漿果多糖的研究進(jìn)展
2021-03-17 08:25:38劉淑燕彭小燕陳美鏈姚閩娜
農(nóng)產(chǎn)品加工 2021年4期
劉淑燕,彭小燕,李 楊,陳美鏈,姚閩娜
(1.漳州科技職業(yè)學(xué)院茶與食品科技學(xué)院,福建漳州 363202;2.福建農(nóng)林大學(xué)食品科學(xué)學(xué)院,福建福州 350002;3.漳州職業(yè)技術(shù)學(xué)院食品工程學(xué)院,福建漳州 363200)
漿果指多汁肉質(zhì)的一類水果,常見的主要有葡萄、藍(lán)莓、枸杞、桑葚、黑穗醋栗等,一般具有獨(dú)特的口感和較高的營養(yǎng)價(jià)值,深受人們喜愛。漿果的果皮、果肉及籽粒中亦含有豐富的營養(yǎng)素和活性物質(zhì),賦予其多種生物活性功能[1-5]。
據(jù)大量研究表明,漿果的生物活性功能主要與其所含的多酚[6-7]、多糖有關(guān)。已有研究顯示,漿果多糖具有抗氧化、抗腫瘤、調(diào)節(jié)免疫、降血糖等多種生物功能[8-9],其生物活性功能與化學(xué)結(jié)構(gòu)密切相關(guān),而化學(xué)結(jié)構(gòu)又受提取方式影響[10]。因此,將國內(nèi)外學(xué)者對(duì)漿果多糖的提取純化、結(jié)構(gòu)鑒定、生物活性和分子修飾的研究進(jìn)行綜述,旨在為漿果多糖的進(jìn)一步開發(fā)利用提供參考。
1 漿果多糖的提取與純化
1.1 漿果多糖的提取
漿果粗多糖的提取方法主要有熱水浸提法、超聲波/微波輔助法、酶法、堿法、超高壓法、亞臨界水提取法等。
漿果多糖的提取方法見表1。
以上各種提取方法各有優(yōu)缺點(diǎn),傳統(tǒng)熱水浸提方法簡單易操作,但是存在時(shí)間長、提取率低且影響多糖活性等缺點(diǎn),近年來的一些高新技術(shù)應(yīng)用于多糖的提取,如超聲波微波法、酶法、超高壓等,縮短了提取時(shí)間,提高了提取效率,而且提取條件相對(duì)溫和,對(duì)多糖的活性影響較小,但是這些方法實(shí)施條件較為復(fù)雜,較難進(jìn)行產(chǎn)業(yè)化。目前,國內(nèi)外學(xué)者也在研究利用多種提取方法交聯(lián)使用,以提高多糖的提取效率。采用以上提取方法獲得的漿果多糖一般還含有蛋白、色素等物質(zhì),需要做純化處理。
1.2 漿果多糖的純化
多糖純化是指對(duì)提取的粗多糖混合物進(jìn)行分離而得到單一的多糖組分。目前,漿果多糖最常用的純化方法是離子交換柱層析和凝膠柱層析。離子交換柱層析多用于分離帶不同電荷的漿果多糖,能有效分離中性和酸性多糖組分,效率高,常用的交換劑有二乙氨基乙基(DEAE)-纖維素、DEAE-瓊脂糖 (Sepharose)和 DEAE- 葡聚糖 (Sephadex)等;凝膠柱層析對(duì)不同分子量的多糖組分進(jìn)行分級(jí),可達(dá)到精細(xì)分離的效果。通常為達(dá)到樣品純度的要求,經(jīng)常會(huì)將幾種方法結(jié)合使用[21]。陳春[22]采用DEAESepharose Fast Flow 離子交換層析柱對(duì)水提后脫色脫蛋白得到的桑葚多糖(MFPs)進(jìn)行分離純化,得到4 個(gè)多糖組分 (MFP-1、MFP-2、MFP-3 和MFP-4),又將MFP-3 通過Sephadex G-100 型凝膠柱進(jìn)一步純化得到MFP-3P,多糖含量從86.3%增加為94.2%。Kim D 等人[23]將從葡萄果皮熱水浸提液分離得到的葡萄粗多糖通過DEAE-Sepharose CL-6B、Sepharose CL-6B 和丙烯葡聚糖凝膠(Sephacryl)S-300 連續(xù)3 個(gè)層析柱,純化得到一種富集巨噬細(xì)胞的多糖。

