擬穴青蟹CYP302a1基因的克隆及表達(dá)模式分析
2021-03-18 01:53:26張林姿張鳳英劉志強(qiáng)馬春艷馬凌波
海洋漁業(yè) 2021年1期
關(guān)鍵詞:分析
張林姿,趙 明,張鳳英,王 偉,劉志強(qiáng),馬春艷,諶 微,王 珂,馬凌波
(1.上海海洋大學(xué)水產(chǎn)與生命學(xué)院,上海 201306;2.中國水產(chǎn)科學(xué)研究院東海水產(chǎn)研究所,農(nóng)業(yè)農(nóng)村部遠(yuǎn)洋與極地漁業(yè)創(chuàng)新重點(diǎn)實(shí)驗(yàn)室,上海 200090)
節(jié)肢動物中,周期性蛻皮對節(jié)肢動物的生長、繁殖和形態(tài)發(fā)生起著重要作用。周期性蛻皮受到多種環(huán)境和自身因素的影響,其中最主要的是由類固醇激素濃度控制,而蛻皮激素(ecdysone,Ecd)是最重要的類固醇激素[1-3]。蛻皮激素首先由BUTENANDT和KARLSON[4]1954年從蛾蛹中分離得到,隨后Ecd的化學(xué)結(jié)構(gòu)也被闡明[5]。昆蟲中蛻皮激素的主要活性形式是20-羥基蛻皮酮(20-hydroxyecdysone,20E),其誘導(dǎo)和調(diào)節(jié)蛻皮周期中一系列生命進(jìn)程的變化[1]。1965年HUBER和HOPPE[5]發(fā)現(xiàn)前胸腺為Ecd的主要分泌合成器官,現(xiàn)已證明,節(jié)肢動物周期性蛻皮主要是由類固醇蛻皮激素的周期性波動調(diào)節(jié)的[6-7]。此外,Ecd的濃度變化也與再生、變態(tài)、卵子發(fā)生等[8-13]密切相關(guān)。因此,對Ecd的代謝調(diào)控研究具有重要意義。
甲殼動物的Ecd主要由Y器官(Y-organ,YO)合成分泌[14],其合成與昆蟲類似。Ecd的合成可以分為兩個(gè)階段:首先,食物中的膽固醇在腸道吸收后,通過血淋巴運(yùn)輸?shù)結(jié)O,膽固醇在Neverland基因、“黑盒子”(包括CYP307a1/CYP307a2)基因編碼酶的作用下催化成為5βdiketol(3D2,22,25dE),此過程在所有十足目物種中保持一致[15];然后,5β-diketol在多種細(xì)胞色素 P450 (包 括 CYP306Al、CYP302Al 和CYP315Al)的催化下轉(zhuǎn)化為Ecd。隨后,Ecd釋放到外周組織,由20-羥化酶(CYP314A1)催化而成其主要的活性形式 20-羥基蛻皮酮(20-hydroxyecdysone,20E)[16-17]。
CYP302A1催化C22的羥基化,是Ecd合成通路中的重要催化酶。在果蠅(Drosophila melanogaster)中通過敲除CYP302a1(dib)基因獲得的dib突變胚胎,發(fā)現(xiàn)其Ecd和20E滴度非常低,且通過表型分析發(fā)現(xiàn)dib突變胚胎在一些后期形態(tài)發(fā)生過程存在嚴(yán)重缺陷,如頭部退化及腸發(fā)育異常等,證明CYP302a1基因在蛻皮激素合成中起重要作用[18];而在家蠶(Bombyx mori)中發(fā)現(xiàn)CYP302a1基因的表達(dá)變化與血淋巴內(nèi)蛻皮類固醇滴度的變化密切相關(guān)[19]。