ARTP誘變選育γ-聚谷氨酸高產(chǎn)菌株快速篩選方法的建立
2021-03-23 07:17:18劉丹丹臧毅鵬王夢夢岳文瑾余晨銳陳俊柳聶光軍
安徽工程大學(xué)學(xué)報 2021年1期
劉丹丹,臧毅鵬,王 利,王夢夢,岳文瑾,余晨銳,陳俊柳,聶光軍
(安徽工程大學(xué) 生物與食品學(xué)院,安徽 蕪湖 241000)
γ-聚谷氨酸(γ-Polyglutamic acid,γ-PGA)是一種由谷氨酸單體聚合而成的高分子多聚物,具有無毒和可降解性,被廣泛應(yīng)用于食品行業(yè)。如作為益生菌的低溫保護(hù)劑,提高益生菌在生產(chǎn)過程中的存活率;在面粉加工過程中,γ-PGA可以減緩淀粉老化,提高面制品的彈性和韌性等。目前,γ-PGA主要通過微生物發(fā)酵生產(chǎn),但目前菌株生產(chǎn)效率滿足不了市場對γ-PGA的需求,導(dǎo)致γ-PGA的價格居高不下。為提升菌株生產(chǎn)效率,各種誘變方法被使用,其中物理射線誘變相對于化學(xué)誘變具有簡單、環(huán)境污染小等特點。常用的物理誘變主要通過紫外線、X射線等射線輻照微生物,以改變DNA,但這些方法存在誘變效率低以及可控性差等問題。近幾年興起的常壓室溫等離子體(Atmospheric and Room Temperature Plasma,ARTP)誘變技術(shù)因具有突變率高,對人體和環(huán)境無毒害,易操作等優(yōu)點,常被用于微生物菌種誘變。
然而,基于ARTP誘變選育γ-PGA高產(chǎn)菌株的過程中,阻礙誘變選育效率的可能是其篩選方法的低效。面對大量誘變菌株,使用傳統(tǒng)方法逐一測定每個菌株的產(chǎn)量則需要耗費過多的精力與時間。γ-PGA產(chǎn)生菌在生長過程中,由于各種蛋白酶的作用,往往在其周圍的培養(yǎng)基上形成一個透明圈,透明圈的大小則反映了蛋白酶的活性,而γ-PGA在生產(chǎn)過程中又與蛋白酶存在著偶聯(lián)關(guān)系。為此,在篩選一株γ-PGA高產(chǎn)菌株并在對其進(jìn)行鑒定的基礎(chǔ)上,分析酪蛋白平板初篩、多孔板復(fù)篩(24孔板與48孔板)和搖瓶復(fù)篩之間的關(guān)系,嘗試建立透明圈與菌落面積的比例(圈徑比)與γ-PGA產(chǎn)量之間的關(guān)系,簡化篩選流程,通過圈徑比大小直觀快速篩選γ-PGA高產(chǎn)突變株,快速提升ARTP誘變選育γ-PGA高產(chǎn)菌的效率。
1 材料與方法
1.1 材料與試劑