表1 漿果多糖的提取方法
在較早的研究中,也有采用沉淀法[24]對(duì)漿果多糖進(jìn)行分級(jí)純化,其分離效率高,但是很難達(dá)到高純度要求;目前,也有研究采用色譜法[25]對(duì)漿果多糖進(jìn)行精細(xì)分級(jí),但是其耗時(shí)長、效率低、成本高。
2 漿果多糖結(jié)構(gòu)的研究
漿果多糖是一類分子結(jié)構(gòu)復(fù)雜的生物大分子,因其單糖組成、分子量、環(huán)構(gòu)象及基團(tuán)間相互作用等的多樣性,其結(jié)構(gòu)鑒定具有一定挑戰(zhàn)性,需利用相關(guān)化學(xué)方法并結(jié)合應(yīng)用現(xiàn)代各種光譜技術(shù)進(jìn)行較為全面的解析。目前,結(jié)構(gòu)研究較多的有黑穗醋栗多糖、枸杞多糖、桑葚多糖等。
2.1 黑穗醋栗多糖結(jié)構(gòu)的研究進(jìn)展
黑穗醋栗多糖結(jié)構(gòu)的研究主要集中在其單糖的組成及比例分析、糖環(huán)形式、糖苷的異構(gòu)形式、表面結(jié)構(gòu)及構(gòu)象分析,對(duì)其糖苷連接順序和連接方式的研究較少。已有研究顯示,黑穗醋栗多糖為吡喃多糖,其單糖組成主要有葡萄糖(glucose,Glc)、阿拉伯糖(arabinose,Ara)、甘露糖和半乳糖等,不具有三螺旋結(jié)構(gòu)。
任中杰[26]通過超聲波輔助法提取黑穗醋栗多糖,并對(duì)純化得到的多糖組分BCPⅡ進(jìn)行結(jié)構(gòu)解析,測(cè)得其是一種含有乙酰氨基結(jié)構(gòu)的β 型吡喃多糖,分子量為51.88 kDa,單糖組成為葡萄糖(Glucose,Glc)、 阿 拉 伯 糖 (Arabinose, Ara)、 半 乳 糖(Galactose,Gal)和甘露糖(Mannose,Man),物質(zhì)量比為4.10∶2.36∶0.106∶0.26。徐雅琴等人[27-28]分別采用雙水相萃取法、超聲波法提取分離黑穗醋栗果實(shí)多糖(BCP),并通過傅里葉變換紅外光譜(Fourier transform infrared spectroscopy,F(xiàn)T-IR)、氣相色譜(Gas chromatography,GC)和高效液相色譜(High performance liquid chromatography,HPLC)進(jìn)行結(jié)構(gòu)分析,測(cè)得其單糖組成主要為阿拉伯糖、鼠李糖(Rhamnose,Rha)、木糖(Xylose,Xyl)、甘露糖、葡萄糖、半乳糖,但不同的提取方法得到的BCP 物質(zhì)的量比例不同,紅外光譜圖顯示其含有多糖的特征吸收峰,可能含有α -糖苷鍵及吡喃糖環(huán),經(jīng)掃描電鏡 (Scanning electron microscope,SEM)和剛果紅測(cè)試表明,BCP-1 具有蜂窩狀結(jié)構(gòu),但不具有三螺旋結(jié)構(gòu)。
2.2 枸杞多糖結(jié)構(gòu)的研究進(jìn)展
已有研究表明枸杞多糖為酸性多糖,糖苷部分由半乳糖、葡萄糖、甘露糖、阿拉伯糖、鼠李糖、木糖等單糖組成,此外還含有氨基酸、半乳糖醛酸(Galacturonic acid,GalA)、葡萄糖醛酸(Glucuronic acid,GlcA)[29],但其單糖的摩爾比、糖鏈結(jié)構(gòu)、連接位置等存在不同[10],不具有三股螺旋結(jié)構(gòu)[30]。