甲殼動物中目前僅有三疣梭子蟹(Portunus trituberculatus)克隆獲得CYP302a1基因,研究發(fā)現(xiàn),CYP302a1基因在YO的表達(dá)量顯著高于其他組織,且在蛻皮周期中YO的表達(dá)變化與Ecd的滴度變化相同,而CYP302a1的表達(dá)高峰期早于蛻皮激素高峰期,說明CYP302a1基因在三疣梭子蟹的蛻皮激素合成中起著重要作用[20]。
擬穴青蟹(Scylla paramamosain)隸屬于梭子蟹科(Portunidae),青蟹屬,廣泛分布于我國東南沿海地區(qū),營養(yǎng)價(jià)值豐富、經(jīng)濟(jì)價(jià)值較高,是我國重要的海水經(jīng)濟(jì)蟹類之一[21-22]。蛻皮調(diào)控是甲殼動物研究中的熱點(diǎn)問題之一,目前甲殼動物蛻皮激素合成通路的相關(guān)報(bào)告較少,因此,對擬穴青蟹蛻皮調(diào)控通路的解析具有重要意義。本研究通過序列分析,結(jié)合組織分布、幼體時(shí)空表達(dá)、蛻皮周期變化規(guī)律對擬穴青蟹的CYP302a1基因進(jìn)行了克隆鑒定,以期為相關(guān)研究提供基礎(chǔ)數(shù)據(jù)支撐。
1 材料與方法
1.1 實(shí)驗(yàn)材料
擬穴青蟹溞狀幼體Ⅰ期至仔蟹Ⅰ期不同發(fā)育時(shí)期樣本取自東海水產(chǎn)研究所海南瓊海養(yǎng)殖基地,不同時(shí)期幼體樣本的鑒定參考曾朝曙等[23]關(guān)于幼體的形態(tài)學(xué)描述,幼體取樣選取兩次蛻皮中間的時(shí)間點(diǎn)進(jìn)行收集。用于組織表達(dá)分析的樣本分別取自瓊海基地養(yǎng)殖的卵巢發(fā)育Ⅲ期的雌性和精巢發(fā)育處于精母細(xì)胞期的雄性青蟹,性腺發(fā)育分期參考上官步敏等[24]和YE等[25]的研究。不同蛻皮時(shí)期的雄蟹取自江蘇省南通維爾思水產(chǎn)科技有限公司,暫養(yǎng)于實(shí)驗(yàn)室循環(huán)水養(yǎng)殖系統(tǒng),選取體質(zhì)量(BW)為25~55g的雄性擬穴青蟹,參考徐賓朋[26]的研究,將擬穴青蟹蛻皮周期分為蛻皮后期(A、B期)、蛻皮間期(C期)、蛻皮前期(D)和蛻皮期(E)4個(gè)階段。取每個(gè)蛻皮時(shí)期的YO組織,每個(gè)時(shí)期5只蟹,加RNA保存液(生工生物工程(上海)股份有限公司)保存至-80℃,用于RNA提取。
1.2 實(shí)驗(yàn)方法
1.2.1 擬穴青蟹CYP302a1的cDNA克隆
從轉(zhuǎn)錄組測序結(jié)果中獲得了擬穴青蟹CYP302a1的cDNA序列,通過分析發(fā)現(xiàn)其具有完整的開放閱讀框(ORF),在ORF兩端設(shè)計(jì)引物(CYP302a1-F: GGCAGGTGTCTGAAGGAACA,CYP302a1-R:ACATATTCACCTCAGCTAGTGC),以YO的cDNA為模板對其進(jìn)行了驗(yàn)證。除試劑盒提供的引物外,本研究中使用的其他引物均委托上海杰李生物技術(shù)有限公司合成。
