大于群體間的遺傳差異水平。
3.5 分子方差分析
根據群體間遺傳距離及遺傳分化情況,將12 個群體作為一個整體進行分子方差分析,結果顯示,群體間的變異百分比為?0.4%,而組群內的變異百分比為100.4%,這亦表明了群體間的遺傳分化程度低,且群體的遺傳變異主要來自群體內,而不是群體間。總的固定指數(FST)為?0.004 02,表明來自同一個群體的兩個隨機個體比來自不同群體的兩個隨機個體有更大的遺傳差異(表5)。
3.6 群體動態分析
由于近親擬相手蟹群體間沒有顯著的遺傳分化,因此將12 個群體合并為一個大群體進行歷史動態分析。中性檢驗顯示(表6),總群體的Tajima’sD檢驗(Tajima’sD=?2.497 80,p<0.05)和Fu’sFs檢驗(Fu’sFs=?29.929 97,p<0.05)結果均為負值,差異顯著,并且Fu’sFs檢驗的絕對值大于Tajima’sD檢驗絕對值,提示近親擬相手蟹曾經發生過群體擴張事件。此外,核苷酸不配對分析結果表明,偏離方差(SSD)和Rg(Raggedness index)均較小,統計結果不顯著(p>0.05),表明未顯著偏離群體擴張模型,進一步暗示了近親擬相手蟹可能經歷過群體擴張。并且,近親擬相手蟹的單倍型核苷酸不配對分布圖呈明顯的單峰泊松分布,且觀測值不明顯偏離模擬值(圖4)。由此可見,中性檢驗和核苷酸不配對結果一致,均提示近親擬相手蟹群體在歷史上可能發生過群體擴張。基于核苷酸不配對分布得到的總群體擴張參數τ值為0.714 84,估算得出中國南部沿海近親擬相手蟹群體發生擴張時間大約在5.1 萬年前。
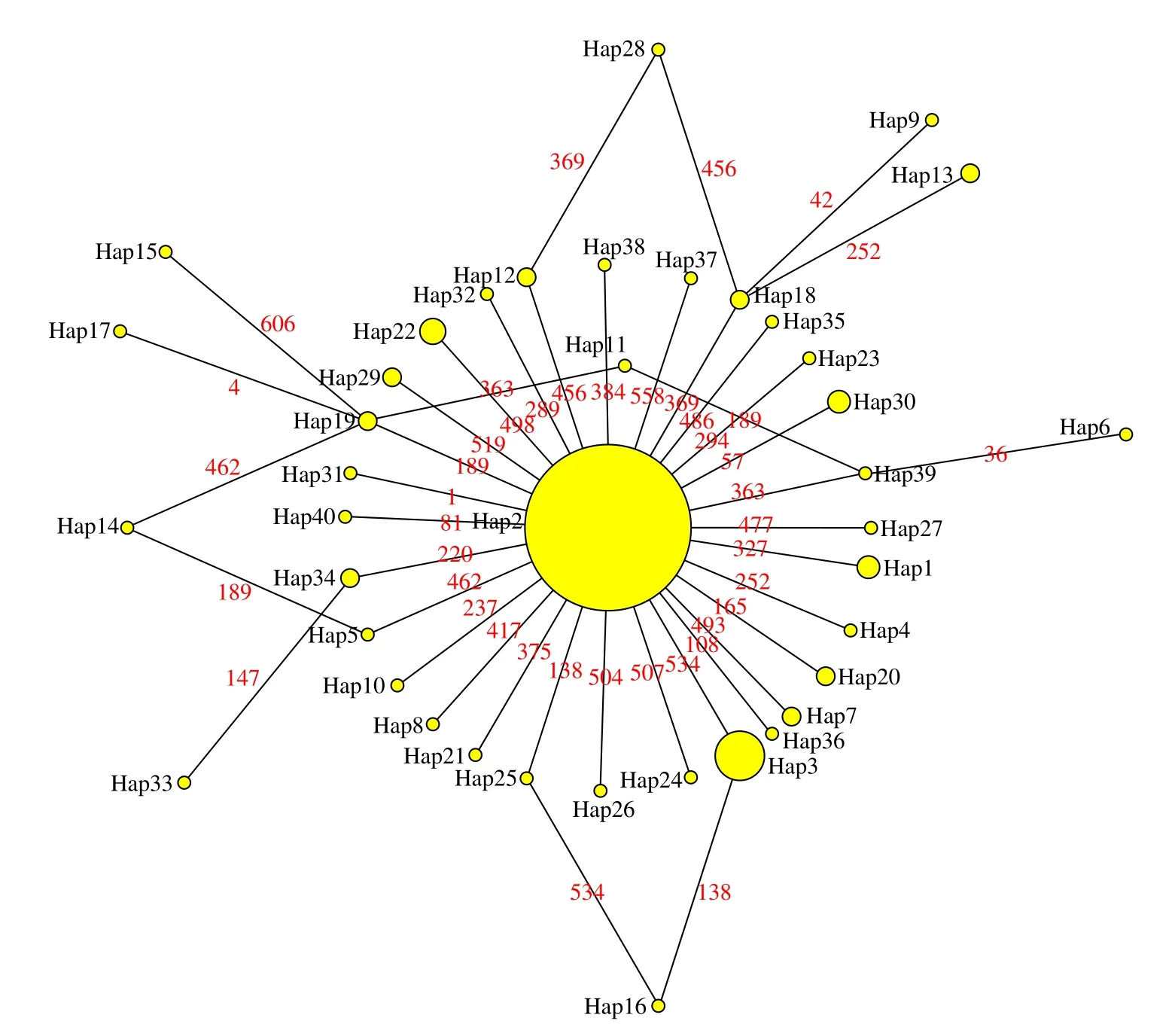
圖3 近親擬相手蟹12 個群體COⅠ基因單倍型中介網絡圖Fig.3 The median-joining network of COⅠ gene haplotypes in twelve populations of P.affine