Zhou L 等人[31]采用復(fù)合酶法提取枸杞多糖,利用DEAE SepharoseTMFast Flow、 Sephacryl S-300 HR 進(jìn)行純化得到酸性多糖LBP1B-S-2,利用部分酸水解分析、甲基化分析、IR 和核磁共振光譜(Nuclear Magnetic Resonance,NMR)方法對(duì)該多糖進(jìn)行結(jié)構(gòu)分析,測(cè)得其平均分子量為80 kDa,單糖組成n(Rha)∶n(Gal)∶n(Ara)∶n(GlcA)為3.13∶39.37∶53.55∶3.95,主鏈結(jié)構(gòu)由 (1 →6)-linked β-D-Galp、 (1→3)-linked β-D-Galp 組成,支鏈由T-linked β-L-Araf、 (1 →6)-linked β-D-Galp、T-linked α-L-Araf、 (1 →4)-linked β-D-GlcpA、T-linked β-L-Rhap、 (1→5)-linked α-L-Araf 和T-linked β-D-Galp 組成,并連接在主鏈的C-3 或C-6 位置。Liu W 等人[32]對(duì)枸杞中酸性多糖p-LBP 進(jìn)行結(jié)構(gòu)鑒定得知,該多糖是一種均勻的果膠分子雜多糖,多糖含量為98.85%,平均分子量為64 kDa。其單糖組分主要有阿拉伯糖、巖藻糖(Fucose,F(xiàn)uc)、鼠李糖、半乳糖、木糖、葡萄糖醛酸、半乳糖醛酸,摩 爾 比 為 54.84 ∶1.00 ∶6.44 ∶22.98 ∶4.05 ∶2.95 ∶3.35 ∶136.98。 p-LBP 的 主 鏈 是 重 復(fù) 的 (1 →4)-α-GalpA 結(jié) 構(gòu) , 但 是 部 分 區(qū) 域 是 由 (1 →4)-α-GalpA 和 (1→2)-α-Rhap 交替連接的;支鏈由 (1→4)-β-Galp、 (1→3)-β-Galp 或 (1→5)-α-Araf 組成,連接在 (1→2)-α-Rhap 的 C-4 位置。
2.3 桑葚多糖結(jié)構(gòu)的研究進(jìn)展
桑葚多糖的結(jié)構(gòu)組成因受產(chǎn)地、品種等的影響而存在差異。已有研究表明,桑葚多糖的單糖組分主要有阿拉伯糖、鼠李糖、葡萄糖、半乳糖,還含有木糖、巖藻糖、甘露糖等單糖,以及糖醛酸、硫酸基、吡喃己糖殘基,部分桑葚多糖中存在α,β -糖苷鍵,不具有三股螺旋結(jié)構(gòu)。
陳春[22]從新疆黑桑中提取桑葚多糖(MFPs)并進(jìn)行分離純化,得到 MFP-1、MFP-2、MFP-3、MFP-4 共4 個(gè)多糖組分,剛果紅試驗(yàn)表明4 組分均不具有三股螺旋構(gòu)象,SEM 顯示各組分結(jié)構(gòu)及外貌形態(tài)不盡相同。將MFP-3 通過凝膠柱層析純化得到MFP3P,利用HPLC 法測(cè)得多糖含量為94.2%,分子量為136.6 kDa,單糖組成主要為阿拉伯糖、鼠李糖、葡萄糖、半乳糖及甘露糖,其對(duì)應(yīng)的摩爾比分別為21.51%,25.98%,13.60%,23.10%,16.35%。通過高碘酸氧化、Smith 降解分析、甲基化-氣質(zhì)色譜、1H-NMR 和13C-NMR 等方法,推斷構(gòu)成糖鏈的單糖殘基類型主要包括 (1-2)-linked α-L-Rha、(1 →6)-linked α-D-Glc、 (1 →3)-linked β-LRha、 (1→3)-linked α-D-Gal 和 (1→)-linked α-L-Ara 單元。