生產(chǎn)γ-PGA的出發(fā)菌株(市售納豆粉中篩選獲得,命名為MA1);L-谷氨酸(國藥化學(xué)試劑有限公司);γ-聚谷氨酸(四川拙誠日化科技有限公司,純度大于95%);細(xì)菌基因組提取試劑盒、Taq酶、引物和相關(guān)PCR試劑(上海生工生物工程有限公司)。
1.2 菌株鑒定
通過對菌株的形態(tài)學(xué)研究、菌落形態(tài)觀察、菌株的生理生化實驗3個方面對菌株進(jìn)行鑒定。將MA1接種到固體培養(yǎng)基(牛肉膏5 g/L、胰蛋白胨5 g/L、NaCl 5 g/L、瓊脂15 g/L、pH 7.0),37 ℃培養(yǎng)24 h,觀察菌落形態(tài)。將MA1接種到種子培養(yǎng)基(牛肉膏3 g/L、葡萄糖5 g/L、胰蛋白胨5 g/L、NaCl 5 g/L、pH 7.0),37 ℃培養(yǎng)24 h,革蘭染色鏡檢觀察菌株形態(tài)。將MA1接種到酪蛋白培養(yǎng)基(脫脂奶粉50 g/L、瓊脂15 g/L、pH 7.0),37 ℃培養(yǎng)24 h,觀察菌落形態(tài)。將MA1接種到液體發(fā)酵培養(yǎng)基(蔗糖28.87 g/L、胰蛋白胨17.56 g/L、L-谷氨酸15.26 g/L、pH 7.0),37 ℃培養(yǎng)24 h后,將發(fā)酵液滴入纖維蛋白平板(2%瓊脂糖、20 IU凝血酶溶液、0.3 mg纖維,總體積為10 mL)中,觀察發(fā)酵液的纖維降解能力。
基于SDS-堿裂解法原理,應(yīng)用細(xì)菌基因組提取試劑盒提取MA1基因組,應(yīng)用引物(27F/1492R)擴(kuò)增16S rDNA,并對其進(jìn)行測序(合肥通用生物),測序結(jié)果通過blast比對,應(yīng)用MEGA 7.0制作進(jìn)化樹,從分子水平鑒定MA1。應(yīng)用引物NK-F(GGGGTACCGTATGAAAATAGTTATTTCGAGTCTCTACG)和NK-R(AACTGCAGTCCGGTGCTTGTGAAGATTTC)PCR擴(kuò)增aprN(納豆激酶編碼基因),并測序比對,鑒定MA1與納豆菌(Bacillus Subtilis Natto)的關(guān)系。
1.3 菌株篩選方法的研究
(1)初篩方法:選用混濁、易被菌種分解利用的酪蛋白平板接種MA1,每4 h測量菌體直徑及水解圈直徑,計算圈徑比(水解圈與菌體直徑之比),分析圈徑比與MA1的γ-PGA產(chǎn)量之間的變化關(guān)系。