1.2.2 序列分析
使用ORF Finder(http://www.ncbi.nlm.nih.gov/gorf/gorf.html)查找序列的開放閱讀框并翻譯,在 GenBank 上下載多個(gè)物種的CYP302a1基因的氨基酸序列,使用DNAMAN軟件進(jìn)行比對,分析其與其他物種的相似性;利用ExPASy ProtParam (https://web.expasy.org/protparam)預(yù)測蛋白質(zhì)的分子量和等電點(diǎn);使用Signal 5.0 Server(http://www.cbs.dtu.dk/services/SignalP)預(yù)測蛋白質(zhì)是否存在信號肽;利用在線TMHMM 工具(http://www.cbs.dtu.dk/services/TMHMM)預(yù)測蛋白質(zhì)是否存在跨膜區(qū)域。
1.2.3 系統(tǒng)進(jìn)化樹的構(gòu)建
在GenBank數(shù)據(jù)庫中下載節(jié)肢動物中多個(gè)物種的CYP302a1基因的氨基酸序列及P450家族的其他參與20E合成及代謝的基因編碼的氨基酸序列,包括CYP306A1、CYP307A1、CYP315A1、CYP314A1、CYP18A1。 使用MEGA7[27]對所選序列進(jìn)行比對,鄰接法構(gòu)建發(fā)育樹,Bootstrap設(shè)置為1 000,使用在線工具Interactive Tree Of Life(https://itol.embl.de)對進(jìn)化樹進(jìn)行注釋修飾[28]。
1.2.4Sp-CYP302a1基因的組織表達(dá)和時(shí)空分布分析
提取成體雌蟹的表皮、中腸、胸神經(jīng)節(jié)、血淋巴、肌肉、YO、心臟組織;成體雄蟹的表皮、血淋巴、中腸、肝胰腺、心臟、精巢、YO組織;雄性蟹不同蛻皮時(shí)期的YO和心臟,從溞狀幼體I期到仔蟹Ⅱ期不同幼體發(fā)育時(shí)期樣本的總RNA,使用總RNA提取試劑盒(北京艾德萊生物技術(shù)有限公司)提取RNA,提取后使用分光光度計(jì)進(jìn)行定量,1%的瓊脂糖凝膠電泳檢測RNA的完整性。對符合要求的RNA使用DNAase I處理后進(jìn)行反轉(zhuǎn)錄(上海TOYOBO)。熒光定量使用試劑為SYBR green PCR master mix(大連TAKARA)。反應(yīng)體系和程序均參考ZHAO等[29]的方法進(jìn)行。
由于18S rRNA在擬穴青蟹的多種不同樣本中能穩(wěn)定表達(dá),適合作為擬穴青蟹基因表達(dá)分析的內(nèi)參基因,本研究使用18S rRNA作為內(nèi)參基因[30],Sp-CYP302a1的 特 異 性 引 物 為Sp-CYP302a1-Q-F(TCTCACAGAGAGTGCCGAGA)和Sp-CYP302a1-Q-R(ATGCAGGCAACCAATCAACG),采用標(biāo)準(zhǔn)曲線法進(jìn)行分析計(jì)算檢測Sp-CYP302a1基因在各個(gè)樣本中的相對表達(dá)量。