李賽娟[33]對(duì)產(chǎn)于浙江的桑葚進(jìn)行提取純化,并對(duì)其中的 4 種均一多糖 (FMP-6-S2,F(xiàn)MP-6-S4,F(xiàn)MP-6-S1,F(xiàn)MP-6-H)進(jìn)行結(jié)構(gòu)分析,得到FMP-6-S2 為RG-I 型多糖,單糖組成的摩爾比n(Rha)∶n(Gal)∶n(Ara)∶n(GalA)為30.86∶28.70∶15.61∶24.78,其主鏈由 (1→2)-α-L-Rhap與(1→4)-α-D-Galp A 交替連接而成,支鏈取代在鼠李糖的 C-4 位,由末端由 (1→3→6)-β-DGalp,T-β-D-Galp,T-α-L- Araf , (1 →4)-β-D-Galp 和 (1→5)-α-L-Araf 構(gòu)成;FMP-6-S4 為一個(gè)蛋白聚糖,其單糖組成的摩爾比n(Glc)∶n(Rha)∶n(Gal)∶n(Ara)∶n(GalA)為3.87∶5.80 ∶3.70 ∶4.50 ∶62.13, 主 鏈 由 1 →4 連 接 的α-D-Galp A 與 1→2 連接的 α-L-Rhap 組成,己烯糖醛酸 (α-L-Hexp A)和 β-D-Galp A 通過 α-D-Galp A 的 C-3 位連接到主鏈, (1→5)-α-L-Araf、α-LAraf、β-D-Glcp、β-D-Galp 或 (1→6)-β-D- Glcp殘基通過 α-L-Rhap 的 C-4 位連接到主鏈;FMP-6-S1 由半乳糖、阿拉伯糖、鼠李糖、半乳糖醛酸、葡萄糖醛酸和葡萄糖組成,其摩爾比為38.05∶20.54∶19.02∶17.86∶3.03∶1.50,主鏈由 (1→4)-α-D-Galp A 與 (1→2)-α-L-Rhap 交替連接構(gòu)成,支 鏈 由 T-β-D-Glc Ap, T-β-D-Galp (或 Glcp),(1→6)-β-D-Galp, (1→4→6)-β-D-Galp, (1→3)-β-D-Galp, (1→4)-β-D-Galp, (1→5)-α-LAraf 和 T-α-L-Araf 構(gòu)成,通過 (1→2)-α-L-Rhap的C4 位連接到主鏈;FMP-6-H 是以(1→4)-連接的直鏈半乳糖醛酸聚糖。
2.4 其他多糖結(jié)構(gòu)的研究進(jìn)展
目前,文獻(xiàn)也有報(bào)道其他漿果多糖的結(jié)構(gòu)研究。Cordeiro C A R 等人[8]對(duì)黑莓酒中分離得到的3 種多糖組分 (BWPs、BWPFs、BWPFp)進(jìn)行結(jié)構(gòu)解析,BWPs 中的主要組分有甘露聚糖、II 型阿拉伯半乳聚糖和I 型鼠李糖半乳糖醛酸;BWPFp 屬于甘露聚糖,主鏈由1→6 連接 α-Manp 組成, O-2 位置被支鏈(1→2)-α-D-Manp 取代;BWPFs 為 II 型阿拉伯半乳聚糖和I 型鼠李糖半乳糖醛酸,II 型阿拉伯半乳聚糖的主鏈由(1→3)-β-D-Galp 組成,支鏈由連接在O-6 的(1→6)-β-D-Galp 組成,I 型鼠李糖半乳糖醛酸由[→4)-α-d-GalpA-(1→2)-α-l-Rhap-(1→]n 組成。徐雅琴等人[34]研究藍(lán)靛果多糖結(jié)構(gòu)藍(lán)靛果多糖為酸性雜多糖,單糖組成主要為半乳糖醛酸、鼠李糖、阿拉伯糖、甘露糖、葡萄糖、半乳糖,不具有三股螺旋結(jié)構(gòu),呈現(xiàn)無定形片狀結(jié)構(gòu)。