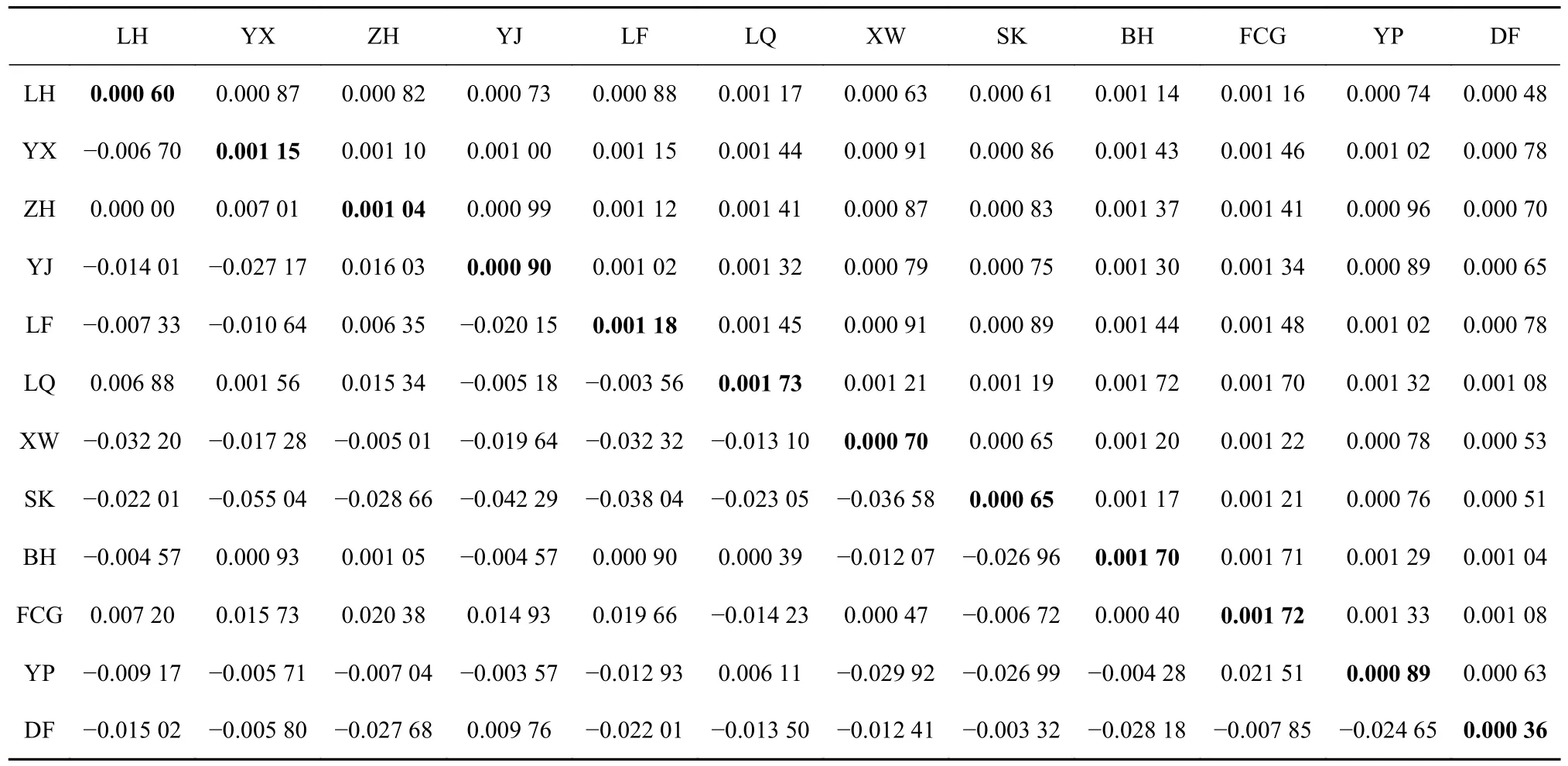
表4 基于COⅠ基因序列的近親擬相手蟹群體間遺傳分化系數Fst值(對角線下)和群體內(對角線)及群體間(對角線上)遺傳距離Table 4 F-statistics (below diagonal) within populations and genetic distance within (diagonal) and between(above diagonal) populations of P.affine based on COⅠ gene
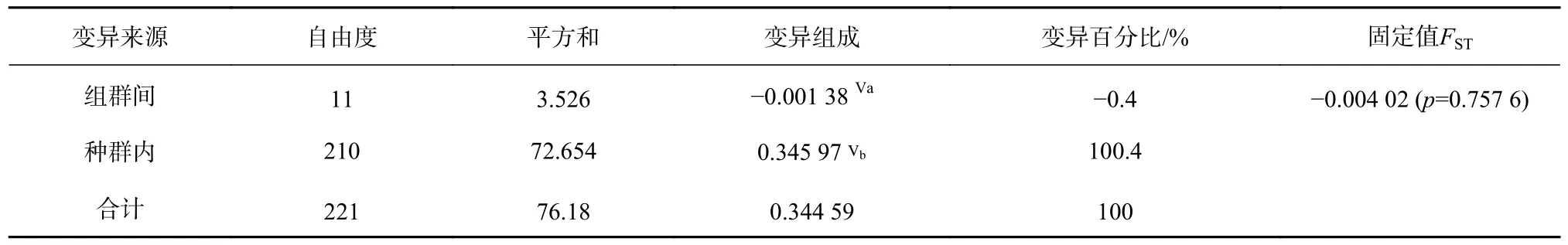
表5 近親擬相手蟹12 個群體的AMOVA 分析Table 5 Analysis of molecular variance (AMOVA) for the twelve P.affine populations
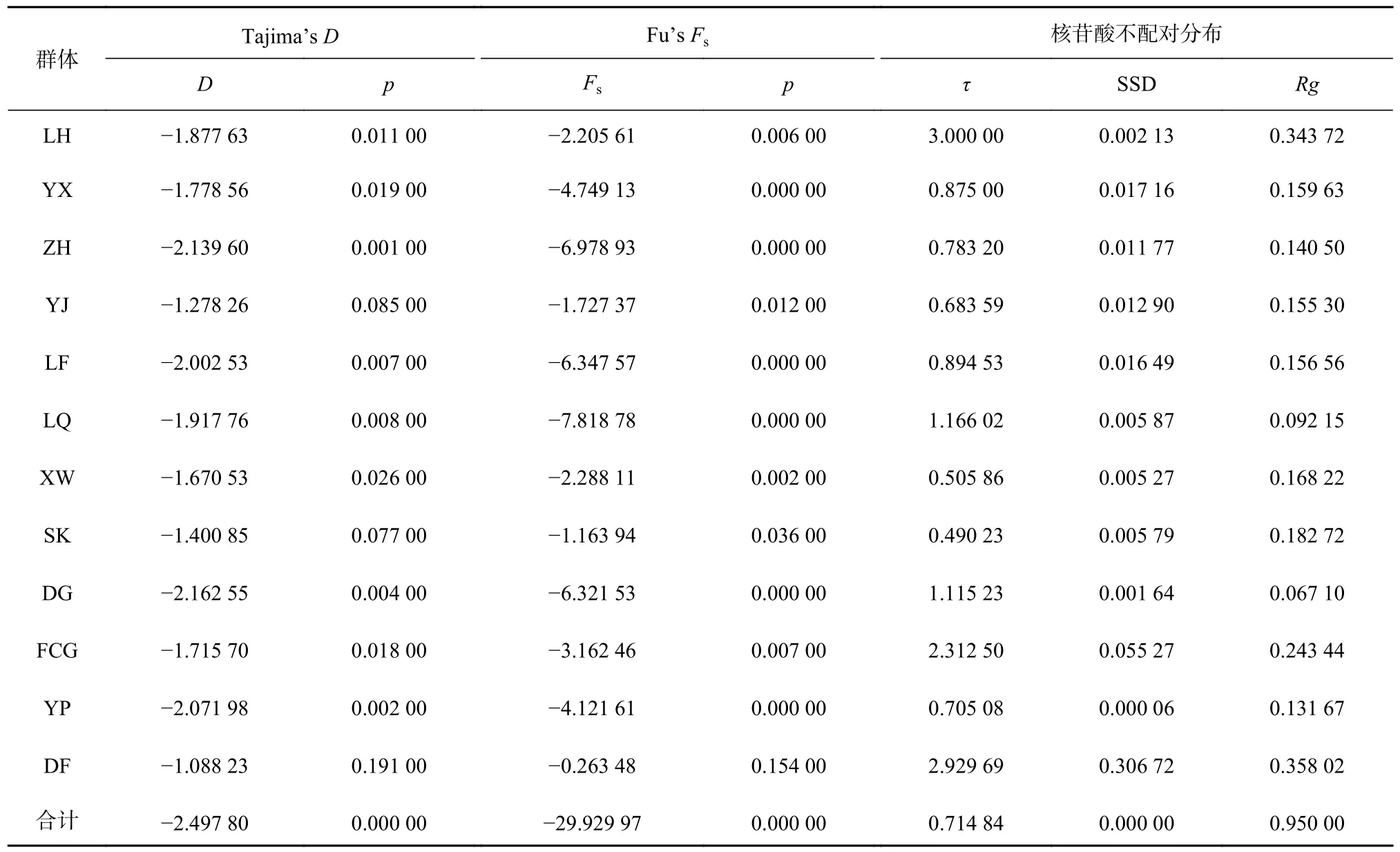
表6 近親擬相手蟹12 個群體的中性檢驗和核苷酸不配對分布Table 6 Neutral testes and mismatch distributions of nucleotide in twelve P.affine populations

圖4 近親擬相手蟹COⅠ基因單倍型的核苷酸不配對分布Fig.4 Mismatch distributions based on COⅠ gene haplotypes of P.affine
4 討論
4.1 遺傳多樣性分析