(2)復(fù)篩方法:分別采用24孔板、48孔板接種對數(shù)期的種子液,孔板接種量和裝液量比例同100 mL搖瓶發(fā)酵(7.5%接種量/30 mL發(fā)酵培養(yǎng)基),37 ℃、200 rpm發(fā)酵培養(yǎng),每4 h取樣檢測γ-PGA產(chǎn)量,以研究孔板體積對γ-PGA產(chǎn)量的影響,確定多孔板類型。在此基礎(chǔ)上,進(jìn)一步應(yīng)用24孔板發(fā)酵(37 ℃、200 rpm),每4 h取樣檢測γ-PGA產(chǎn)量,考察其與相同條件下?lián)u瓶發(fā)酵之間的相關(guān)性,確定最佳復(fù)篩方法。
1.4 ARTP誘變選育
(1)誘變菌懸液的制備:種子液37 ℃、200 rpm培養(yǎng)至對數(shù)期,取l mL菌液4 500 rpm離心去上清液,加入等量生理鹽水重懸菌體,OD在0.8~1.0之間待用。
(2)生長及發(fā)酵曲線的測定:將對數(shù)期的種子液以7.5%的接種量接入裝液量為30 mL的發(fā)酵培養(yǎng)基中(100 mL錐形瓶),37 ℃、200 rpm震蕩培養(yǎng),每4 h取樣測菌濃和γ-PGA產(chǎn)量。
(3)ARTP誘變:將8個裝有990 μL生理鹽水的EP管依次置于ARTP誘變儀(無錫源清天木生物科技公司)相應(yīng)凹槽的底部,紫外滅菌20 min,將8個鋪滿10 μL菌液的ARTP載片依次置于ARTP誘變儀相應(yīng)凹槽中,誘變時間依次為0 s、15 s、30 s、45 s、60 s、90 s、120 s和160 s(誘變條件:99.99%的高純氦氣,電源功率120 W、氣體流量10 SLM、照射距離2 mm、溫度<30 ℃)。誘變結(jié)束后,震蕩洗脫EP管2 min,使載片上的菌體完全混于生理鹽水中,再將EP管置于4 ℃冰箱中,保存2 h,避免自省對突變損傷進(jìn)行修復(fù)。最后將誘變液稀釋一定倍數(shù)(原液菌濃度約為1.08×10cfu/mL),取100 μL分別涂布于固體培養(yǎng)基和酪蛋白平板,每組3個平行,37 ℃培養(yǎng),記錄固體培養(yǎng)基平板上的菌落數(shù)量,根據(jù)下式計算致死率,并繪制致死率曲線。測量酪蛋白板上的圈徑,并計算圈徑比。
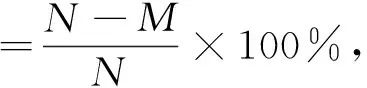
N
為誘變固體平板上的菌落數(shù);M
為誘變處理后平板上的菌落數(shù)。(4)遺傳穩(wěn)定性研究:將篩選的突變株傳代培養(yǎng)5次,檢測γ-PGA的產(chǎn)量以考察其遺傳穩(wěn)定性。
1.5 γ-PGA產(chǎn)量測定