綜上所述,受產(chǎn)地、品種、提取方式等的影響,不同漿果中的多糖結(jié)構(gòu)不同,其結(jié)構(gòu)、單糖組成、化學(xué)結(jié)構(gòu)、分子量等存在差異,但多種漿果多糖均不具有三股螺旋結(jié)構(gòu)。目前,關(guān)于漿果多糖的構(gòu)象特征特別是其分子尺寸及分子形態(tài)的研究還較欠缺,有待更深入的解析與闡述。
3 漿果多糖的生物活性
目前,有關(guān)漿果多糖生物活性的研究主要集中在其抗氧化、增強(qiáng)免疫調(diào)節(jié)功能、抗腫瘤、降血糖、調(diào)節(jié)腸道菌群等方面。
3.1 抗氧化活性
不受控制的細(xì)胞代謝中自由基的增加會(huì)對(duì)防御細(xì)胞造成各種損傷,引起免疫功能下降,從而導(dǎo)致腫瘤、炎癥和心血管疾病等的發(fā)生[35]。有研究已表明漿果多糖具有一定的抗氧化活性,可以有效清除自由基。陳春[22]對(duì)桑葚進(jìn)行超聲提取、脫蛋白、脫色處理得到3 個(gè)多糖組分(MFP、MFP-1、MFP-2)進(jìn)行抗氧化活性研究,結(jié)果表明3 組分對(duì)2,2-聯(lián)氮-二(3 - 乙基 - 苯并噻唑 - 6 - 磺酸)二銨鹽 (2,2'-azinobis- (3-ethylbenzthiazoline-6-sulphonate,ABTS)自由基清除能力都高于90%,而MFP-1 的氧化自由基吸收能力、羥自由基(·OH)清除能力、1 -二苯基- 2 -三硝基苯肼(1,1-diphenyl-2-picrylhydrazyl,DPPH)自由基清除能力優(yōu)于MFP 和MFP-2。
Amagase H 等人[36]研究枸杞多糖 (LBP)的體內(nèi)抗氧化作用發(fā)現(xiàn),其抗氧化作用是通過刺激內(nèi)源性因子的作用,從而達(dá)到抗氧化功效。Amagase H 等人[37]以50 名55~72 歲健康的中國成年人為研究對(duì)象,發(fā)現(xiàn)研究對(duì)象食用LBP 后體內(nèi)血清中谷胱甘肽過氧化物酶和超氧化物歧化酶的含量顯著提高,血清丙二醛顯著降低,從而提高體內(nèi)抗氧化功效。
具有抗氧化活性的漿果多糖見表2。
由表2 可知,不管是枸杞多糖、黑穗醋栗多糖,還是藍(lán)莓多糖,均具有一定的抗氧化活性,其作用方式主要是通過清除自由基以達(dá)到抗氧化效果[38-39]。
3.2 免疫調(diào)節(jié)活性
有研究表明漿果多糖能激活樹突狀細(xì)胞、巨噬細(xì)胞等固有性免疫細(xì)胞,從而發(fā)揮免疫調(diào)節(jié)作用。Chen Z 等人[40]研究發(fā)現(xiàn)枸杞多糖(50 mg/kg,腹膜內(nèi))增加了CD40,CD80,CD86 和 II 類主要組織相容性復(fù)合體分子的表達(dá),上調(diào)巨噬細(xì)胞的內(nèi)吞和吞噬作用,并通過RAW264.7 巨噬細(xì)胞激活核轉(zhuǎn)錄因子和激活蛋白,誘導(dǎo)白細(xì)胞介素的表達(dá),增強(qiáng)腫瘤壞死因子的產(chǎn)生,且呈劑量依賴性方式。Cordeiro C A R 等人[8]首次證明了黑莓多糖對(duì)細(xì)菌脂多糖(LPS)誘導(dǎo)的RAW 264.7 巨噬細(xì)胞的抗炎作用。LPS 可刺激巨噬細(xì)胞有效降低NO 和促炎性細(xì)胞因子(IL-1β,TNF-α)的產(chǎn)生,黑莓多糖可增強(qiáng)淋巴細(xì)胞體液免疫和細(xì)胞免疫,從而起到抗炎效果,可能是一種很有前途的免疫調(diào)節(jié)化合物。