本研究中,我國南部沿海12 個近親擬相手蟹群體222 個個體的COⅠ堿基組成中A+T 含量占總堿基的66.18%,表現出較強的AT 偏向性,與已經報道的相手蟹科的雙齒近相手蟹(Perisesarma bidens)[14]、邁納新脹蟹(Neosarmatium meinerti)[26]、其他軟體動物(疣荔枝螺Reishia clavigera)[27]、星蟲動物(可口革囊星蟲Phascolosoma esculenta)[28]等的COⅠ堿基組成特點相似,符合無脊椎動物線粒體DNA 的堿基組成特征。
單倍型多樣性指數和核苷酸多樣性指數是衡量群體遺傳多樣性的一項重要指標[29]。根據單倍型多樣性指數和核苷酸多樣性指數的特征,Grant 和Bowen[30]對遺傳多樣性劃分為低Hd和低Pi(Hd<0.5,Pi<0.5%)、高Hd和低Pi、低Hd和高Pi、高Hd和高Pi等4 種遺傳特征類型。近親擬相手蟹12 個群體的單倍型多樣性指數為0.508 9,核苷酸多樣性指數為0.001 126,符合高Hd和低Pi類型,暗示其群體近期可能經歷了快速擴張事件[30]。這種高Hd和低Pi的遺傳多樣性特征不僅存在于蟹類等甲殼動物中[13,31],許多種海洋無脊椎動物,尤其是具有浮游幼體階段和高繁殖潛力的海洋無脊椎動物也存在這種特征[27?28,32]。此外,近親擬相手蟹群體較低的核苷酸多樣性可能是由于新形成的單倍型存在的時間較短,其累積的核苷酸突變很快就隨之消失而造成[33],瓶頸效應及隨后的群體擴張也可能會導致這種遺傳多樣性格局。
單倍型中介網絡圖中單倍型呈星狀散射分布,Hap2 位于網絡中介圖中心,其廣泛分布于各個群體,占總個體數的69.81%,這暗示著Hap2 為祖先單倍型。本研究檢測到40 種單倍型中有11 個共享單倍型和29 個獨有單倍型,獨有單倍型僅在1 個群體中的1 個個體表現,并且其他單倍型與主體單倍型Hap2僅保留一步或兩步突變,這種現象通常可以認為是群體瓶頸效應后,少量有效群體近期迅速擴張事件引起[34],進一步表明近親擬相手蟹可能經歷過群體快速爆發事件。