參照Zeng等報道的方法。10 000 rpm離心發(fā)酵液10 min,取上清液;加入4倍體積冰乙醇,置于4 ℃冰箱中過夜,10 000 rpm離心10 min取沉淀;將沉淀55 ℃下烘干至恒重,加入與發(fā)酵液等體積的蒸餾水復(fù)溶;溶解完全后,10 000 rpm離心10 min去除不溶物,將上清適當(dāng)稀釋后測定OD216(TU-1810PC紫外可見光分光光度計,北京普析通用儀器有限責(zé)任公司),根據(jù)γ-PGA標(biāo)準(zhǔn)曲線計算γ-PGA產(chǎn)量。
2 結(jié)果與分析
2.1 菌株的鑒定
對菌株MA1進(jìn)行形態(tài)學(xué)鑒定如圖1所示。圖1a顯示菌株MA1是一種類圓形,邊緣呈鋸齒狀,表面褶皺不透明,在菌體中央有粘液并且可產(chǎn)孢子的菌種;對菌株MA1的菌落形態(tài)進(jìn)行革蘭氏染色顯微觀察。圖1b為桿狀紫色,證明菌株MA1為革蘭氏陽性細(xì)菌;對菌株MA1進(jìn)行生理生化實驗的鑒定。圖1c顯示在酪蛋白板上出現(xiàn)水解圈,說明該菌株含有蛋白酶,對酪蛋白有降解能力。圖1d顯示在纖維蛋白平板上有明顯的水解圈形成,說明該菌株產(chǎn)生了纖溶蛋白酶可降解纖維蛋白原以及纖維蛋白。觀察結(jié)果與枯草芽孢桿菌168(模式菌)生理生化特性基本相同,因而,初步推斷菌株MA1屬于可生產(chǎn)蛋白酶的芽孢桿菌。
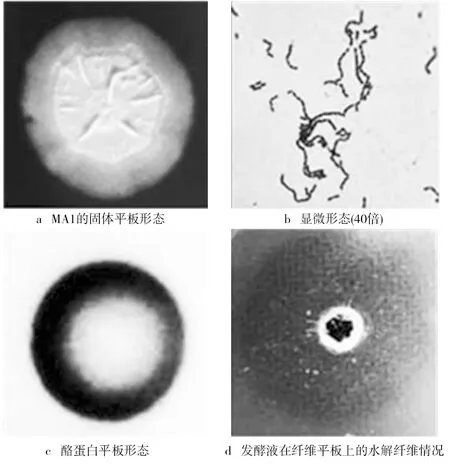
圖1 菌株鑒定
對菌株MA1進(jìn)行分子生物學(xué)鑒定如圖2所示。由圖2可見,在約1 500 bp處有一條明顯的條帶,與預(yù)期16S rDNA的大小相符。經(jīng)測序后得到菌株MA1的16S rDNA序列,通過與GenBank已報道的序列比對,利用MAGE 7.0軟件繪制菌株MA1系統(tǒng)發(fā)育樹(見圖3),菌株MA1與枯草芽孢桿菌(Bacillus subtilis)的同源性高達(dá)99%,由此確定菌株MA1為枯草芽孢桿菌。進(jìn)一步PCR擴(kuò)增該菌株的aprN基因,圖2b顯示該擴(kuò)增片段與已報道aprN基因序列(1473 bp)基本一致。測序結(jié)果顯示,所得NK基因與Nakamura報道的aprN基因序列(GI:262756)相同。因為aprN基因序列具有高度的特異性,主要產(chǎn)生菌為納豆枯草芽孢桿菌(Bacillus Subtilis Natto,簡稱納豆菌)。因此,可以確定菌株MA1屬于納豆菌。
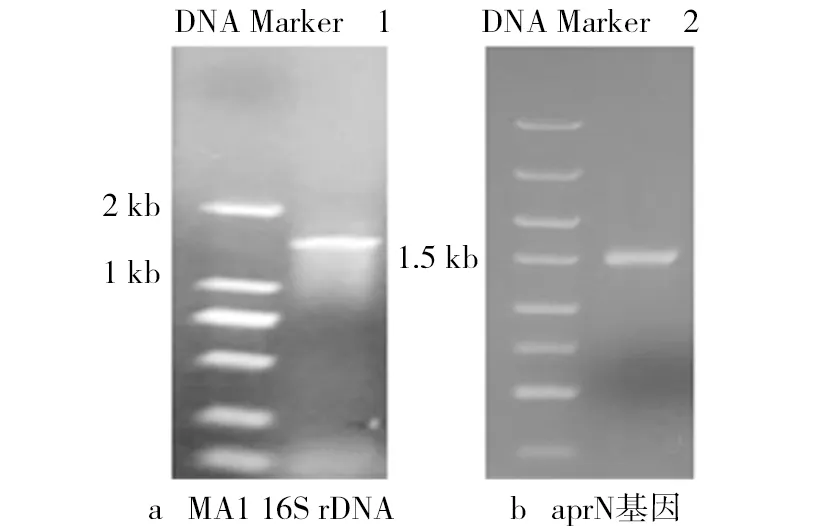
圖2 MA1 16S rDNA和aprN基因PCR產(chǎn)物電泳圖譜

圖3 菌株MA1 16S rDNA的系統(tǒng)發(fā)育樹
2.2 篩選方法確定