1.2.5 統(tǒng)計(jì)分析
表達(dá)量所有數(shù)據(jù)均以平均值±標(biāo)準(zhǔn)差(mean±SD)表示,使用SPSS 17.0軟件對數(shù)據(jù)one-way ANOVA分析,用Duncan氏法進(jìn)行比較,取P<0.05為差異顯著,P<0.01為差異極顯著。
2 結(jié)果與分析
2.1 擬穴青蟹Sp-CYP302a1序列及進(jìn)化樹分析
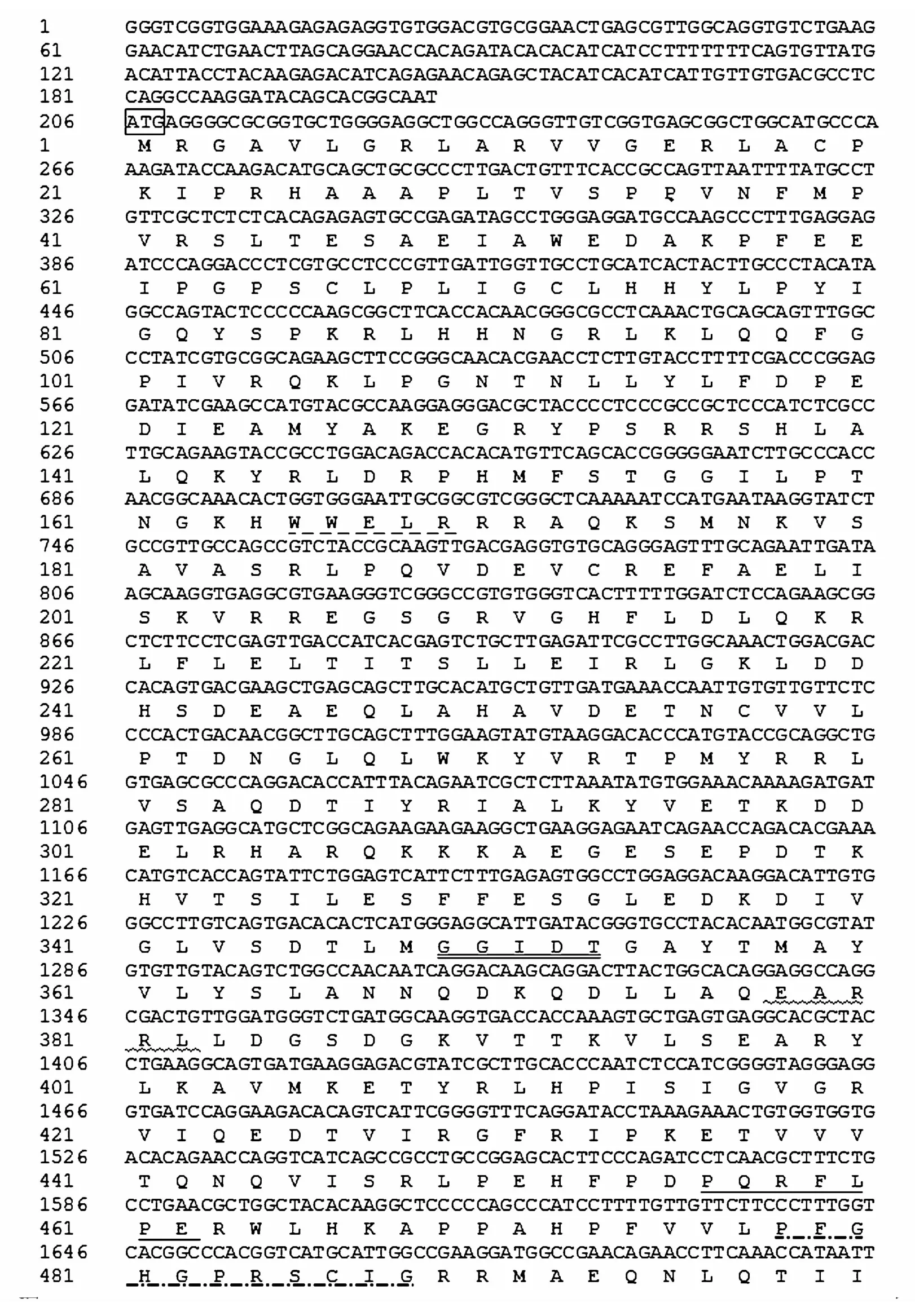
本研究克隆獲得了擬穴青蟹CYP302a1基因,命名為Sp-CYP302a1(GenBank 接 收 號:MN782363)。Sp-CYP302a1的cDNA 序列長為3 032 bp,開放閱讀框(OFR)為1 617 bp,可編碼538個(gè)氨基酸。此外,有205 bp的5′UTR和1 210 bp的3′UTR。ExPASy ProtParam預(yù)測結(jié)果顯示蛋白分子式為C2729H4344N780O782S17,平均分子質(zhì)量為61.14 kDa,等電點(diǎn)(pI)為8.76,帶負(fù)電荷的氨基酸(天冬氨酸+谷氨酸)67個(gè),帶正電荷氨基酸(精氨酸 +賴氨酸)73 個(gè)。其中,預(yù)測的CYP302a1蛋白的大小與其他脊椎動物和無脊椎動物細(xì)胞色素P450(56~62 kDa)的大小一致[31]。Signal 5.0 Server和TMHMM分析發(fā)現(xiàn)其氨基酸序列無信號肽序列和跨膜結(jié)構(gòu)域。序列分析顯示該氨基酸序列含有helix-K、helix-C、helix-I、PERF及heme-binding共5個(gè)P450特征保守區(qū)域[32](圖1)。
多序列比對分析發(fā)現(xiàn),Sp-CYP302a1的氨基酸序列與三疣梭子蟹的一致度最高(91%),其次為凡納濱對蝦(Penaeus vannamei)、木黃蜂(Orussus abietinus)和歐洲熊蜂 (Bombus terrestris),一致度分別為56% 、46% 和46%,。有4個(gè)保守區(qū)域的非保守序列與三疣梭子蟹及凡納濱對蝦完全一致,包括helix-C、helix-K、helix-I和heme-binding;多處非保守區(qū)域的氨基酸序列與三疣梭子蟹和凡納濱對蝦一致性極高甚至完全一致,包括氨基酸108~111、204~214、288~343 和384 ~401 等(圖2)。而Sp-CYP302a1的氨基酸序列與其他所比對物種一致度極低。
系統(tǒng)進(jìn)化樹顯示不同的CYP基因分別聚為一類,分別為CYP306a1、CYP307a1、CYP315a1、CYP314a1、CYP302a1和CYP18a16個(gè)大類。Sp-CYP302a1與其他物種的CYP302a1聚為一類,且Sp-CYP302a1與三疣梭子蟹的進(jìn)化關(guān)系最近,其次為凡納濱對蝦(圖3)。