4.2 群體遺傳結構特征與群體歷史動態
遺傳分化系數Fst結果顯示,近親擬相手蟹12 個群體間的Fst值范圍為?0.055 04~0.021 51,統計學差異不顯著(p>0.05),群體間的遺傳分化水平很低。基于遺傳距離分析的結果發現,群體間的遺傳距離接近甚至小于群體內的遺傳距離,各群體間沒有形成地理隔離,AMOVA 分析結果也表明,群體間的遺傳分化程度低,群體的遺傳變異主要來自群體內。這些結果暗示著近親擬相手蟹各群體間存在廣泛的基因交流,并且不同地理群體可以以隨機交配的方式進行基因交流[29],推測中國南部沿海可能缺少能明顯影響近親擬相手蟹群體擴散的地理屏障。
許多具有浮游階段且有較強潛在擴散能力的海洋生物,在非常廣闊的范圍內通常會表現出很低的遺傳分化[35]。在大多數海洋無脊椎動物中,浮游性幼蟲發育是它們繁殖策略中很重要的一部分,浮游幼蟲期持續的時間越長,它們可能在洋流的作用下擴散得越遠,不同地理群體在較大的地理范圍內潛在的基因流就越高,從而導致遺傳同質性[36]。近親擬相手蟹是一種廣泛分布于紅樹林中的常見種類,與大多數紅樹林蟹類一樣,會與潮汐同步釋放幼蟲,這些浮游幼蟲可能會借助潮汐和洋流運輸作用跨越數十千米至數百千米進行擴散[37?38]。相手蟹的幼蟲發育通常經過4~5個溞狀幼體期和1 個大眼幼體期,幼蟲期持續將近1 個月,有的甚至超過1 個月[26,31,39]。較長的幼蟲浮游期以及海洋環境中缺少影響群體擴散的屏障可能是近親擬相手蟹群體能進行頻繁的基因交流,從而在較大地理范圍內表現出較低的遺傳分化水平的重要原因。類似的結果在雙齒近相手蟹[14]、環紋招潮(Uca annulipes)[40]的群體遺傳研究中也有報道。