對數(shù)生長期的納豆菌代謝旺盛,菌體比生長速率最大,細(xì)胞的數(shù)量呈指數(shù)上升,且抗不利環(huán)境的能力最強(qiáng)。因此,微生物發(fā)酵和誘變選擇對數(shù)生長期的菌株。發(fā)酵液與種子液中的培養(yǎng)基成分、接種量及菌齡都不相同,菌株在發(fā)酵液中的生長曲線就會受到一定的影響。通過測定菌體生長與γ-PGA產(chǎn)量,可以更直接地觀察兩者間的關(guān)系,對菌株篩選與分析發(fā)酵液中γ-PGA生產(chǎn)過程有重要的意義。
種子液菌株MA1的生長及發(fā)酵曲線如圖4所示。由圖4可知,菌株前4 h增殖緩慢,處于延滯期;4~12 h菌體急劇增殖,比生長速率最大,處于對數(shù)期;14~36 h菌體數(shù)量處于動態(tài)平衡,此時為穩(wěn)定期;36 h后菌體數(shù)量開始下降,進(jìn)入衰亡期。為了滿足足夠的接種量、旺盛的菌種生長活力及實驗的統(tǒng)一性,后期誘變和發(fā)酵選擇10~12 h的菌種液。相對于菌體生長,γ-PGA生產(chǎn)進(jìn)程明顯滯后。細(xì)胞生長處于延滯期時無γ-PGA產(chǎn)生,細(xì)胞對數(shù)期γ-PGA產(chǎn)量增加緩慢,穩(wěn)定期γ-PGA生產(chǎn)明顯加速。說明γ-PGA生產(chǎn)與菌體增殖存在一定關(guān)聯(lián),這種次級代謝產(chǎn)物與初級產(chǎn)物聯(lián)系更為緊密。之前的研究結(jié)果顯示,隨菌體的生長納豆激酶的含量增加,菌體細(xì)胞處于對數(shù)期,納豆激酶含量達(dá)到最大,菌體生長進(jìn)程與納豆激酶等蛋白酶的生產(chǎn)偶聯(lián),γ-PGA生產(chǎn)由蛋白酶水解底物產(chǎn)生的代謝物質(zhì),由此推斷γ-PGA生產(chǎn)與蛋白酶的生產(chǎn)也可能存在一定關(guān)聯(lián)。孔板對菌株γ-PGA產(chǎn)量的影響、24孔板與搖瓶發(fā)酵的相關(guān)性以及搖瓶發(fā)酵γ-PGA產(chǎn)量與菌株酪蛋白平板上的菌圏比之間的變化趨勢如圖5所示。由圖5a顯示,24和48孔板中γ-PGA產(chǎn)量變化趨勢基本相同。其中,24孔板中的γ-PGA產(chǎn)量明顯較高,這是由于納豆菌是好氧菌,48孔板內(nèi)菌液的表面接觸氧氣較24孔板小,從而影響菌株生長代謝過程中的溶氧量,因此,24孔板更有利于菌株 MA1的生長代謝,相對于孔板溶氧,搖瓶溶氧效果更好。由圖5b顯示,24孔板和同期搖瓶的γ-PGA產(chǎn)量相近(R=0.978 8),說明24孔板具有較好的溶氧量,滿足γ-PGA生產(chǎn)對溶氧的需求。因此,為提升誘變復(fù)篩效率,可用24孔板培養(yǎng)替代搖瓶培養(yǎng)。