駱新等人[41]研究桑葚多糖(MP)對(duì)環(huán)磷酰胺誘導(dǎo)的免疫低下小鼠免疫功能的調(diào)節(jié)作用,發(fā)現(xiàn)灌胃MP 中、高劑量組的小鼠其脾臟、胸腺指數(shù)升高,脾臟淋巴細(xì)胞的轉(zhuǎn)化功能和抗體生成細(xì)胞的功能顯著增強(qiáng),MP 高劑量組的血清溶血素水平升高顯著,說明MP 對(duì)環(huán)磷酰胺誘導(dǎo)的免疫低下模型小鼠具有免疫保護(hù)作用。孫希云[42]采用小鼠足趾腫脹法進(jìn)行遲發(fā)型過敏反應(yīng)(Delayed-type hypersensitivity,DTH)試驗(yàn)來研究藍(lán)莓多糖的免疫活性,結(jié)果表明高劑量組 (400 mg/kg/d)對(duì)綿羊紅細(xì)胞(Ssheep blood cells,SRBC)誘導(dǎo)小鼠 DTH 具有顯著的抑制作用。
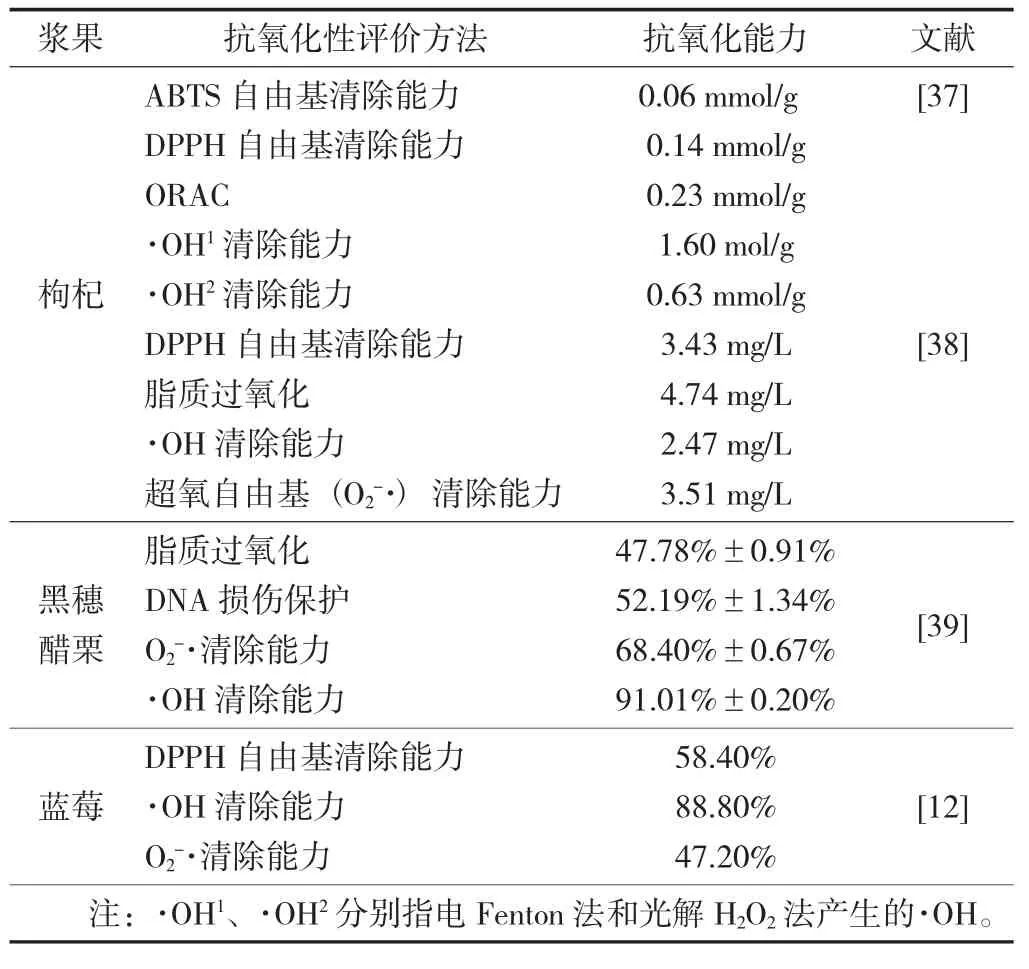
表2 具有抗氧化活性的漿果多糖
此外,有研究發(fā)現(xiàn)漿果多糖還通過促進(jìn)細(xì)胞因子的生成來增強(qiáng)免疫作用。Liu C J 等人[43]從桑椹和草莓中分離多糖,研究表明適當(dāng)濃度的草莓多糖(SP)和桑葚多糖(MP)處理可顯著增加脾細(xì)胞的增殖。MP 刺激 IL-2、IL-4 和 IL-5 的生成,SP 刺激TNF-α、IL-10 和IL-12 的形成,從而保護(hù)原代免疫細(xì)胞免于凋亡。結(jié)果顯示,MP 在體外具有更好的細(xì)胞增殖和抗凋亡潛力,而SP 在體外具有更好的抗炎潛力。Jeff H 等人[44]研究發(fā)現(xiàn)巴西莓多糖在牛、小鼠和人外周血單核細(xì)胞培養(yǎng)物中誘導(dǎo)了強(qiáng)大的γδT 細(xì)胞刺激活性,分子量最高的多糖在體外最活躍。當(dāng)巴西莓多糖在體內(nèi)遞送時(shí),可誘導(dǎo)肺和腹膜組織中的髓樣細(xì)胞,并增強(qiáng)肺中IL-12 的產(chǎn)生。
3.3 抗腫瘤活性
目前,研究漿果多糖的抗腫瘤方式主要有2 種:一是通過提高機(jī)體免疫能力而起作用的間接方式;二是直接殺傷或抑制腫瘤細(xì)胞的直接方式。Sun X Y等人[9]研究藍(lán)莓多糖(BBP3-1)在S180 荷瘤小鼠體內(nèi)的抗腫瘤作用,結(jié)果表明BBP3-1 能夠顯著提高脾臟和胸腺指數(shù),增加巨噬細(xì)胞吞噬作用,增強(qiáng)NK 細(xì)胞活性,促進(jìn)淋巴細(xì)胞增殖轉(zhuǎn)化,刺激淋巴細(xì)胞釋放 IL-2、TNF-α、干擾素 (interferon,IFN)-γ,增加荷瘤小鼠的免疫防御系統(tǒng)。因此,推測(cè)BBP3-1 可能是通過增強(qiáng)機(jī)體的免疫作用而間接起到抗腫瘤作用。Kim D 等人[23]從葡萄果皮中分離得到的葡萄多糖在體外試驗(yàn)中可誘導(dǎo)腹腔巨噬細(xì)胞產(chǎn)生多種細(xì)胞因子(IL-6,IL-10 和IL-12),通過靜脈內(nèi)注射葡萄多糖顯著增強(qiáng)了自然殺傷細(xì)胞對(duì)小鼠淋巴瘤細(xì)胞Yac-1的細(xì)胞毒性,可劑量依賴性地顯著抑制結(jié)腸26-M3.1癌細(xì)胞的肺轉(zhuǎn)移。