2.2 Sp-CYP302a1基因的表達(dá)模式分析
qRT-PCR檢測結(jié)果顯示,Sp-CYP302a1基因在擬穴青蟹成體蟹不同組織中的表達(dá)差異較顯著。Sp-CYP302a1在雌性和雄性蟹中的組織分布較為相似,在所檢測的組織中均有表達(dá),但在YO中的表達(dá)量明顯高于其他組織,其他組織中表達(dá)量均較低,統(tǒng)計(jì)學(xué)檢驗(yàn)為差異顯著(P<0.05);此外,Sp-CYP302a1在雌性擬穴青蟹中表達(dá)量明顯比雄性個(gè)體中高(圖4)。
在幼體發(fā)育過程中,CYP302a1基因在溞狀幼體Ⅲ期(Z3)最高,而在其他時(shí)期均相對較低(圖5)。

作者選用雄性擬穴青蟹的YO來檢測Sp-CYP302a1在蛻皮周期中表達(dá)的變化,以期初步揭示其在擬穴青蟹蛻皮調(diào)控中的作用。在一個(gè)蛻皮周期內(nèi),Sp-CYP302a1表達(dá)量呈顯著變化,在蛻皮后期(A、B期)表達(dá)量最低,隨后開始上升,至蛻皮前期(D期)達(dá)到最大值,隨后開始下降,但在蛻皮期(E期)的表達(dá)量仍高于A期和B期(圖6)。
3 討論
細(xì)胞色素P450基因CYP302a1已從多個(gè)節(jié)肢動物中克隆并證明其在蛻皮調(diào)控中具有重要作用[19-20,33]。本研究從擬穴青蟹中獲得的基因CYP302a1具有細(xì)胞色素P450基因的5個(gè)保守結(jié)構(gòu)域(圖2),證明Sp-CYP302a1屬于P450家族基因,此外Sp-CYP302a1序列推導(dǎo)的氨基酸序列與三疣梭子蟹和凡納濱對蝦的CY302A1聚為一小支(圖3),而后與其他物種的CY302A1聚為一支,而 P450 其他成員基因編碼的氨基酸CYP306A1、CYP307A1、CYP315A1、CYP314A1、CYP18A1分別聚為一支,結(jié)構(gòu)域和進(jìn)化聚類結(jié)果初步證明所獲得的為擬穴青蟹CYP302a1基因。
雌性和雄性擬穴青蟹Sp-CYP302a1基因均在YO中具有明顯高于其他組織的表達(dá)(圖4),與YO是甲殼動物蛻皮激素合成的主要器官的論斷相吻合[14],這也與三疣梭子蟹的結(jié)果一致[20]。此外,Sp-CYP302a1在雌性擬穴青蟹中表達(dá)量明顯比雄性個(gè)體中高,在果蠅中也發(fā)現(xiàn)了這種現(xiàn)象,這顯示蛻皮激素的濃度可能在不同性別中具有差異,蛻皮激素在不同性別的功能值得進(jìn)一步深入探索。此外,Sp-CYP302a1在幼體發(fā)育過程中的Z3時(shí)期出現(xiàn)表達(dá)量峰值,而在其他時(shí)期Sp-CYP302a1的表達(dá)量相對較低,在實(shí)際生產(chǎn)過程中,Z1和Z2期主要以輪蟲為餌料,自Z3期開始會增加鹵蟲幼體作為其生物餌料,這些現(xiàn)象是否具有一定聯(lián)系值得進(jìn)一步探討。