本研究中單倍型中介網絡圖呈星狀結構、中性檢驗和核苷酸不配對分析的結果均暗示著近親擬相手蟹近期發生過群體擴張事件,根據核苷酸不配對τ值估算群體擴張時間大約發生在5.1 萬年前的更新世晚期。更新世的劇烈氣候波動,冰期和間冰期變化頻繁,冰川不斷進退消融,由此引起的海平面的升降對地球上的動植物地理空間分布格局和遺傳結構均產生了重大影響[41?42]。更新世海平面周期性的波動使海洋生物經歷了棲息地的反復收縮和擴張,尤其是末次冰盛期,大部分地區為巨大的冰原所覆蓋,西太平洋邊緣海平面下降,棲息地急劇收縮,迫使眾多海洋生物進入生物避難所,隨著冰川消退和氣溫回升,海平面上升,棲息地范圍擴大,幸存物種開始重新擴散并快速增長[41,43]。近親擬相手蟹的群體擴張時間大約發生在5.1 萬年前,推測近親擬相手蟹在更新世冰盛期經歷了棲息地劇烈收縮和種群數量的減少,隨著氣溫回升,海平面的上升和棲息地范圍的擴大,適宜的環境讓近親擬相手蟹幸存種群開始迅速擴張。不少研究也發現蟹類在更新世發生過群體擴張事件[12,14,40],海平面和溫度的變化可能導致了其群體遺傳結構的變化。由此可見,更新世的劇烈氣候變遷對海洋蟹類群體的遺傳結構和分布格局產生了巨大影響。
綜上所述,近親擬相手蟹12 個地理群體內和群體間的遺傳距離小,不同地理群體的單倍型沒有明顯的地理系譜結構,群體間的遺傳分化程度低,群體的遺傳變異主要來自群體內,這些暗示著近親擬相手蟹不同地理群體間存在著廣泛的基因交流。幼蟲浮游期長和較強的潛在擴散能力,海洋環境中缺少影響群體擴散的屏障以及近期的群體擴張事件,可能是近親擬相手蟹不同地理群體間存在著廣泛的基因交流,沒有形成顯著遺傳結構的重要原因,研究結果為近親擬相手蟹自然資源的保護及合理開發利用提供了一定理論依據。然而,浮游幼蟲的擴散是非常復雜的過程,易受到生活史特征、洋流、地理屏障等生物和非生物因素的影響,也有越來越多研究證明,海洋區域中存在著邊界和屏障,導致一些蟹類群體的基因交流受限和出現遺傳分化[26,44?45]。