Zhang S H 等人[45]通過試驗(yàn)證實(shí)枸杞多糖(LRP3-S1)可抑制胰腺癌細(xì)胞的生長,同時(shí)能減弱BxPC-3 細(xì)胞的侵襲能力,并下調(diào)p-FAK,p-AKT,p-GSK-3β 和 p-p38 MAP 激酶的蛋白表達(dá)。Ryoji T 等人[46]研究發(fā)現(xiàn)口服黑醋栗汁和黑醋栗多糖(CAPS)對(duì)荷瘤小鼠實(shí)體瘤的生長分別有45%和51%的延緩作用,CAPS 可顯著增加細(xì)胞因子IL-2,IL-4,IL-10 和IFN-γ 的分泌,對(duì)腫瘤細(xì)胞有一定的直接殺傷作用。
3.4 降血糖活性
漿果多糖的降血糖機(jī)制主要有以下幾個(gè)方面[22]:①調(diào)節(jié)代謝酶的活性,抑制葡萄糖的吸收或擴(kuò)散速度;②增強(qiáng)外周組織對(duì)胰島素的敏感性,增強(qiáng)對(duì)葡萄糖的攝取能力;③改善胰島β 細(xì)胞功能,使其免受損傷,促進(jìn)胰島素分泌。Chen C 等人[16]研究發(fā)現(xiàn)桑葚多糖對(duì)α -淀粉酶及α -葡萄糖苷酶的活性均具有一定的抑制作用,其也能夠吸附葡萄糖,抑制葡萄糖擴(kuò)散,起到延緩葡萄糖吸收的作用,從而降低餐后血糖。李朝暉等人[47]研究發(fā)現(xiàn)枸杞多糖能夠增強(qiáng)3T3-L1 脂肪細(xì)胞對(duì)葡萄糖的攝取而抑制肝糖產(chǎn)生,保護(hù)鏈脲佐菌素?fù)p傷的NIT-L1 胰島β 細(xì)胞,且能降低小腸刷狀緣對(duì)葡萄糖的吸收和消化道內(nèi)α -葡萄糖苷酶的活性,從而起到較好的體外降血糖作用。
3.5 調(diào)節(jié)腸道菌群
研究表明,漿果多糖與腸道菌群之間是相互作用的,一方面漿果多糖被腸道菌群降解,產(chǎn)生具有一定生物活性的代謝產(chǎn)物;另一方面漿果多糖能夠促進(jìn)腸道微生物的多樣性并調(diào)節(jié)其比例[48]。
王莉等人[49]研究不同劑量黑枸杞多糖對(duì)菌群人源化小鼠(HFA-小鼠)模型的腸道微生物的影響,結(jié)果表明3 個(gè)劑量組(50,100,200 mg/kg)的黑枸杞多糖均可抑制HFA-小鼠腸道內(nèi)腸球菌和腸桿菌的繁殖,刺激原有乳酸桿菌和益菌雙歧桿菌的繁殖,還能促進(jìn)腸黏膜sIgA 的分泌,從而改善HFA-小鼠腸道菌群環(huán)境,具有益生元作用。李賽娟[33]研究桑葚多糖FMP-6-S2 及其降解產(chǎn)物FMP-6-S2-01a 對(duì)多形擬桿菌的影響,結(jié)果表明2 種多糖均能促進(jìn)多形擬桿菌的生長,多形擬桿菌也可利用2 種多糖產(chǎn)生短鏈脂肪酸丁酸和乙酸,而短鏈脂肪酸為腸道細(xì)菌和腸道黏膜細(xì)胞提供能量,發(fā)揮維持腸道穩(wěn)態(tài)的調(diào)控作用。
隨著研究的深入,漿果多糖還被證實(shí)具有抗衰老[33]、保肝[50]、抗病毒[51]等生物活性,展現(xiàn)出較好的開發(fā)利用前景。然而,部分從天然生物中分離得到的多糖,其生物活性較弱,需要對(duì)其進(jìn)行分子修飾,從而可提高某一生物活性或新增一種生物活性[52]。
4 漿果多糖的分子修飾及其生物活性
目前,多糖修飾的方法主要有物理修飾法、化學(xué)修飾法和生物修飾法等。多糖的物理修飾是指通過物理手段使多糖降解,得到分子量更低的多糖衍生物,如輻照法和超聲波降解法;化學(xué)修飾是指通過化學(xué)手段對(duì)其結(jié)構(gòu)進(jìn)行修飾,從而獲得具有更新或高的生物活性的多糖衍生物[53],如硫酸化法、氧化降解法、羧甲基化法、乙酰化法等;生物修飾法應(yīng)用最廣泛的是酶法,主要是利用多糖剪切酶的剪切作用使多糖降解。
