懸鈴木幼苗的初生維管系統演化結構研究
2021-03-27 01:18:50王桂文張立秋王世東由繼紅劉丹丹陸靜梅
東北師大學報(自然科學版) 2021年1期
王桂文,高 偉,張立秋,王世東,李 巖,由繼紅,劉丹丹,朱 軍,陸靜梅
(1.東北師范大學生命科學學院,吉林 長春 130024;2.公安部物證鑒定中心,北京 100038)
懸鈴木為懸鈴木科(Platanaceae)懸鈴木屬(PlatanusLinn.)植物.懸鈴木科是一個只有1屬約11種的小科[1].我國南北各地栽培懸鈴木科植物約3種,多作為行道樹和庭蔭樹[2],即一球懸鈴木(PlatanusoccidentalisL.)、二球懸鈴木(PlatanusacerifoliaWilld.)和三球懸鈴木(PlatanusorientalisL.).其中二球懸鈴木是三球懸鈴木和一球懸鈴木的雜交種,1640年在英國倫敦育成,后引種到世界各大城市,廣泛栽培[3].每年5月份左右,懸鈴木老果脫落導致大量的果毛飄散,而伴隨著新生雄花的開放花粉大量散落,造成環境污染的同時,對人的皮膚和呼吸系統具有一定刺激作用[4-5].近年來,國內外研究者們對懸鈴木的研究多集中在如何解決其花粉散落、落果飛毛的問題上,并通過篩選自然突變體植株[6-7]、化學誘導球果不正常發育促使果實脫落[8]、利用嫁接或組織培養的方式培育少球株系[9-11]、誘變育種[12]、多倍體育種[13]、基因工程育種[14-16]等多種手段進行了系統研究.成俊卿等[17]對懸鈴木木材進行過解剖學研究,但是,有關懸鈴木幼苗的初生維管系統演化結構的研究,鮮見報道.為此,本文對懸鈴木幼苗初生維管組織系統進行了研究,以探索其維管演化進程,為證明子葉節區的原始性及其系統演化提供一定的依據.
1 材料與方法
實驗材料為采自大連地區的普通二球懸鈴木(PlatanusacerifoliaWilld.)種子.室溫萌發,至具有1~3片真葉的幼苗.將幼苗切成6 mm小段,置于于FAA固定液中,采用石蠟切片法制成連續切片,厚度10 μm,番紅、固綠雙重染色,中性樹膠封片.在數碼生物顯微鏡下觀察、照相.
2 實驗結果
二球懸鈴木幼苗為子葉出土幼苗,子葉長條線形.根據子葉節區理論[18],可將幼苗分為子葉節區、上胚軸苗區和下胚軸根區3部分.
2.1 子葉節區
二球懸鈴木子葉節區較長,一般為8.9~13.8 mm,幾乎占據整個幼苗下胚軸的全長.
二球懸鈴木子葉節區上部長2.9~3.2 mm、直徑545~830 μm,至此,子葉節區上部向莖的真中柱過渡的雛形已經基本形成.越靠近苗端時,橫切面上韌皮部彼此距離越明顯,呈間斷性條帶狀分離排列.在橫切面上可見到子葉跡一分為二,形成二葉跡.子葉跡直接與子葉節區上部的管狀中柱相連接,既有子葉跡又有子葉隙.被子葉跡隔開的兩條形的維管束又分成1~3束,且中央部分被薄壁的髓細胞填充,同時子葉跡向外生長(見圖1).

a.二葉跡形成
二球懸鈴木子葉節區中部長4.0~4.3 mm,直徑531~730 μm.位于子葉節區下部的上方.在這個區域,初生木質部出現了中始式管狀中柱階段.中始式二原型管狀中柱的原生木質部位于兩端,從原生木質部處一分為二,同時弦向發育,且中央部分被髓細胞占據,其后生木質部各向外側轉約45°,形成橢圓至方形的中柱類型(見圖2).