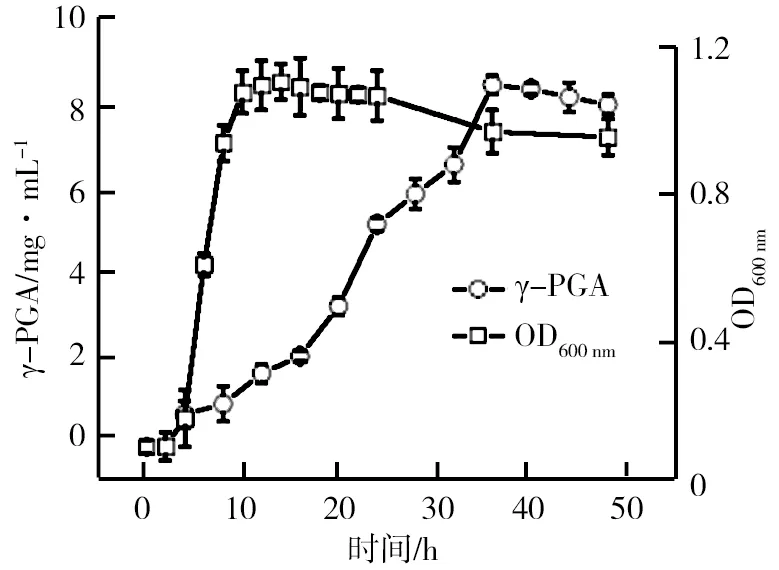
圖4 種子液菌株MA1的生長及發(fā)酵曲線
面對誘變出現(xiàn)的大量菌株,使用快速有效的篩選方法是提高誘變選育效率的關(guān)鍵。納豆菌發(fā)酵過程中會產(chǎn)生蛋白酶,在其水解酪蛋白后會在菌落周圍形成一個酪蛋白水解圈,水解圈與菌落大小的比值可以間接表示蛋白酶活力的大小,這一方法被廣泛用于產(chǎn)酶菌種的初篩。為了驗證前文關(guān)于蛋白酶與γ-PGA生長之間存在關(guān)聯(lián)的推斷,菌液涂布于酪蛋白平板后,觀察圈徑比與γ-PGA產(chǎn)量之間的變化關(guān)系,由圖5c顯示,圈徑比隨著菌體的生長在16~24 h迅速增加,與酪蛋白的生長高度偶聯(lián),因為次級代謝產(chǎn)物的生產(chǎn)一般滯后于初級代謝產(chǎn)物,因此γ-PGA生產(chǎn)曲線相對于酪蛋白生產(chǎn)同步后移,且生長趨勢基本相同,間接證明了菌株MA1蛋白酶的生產(chǎn)與γ-PGA產(chǎn)量可能存在關(guān)聯(lián)。
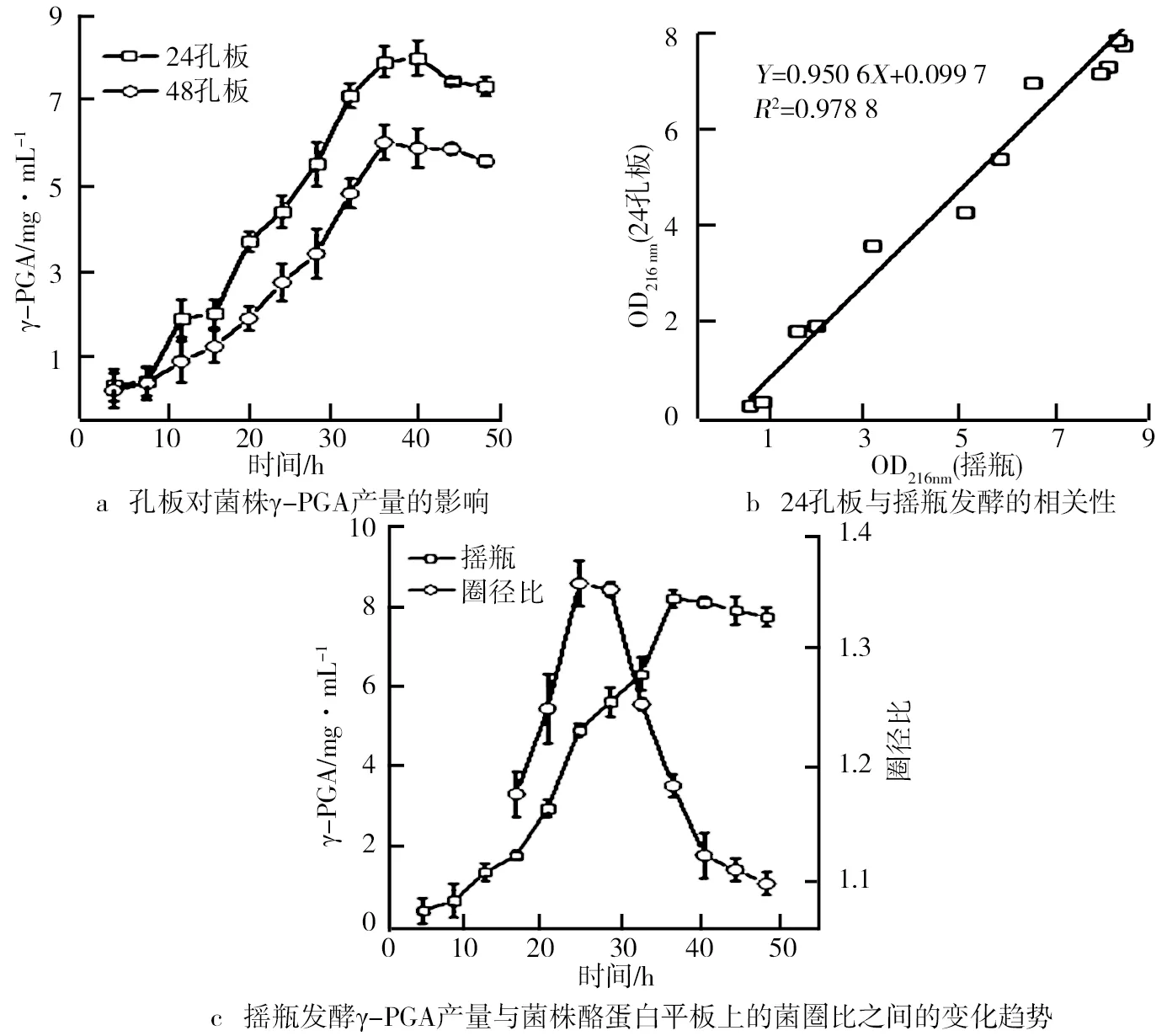
圖5 孔板對菌株γ-PGA產(chǎn)量的影響、24孔板與搖瓶發(fā)酵的相關(guān)性以及搖瓶發(fā)酵γ-PGA產(chǎn)量與菌株酪蛋白平板上的菌圏比之間的變化趨勢
2.3 ARTP誘變選育
致死率是獲得優(yōu)良突變株的關(guān)鍵參數(shù)。ARTP輻照MA1的致死曲線如圖6a所示。由圖6a可知,誘變致死率隨誘變時間的增加在不斷升高,誘變時間為90 s時,致死率達(dá)到88.16%。誘變300 s時,致死率為100%。由于突變具有隨機(jī)性,正負(fù)突變與誘變時間無明確的規(guī)律可循,但因為誘變時間越長,會使DNA鏈和氫鍵斷裂的程度、胸腺嘧啶二聚體形成的程度及宿主修復(fù)后造成的差錯程度加大,從而造成菌株損傷加重,與出發(fā)菌株相比變化更大。因此,為提高突變株篩選效率,誘變時間選為270 s。經(jīng)過5輪誘變(出發(fā)菌株為實驗室前期篩選菌株MA1),在23株菌株中篩選獲得8株相對高產(chǎn)的突變株(見圖6b)。其中,3株為正突變株,m3的γ-PGA產(chǎn)量最高,達(dá)到9.22 g/L,較出發(fā)菌株提高了22.03%。誘變菌的產(chǎn)量提高率相較于張浩誘變菌產(chǎn)量提高的42.11%偏低,但其出發(fā)菌株的γ-PGA產(chǎn)量僅為本實驗出發(fā)菌株產(chǎn)量的十分之一。對突變株m3進(jìn)行連續(xù)5次發(fā)酵后,其產(chǎn)量在9.2 g/L左右(見圖6c),說明突變體遺傳穩(wěn)定性較好。分析23株突變體的圈徑比與其γ-PGA產(chǎn)量之間的關(guān)系,如圖6d所示,樣本量為24個(包括出發(fā)菌株),圈徑比和γ-PGA產(chǎn)量之間R