王杏[52]對(duì)桑葚多糖進(jìn)行羧甲基化處理,結(jié)果顯示其羧甲基化衍生物具有更好的抗化學(xué)性肝損傷作用。陳春[22]和嚴(yán)婭娟等人[54]均對(duì)天然桑葚多糖進(jìn)行硒化修飾,結(jié)果表明其硒化衍生物抗氧化活性明顯優(yōu)于天然桑葚多糖。此外,陳春[22]還發(fā)現(xiàn)硒化桑葚多糖具有較強(qiáng)的降血糖活性。Xu Y Q 等人[55-56]采用了氧化降解法(Fe2+和H2O2體系)、硫酸化法、超聲輻射法[39]和羧甲基化法等4 種方法對(duì)黑穗醋栗多糖進(jìn)行改性,生物活性測(cè)定結(jié)果均表明,改性多糖比天然多糖具有更高的抗氧化能力和α -淀粉酶抑制活性。這些結(jié)果表明,對(duì)漿果多糖進(jìn)行適當(dāng)?shù)姆肿有揎椏梢蕴岣咂渖锘钚浴?/p>
漿果多糖分子修飾之所以能提高多糖生物活性的原因主要有以下幾個(gè)方面:①降低多糖的分子量,使其黏度降低,水溶性升高,生物活性得以充分發(fā)揮;②增加或改變多糖的功能基團(tuán),使其親水性增強(qiáng),生物活性得以提高;③改變多糖的糖鏈分支,使其親水基暴露,增加其水溶性,提高其生物活性。
5 結(jié)語
綜上所述,漿果多糖是漿果中的一類重要的功能物質(zhì),關(guān)于其的研究主要集中在多糖的提取、純化、結(jié)構(gòu)分析、生物活性分析及分子修飾等方面。高新技術(shù)的應(yīng)用使?jié){果多糖的提取和純化研究取得了一定成果,然而由于漿果多糖結(jié)構(gòu)組成比較復(fù)雜,其空間結(jié)構(gòu)受多方因素影響,關(guān)于漿果多糖結(jié)構(gòu)與生物活性之間關(guān)系的研究還遠(yuǎn)遠(yuǎn)落后于海藻多糖和真菌多糖等的研究廣度和深度,這也導(dǎo)致人們不能完全認(rèn)識(shí)水果的營養(yǎng)保健價(jià)值,阻礙了漿果多糖的有效利用。我國漿果資源十分豐富,其加工副產(chǎn)物的高效利用也是推動(dòng)漿果行業(yè)發(fā)展的一大動(dòng)力,因此研究漿果多糖高效提純方法、構(gòu)效關(guān)系及通過各種高新技術(shù)定向修飾改造漿果多糖,使其具有更高的活性功能,將是今后漿果多糖的研究重點(diǎn)。
猜你喜歡
天天愛科學(xué)(2022年9期)2022-09-15 01:12:54
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
天天愛科學(xué)(2022年4期)2022-05-23 12:41:48
當(dāng)代水產(chǎn)(2022年3期)2022-04-26 14:26:56
哲學(xué)評(píng)論(2021年2期)2021-08-22 01:53:34
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
航空世界(2020年10期)2020-01-19 14:36:20
科技傳播(2019年22期)2020-01-14 03:06:54
中華詩詞(2019年7期)2019-11-25 01:43:04
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