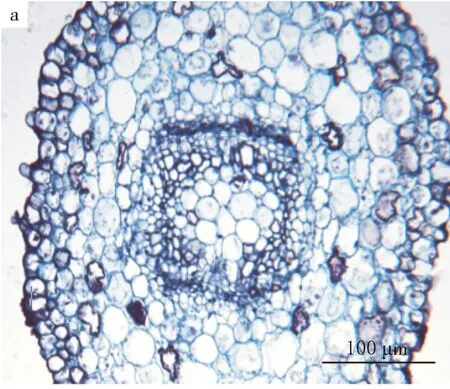
a.子葉節的二葉跡
二球懸鈴木子葉節區長1.6~2.0 mm,其橫切面直徑為430~662μm.中柱類型為中始式二原型單中柱.原生木質部導管直徑很小,由環紋和螺紋加厚的導管構成;后生木質部導管直徑較大,由梯紋、網紋和孔紋導管構成(見圖3—4).
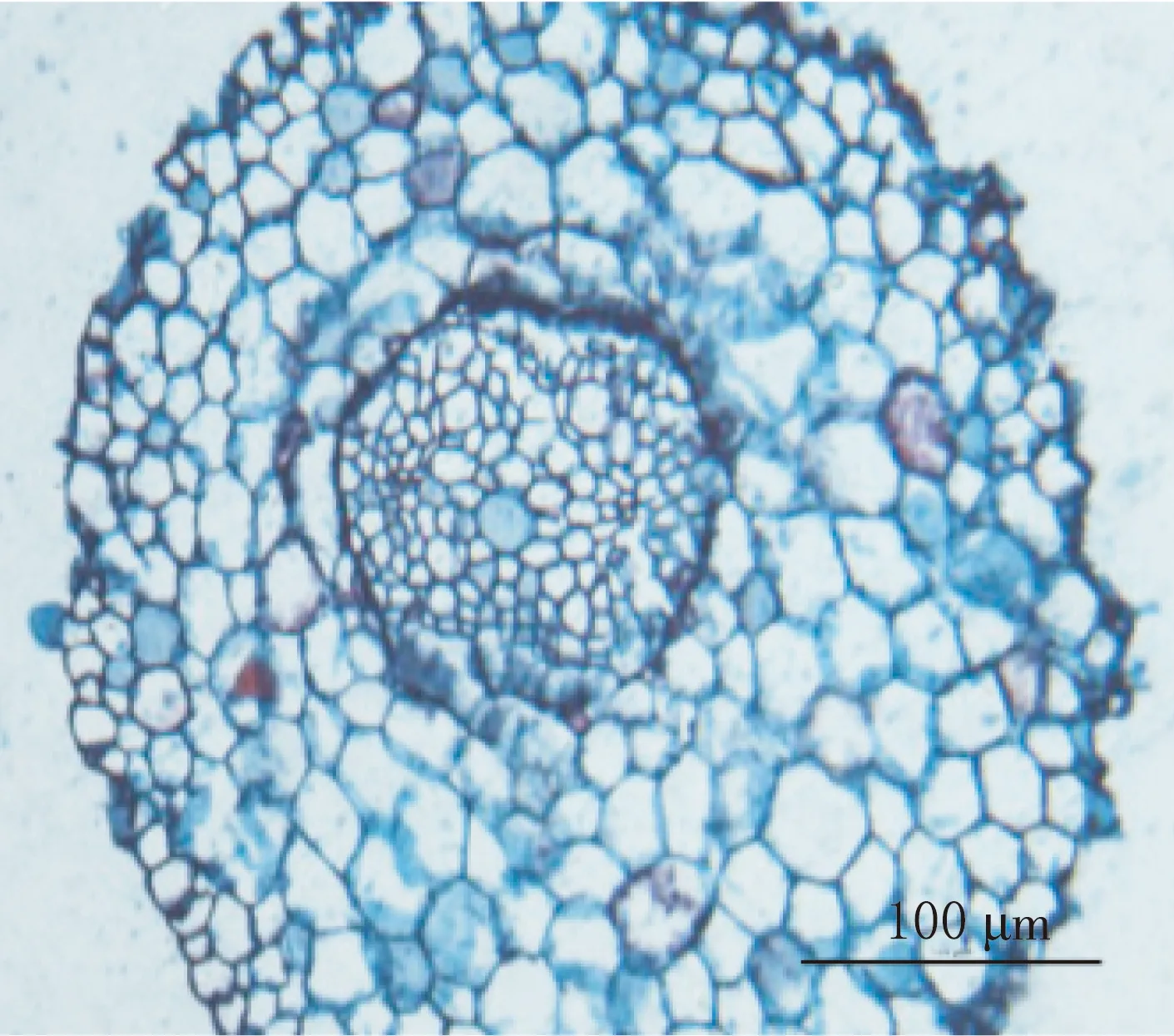
中始式二原型中柱
2.2 上胚軸——苗區
上胚軸苗區屬于植物體的軸向后生體,位于子葉節區的上部.上胚軸下連子葉節區上部,上連頂芽,由子葉節區中、上部分化出來的外韌維管束雛形(見圖1b)逐漸與莖的內始式真中柱連接(見圖5).