此外,近年來中國沿海環境不斷發生變化,特別是人為干擾的加劇,可能會對近親擬相手蟹這種生活于潮間帶區域的蟹類種群動態產生較大影響。本研究的樣品采集于2012?2013年,至于這幾年的沿海環境變化和人為活動是否會導致近親擬相手蟹群體的遺傳結構在不同時間尺度上出現差異尚不明確。穩定的遺傳變異對群體的演化及適應環境變化有重要作用,不同區域動物群體遺傳變異在時間尺度上具有一定的穩定性[46],但也有研究發現在不同的年份中,人為干擾嚴重的區域一些動物群體的遺傳結構出現分化,而遠離人為干擾環境中的群體,在不同年份中群體之間遺傳結構不存在差異[47]。由于本研究的采樣時間、地點和樣品量存在一定局限性,至于近親擬相手蟹群體在更大的地理范圍內是否存基因交流限制和顯著遺傳結構,以及在不同的時間尺度上群體遺傳結構是否存在差異,還需擴大研究范圍,深入研究群體遺傳變異的時空動態變化,并運用多種分子標記手段,以得出更全面和客觀的結論。
猜你喜歡
百科探秘·海底世界(2025年4期)2025-05-07 00:00:00
歌海(2024年6期)2024-03-18 00:00:00
兒童故事畫報·自然探秘(2022年3期)2022-04-27 00:17:10
湖南包裝(2022年6期)2022-02-11 09:01:18
幼兒園(2021年4期)2021-07-28 07:38:04
發明與創新·小學生(2020年7期)2020-07-18 16:13:39
瘋狂英語·新讀寫(2020年4期)2020-06-03 07:01:02
瘋狂英語·初中天地(2018年6期)2018-11-24 02:39:24
歌海(2018年4期)2018-05-14 12:46:15
少兒科學周刊·兒童版(2015年11期)2015-12-17 03:38:09