值為0.64。一般而言,當(dāng)樣本量超過9個,R
值達(dá)到0.7,或當(dāng)樣本量超過25個,R
值達(dá)到0.4,都可認(rèn)為兩變量間存在相關(guān)性。據(jù)此,我們認(rèn)為菌株圈徑比和γ-PGA產(chǎn)量存在相關(guān)性,為基于酪蛋白透明圈篩選γ-PGA高產(chǎn)菌株提供了依據(jù)。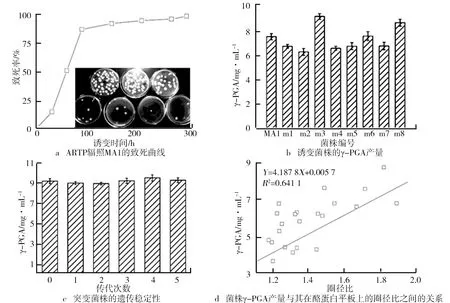
圖6 ARTP輻照MA1的致死曲線、誘變菌株的γ-PGA產(chǎn)量、突變菌株的遺傳穩(wěn)定性以及菌株γ-PGA產(chǎn)量與其在酪蛋白平板上的圈徑比之間的關(guān)系
3 結(jié)論
研究在篩選到一株高產(chǎn)γ-PGA納豆菌的基礎(chǔ)上,系統(tǒng)分析酪蛋白板初篩、多孔板復(fù)篩和搖瓶復(fù)篩,發(fā)現(xiàn)多孔板復(fù)篩與搖瓶復(fù)篩結(jié)果一致。其中,多孔板復(fù)篩與酪蛋白平板形成的納豆菌的圈徑比具有線性關(guān)聯(lián),這為直接應(yīng)用酪蛋白平板初篩代替原用的平板初篩加多孔板復(fù)篩提供了依據(jù)。基于酪蛋白平板的透明圈快篩方法,從ARTP輻照的菌群中篩選出一株γ-PGA高產(chǎn)株,產(chǎn)量較出發(fā)菌株提升了22%,且遺傳穩(wěn)定性良好,表明酪蛋白平板的透明圈快篩方法有效,這大幅壓縮了篩選的時間,顯著提升了菌株誘變選育的效率。
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
今日農(nóng)業(yè)(2021年14期)2021-11-25 23:57:29
小讀者(2021年2期)2021-03-29 05:03:48
石油與天然氣地質(zhì)(2021年1期)2021-02-22 14:14:44
今日農(nóng)業(yè)(2020年20期)2020-11-26 06:09:10
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
中國果業(yè)信息(2019年10期)2019-11-13 01:21:34
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