莖的內始式中柱由大于6束的外韌維管束組成.子葉雙跡清晰可見(見圖6a).胚芽位于上胚軸上方.由頂芽發生的葉為三葉跡(見圖6b).

a.子葉分化出外韌維管束
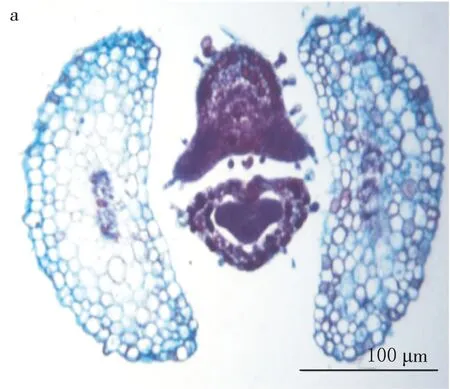
a.子葉的雙葉跡
由生長點和數枚幼葉及腋芽原基等結構,包括:莖的生長點、幼葉原基、幼葉和幼莖的初生結構,表皮、皮層和維管柱見圖7.
2.3 下胚軸—根區
下胚軸根區過渡區位于子葉節區下部的下面,這一階段極短,為0.88~1.08 mm.后生木質部向軸的中央部位旋轉45°,完成了中始式的二原型單中柱向外始式過渡.二球懸鈴木的主根為二原型(或真二原假四原)輻射中柱(見圖8)

莖的生長點、葉原基和幼葉
3 分析與討論
3.1 二球懸鈴木子葉節區初生維管系統演化過程
根據子葉節區理論[18],子葉節區是種子蕨的孑遺部分,故可稱作幼苗的軸向原生體,而分別稱為上胚軸苗區與下胚軸根區的均為其幼苗的軸向后生體部分.從系統發育來看,后兩者均系由前者所衍生.子葉節區相當于一個等二叉狀或復等二叉狀分枝的頂枝束,可分成上、中、下三部分.谷安根等[18]已經對毛茛科、葫蘆科等雙子葉草本植物進行了子葉節區研究,但是對木本植物子葉節區的研究目前尚未報道.
本論文中對二球懸鈴木幼苗的研究結果表明,二球懸鈴木幼苗的初生維管系統具有較長的子葉節區,幾乎占據幼苗下胚軸的全部.通過其子葉橫、縱切面觀察,根端為外始式二原型(或真二原假四原)輻射中柱.子葉節區下部較為原始,具有中始式二原型單中柱,且這一階段特別短;中央出現了具髓的中始式二原型管狀中柱階段,且這一階段較長,髓細胞較大且均由薄壁細胞組成;上部維管束的數目逐漸增多,并且與莖的內始式真中柱極為相似,所以只存在子葉節區莖連接區.上胚軸苗區具有內始式真中柱.由于子葉節區部分中始式管狀中柱的存在,為種子植物與真蕨類可能有過具有管狀中柱階段的共同祖先提供了證據,同時又為幼苗初生維管組織從子葉節區分別向根端和苗端演化提供了依據.由于二球懸鈴木幼苗子葉節區下部出現中始式二原型單中柱及子葉與營養葉不同跡,充分證明了幼苗子葉節區的原始性.二球懸鈴木幼苗的兩子葉跡匯合形成長“V”字形.