蛻皮激素與甲殼動物的蛻皮活動緊密聯(lián)系,是蛻皮活動的重要調(diào)控因子,對凡納濱對蝦[34]、羅氏沼蝦(Macrobrachium rosenbergii)[35]、三疣梭子蟹[36]等的相關(guān)研究表明其在蛻皮周期中呈規(guī)律性變化,且分別在蛻皮前期的不同亞期達(dá)到高峰。Sp-CYP302a1在蛻皮周期YO中的表達(dá)呈現(xiàn)顯著變化,在蛻皮前期出現(xiàn)峰值,這與其他甲殼動物中蛻皮周期內(nèi)蛻皮激素的濃度變化趨勢及三疣梭子蟹的CYP302a1表達(dá)模式一致[20],表明CYP302a1在擬穴青蟹蛻皮激素的合成中也具有重要作用。

圖2 擬穴青蟹CYP302a1氨基酸序列與其他物種CYP302a1序列比對Fig.2 Alignment of the deduced am ino acid sequence of S.paramamosa in CYP302a1w ith other species

圖3 節(jié)肢動物部分P450家族基因系統(tǒng)進(jìn)化樹Fig.3 Phylogenetic tree of part arthropod P450 fam ily

圖4 Sp-CYP302a1基因在不同組織中的表達(dá)Fig.4 Tissue distribution analysis of Sp-CYP302a1

圖5 Sp-CYP302a1基因在不同發(fā)育階段中的表達(dá)Fig.5 Exp ression pattern of Sp-CYP302a1during different larval development stages

圖6 Sp-CYP302a1在雄蟹蛻皮周期YO中的相對表達(dá)Fig.6 Relative expression of Sp-CYP302a1 in YOduring molting cycle in male S.paramamosain
綜上,本研究首次從擬穴青蟹中克隆獲得Sp-CYP302a1基因的cDNA序列,通過生物信息學(xué)及表達(dá)模式綜合分析初步證明Sp-CYP302a1參與了Ecd的合成,研究結(jié)果可為擬穴青蟹蛻皮相關(guān)研究提供基礎(chǔ)數(shù)據(jù)。
猜你喜歡
現(xiàn)代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機(jī)設(shè)計(jì)與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
當(dāng)代經(jīng)濟(jì)研究(2016年5期)2016-12-01 03:12:05
現(xiàn)代農(nóng)業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫(yī)藥現(xiàn)代遠(yuǎn)程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學(xué)學(xué)報(bào)(社會科學(xué)版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