3.2 子葉跡與營養葉葉跡
子葉節區的子葉為二葉跡,而胚芽起源的營養葉為三葉跡,由此可判斷營養葉非子葉節區生成.子葉跡直接與子葉節區上部的管狀中柱相連接,既有子葉跡又有子葉隙.
3.3 連接區與過渡區
鑒于子葉節區上部的維管束類型已經與莖的內始式真中柱極為相似并且較短,即子葉節區上部至上胚軸苗區維管束的變化并不是特別激烈,所以特稱為子葉節區-莖連接區.子葉節區下部與根之間雖然距離較短,但是變化明顯,所以特稱為子葉節區-根過渡區.
3.4 擬分生組織
根據子葉節區理論[18],幼苗子葉節區的中、上部分保留有一團與頂端分生組織無關的分生組織性組織(meristematic tissue),即擬分生組織.但是從二球懸鈴木幼苗橫切面上來看,從中部開始形成具有髓的中始式二原型管狀中柱;縱切面上看,中上部的擬分生組織不明顯,這可能與選苗期有關,尚需進一步探究.
3.5 子葉節區類型
子葉節區類型可分為5種類型[18],其中頂枝伸長型的特點為:子葉節區頗長,幾乎占據整個下胚軸的全長.從系統發育來看,它相當于由數次等二叉分枝的復合頂枝(syntelome)所衍生.其子葉柄稍下部的橫切面上,可以觀察到1~3或4至多枚子葉跡(偶數多跡).子葉節區下部和子葉節區-根過渡區甚短,其維管組織的向基過渡亦不甚激烈.子葉節區中部特長,其維管組織向頂(子葉)方向的轉變十分激烈.在具頂枝生長型子葉節區的幼苗,只存在短的子葉節區-根過渡區和子葉節區-莖連接區.
根據二球懸鈴木的初生維管組織特點,其子葉節區類型屬于頂枝伸長型.
國內外,有關被子植物幼苗初生維管系統發育的研究,已經積累了大量的資料,研究重點主要集中在初生維管系統的過渡區上.目前,關于植物初生維管系統根、莖過渡方式主要包括3種主要理論:
伊姆斯和麥克唐納(Eames and MacDaniels,1947)理論,這一學派認為:根和莖是互相連續的結構,共同組成植物體的體軸.在植物幼苗時期的莖和根相接的部分,出現雙方各自特征性結構,即根的初生維管組織為間隔排列,木質部為外始式;莖的初生維管組織為內外排列.木質部為內始式的過渡,即:根-莖過渡區.過渡區通常很短,從小于1 mm到2~3 mm,很少達到幾厘米.過渡區發生在胚根以上的下胚軸的最基部、中部或上部,終止于子葉節上.在過渡區內,表皮、皮層等是直接連續的,但維管組織要有一個改組和連接的過程.
Katherine Esau理論(1950),主張幼苗可分成根-下胚軸-子葉與上胚軸苗等兩個單位, 上胚軸常似一種分離的結構,生在根-下胚軸-子葉的單位上,根只與子葉之間存在維管組織的轉換,而與上胚軸(莖)間不存在過渡.就是說下胚軸是根與子葉之間的過渡區,并非是“根-莖過渡區”.過渡區代表一種連接,這并不能認為是介乎組織上有不同排列的兩種器官之間的連接,而應看作是維管系統與葉子發育有關系的連接.因此,過渡區的研究必須說明根的維管系統和最早葉跡之間的關系.并以甜菜幼苗為例,說明雙子葉植物根和子葉連接的過渡區.
子葉節區(Cotyledon node zone,1990)理論,谷安根等對毛茛科某些屬植物幼苗初生維管系統的發育方面做了大量研究,并探討了毛茛科不同屬間的中柱演化關系,在此基礎上,創建了子葉節區理論,并就其在被子植物演化形態學上的應用進行了闡述.根據子葉節區理論,將雙子葉植物幼苗形態分為上胚軸苗區、子葉節區和下胚軸根區.其中的植物子葉節區是種子蕨的孑遺部分,故可稱作幼苗的軸向原生體,而分別稱其上胚軸苗區與下胚軸根區均為其幼苗的軸向后生體部分.從系統發育來看,后兩者均由前者所衍生.本文驗證了谷安根的子葉節區理論.
