池塘專養模式下中華鱉養成品組織和臟器質量對體質量的影響效果分析
2021-03-31 08:26:04牟恩鏇申屠琰葉文婷李有志朱衛東王志錚
浙江海洋大學學報(自然科學版) 2021年6期
牟恩鏇,申屠琰,葉文婷,李有志,朱衛東,王志錚
(1.浙江海洋大學水產學院,浙江舟山 316022;2.余姚市水產技術推廣中心,浙江余姚 315400)
動物的組織和臟器按其生理代謝功能可分為活性和惰性兩部分。“器官代謝活性假說”指出,動物組織和臟器質量與體質量間呈異速生長,心、肝、腦、鰓等活性部位和白肌、脂肪塊等惰性部位與體質量間的比例隨個體生長分別呈下降和上升的趨勢[1-2]。與之相對應,動物在生長過程中所表露的活性臟器質量逐漸增大而其臟器系數逐漸減小的特征[3-5],既反映了活性部位因代謝損耗而喪失了與體質量同步增長的可能性,也揭示了惰性部位通過不斷積聚物質和能量具備了超越體質量增長的增重速度。無疑,大西洋黃貂魚Dasyatis sabina 和圓口銅魚Coreius guichenoti 活性臟器的臟器系數與其體質量均呈負相關的情形[6-7],也進一步印證了活性臟器在動物體質量增長過程中因發揮其正常生理代謝功能而產生大量能耗的事實,即動物體質量增長既依賴活性臟器代謝能耗的持續增加,更表露為惰性部位對代謝產物的不斷積累。因此,借助相關分析、通徑分析、復相關分析和偏回歸分析等方法開展動物組織和臟器質量對體質量影響效果的定量研究,深入剖析機體各組織和臟器質量與體質量間的真實關系,無疑對于揭示其體質量增重機制具重要現實意義。但,迄今國內外基于形態表型的水產動物體質量增長對策研究,僅涉及形態體尺對體質量影響效應的報道[8-13]。鑒于此,本研究團隊以池塘專養模式下25 月齡中華鱉Trionyx sinensis 余姚本地品系同生群雄性個體為研究對象,在較系統開展形態體尺性狀對體質量影響效果的基礎上[13],再次就實驗鱉組織和臟器質量對其體質量的影響效應開展研究,以期從體尺性狀與組織和臟器質量性狀兩個維度,綜合分析實驗鱉的體質量增長機制,以期為以進一步豐富余姚本地中華鱉速生品系的精準選育提供科學依據。
1 材料與方法
1.1 實驗鱉
本研究實驗鱉的來源及前處理完全同文獻[13]。
1.2 性狀測定
自然解凍-20 ℃下活體凍藏24 h 的實驗鱉,用紗布擦凈體表并稱量其體質量;按常規解剖方法,逐尾剪取裙邊、剝離背甲和腹甲、摘取心、肝、腸并去除附于其上的脂肪和結締組織,依次用濾紙吸干表面水分后稱量。其中,體質量的稱量工具為HZ1201A 電子天平(慈溪紅鉆衡器設備有限公司出品,精度0.1 g),各組織和臟器質量的稱量工具均為BSA223S 電子天平(賽多利斯科學儀器有限公司出品,精度1 mg)。
1.3 數據處理
整理所測數據,分別計算各項稱量性狀的均值、標準差和變異系數,并采用多重比較法,比較各組織、臟器質量與體質量比值間的差異顯著性(視P<0.05 為顯著水平);參照文獻[13]的方法,運用SPSS 23.0 軟件依次開展各項稱量性狀間的簡單相關性分析以及組織和臟器質量性狀對體質量的通徑分析、復相關分析和偏回歸分析,確定影響體質量的關鍵性狀組合,并對所建估算體質量的多元線性回歸方程進行擬合度檢驗。
2 結果
2.1 參數統計
由表1 可知,本研究所測性狀的變異系數均波動于10%~20%之間,呈W6>W4>W5>BW>W3>W2>W1;由表2 可見,各組織、臟器與BW 的比值分別呈W4>W5>W6>W2>W1>W3(P<0.05)。即,以裙邊和體甲為代表的機體惰性組織質量的變異程度和對體質量的貢獻率,均明顯大于以肝、腸和心為代表的機體代謝活性臟器,表明機體代謝惰性部位的質量變異程度和對體質量的貢獻率均明顯高于其代謝活性臟器。

表1 實驗鱉各測定性狀的統計量描述(n=30)Tab.1 Description of statistics for measurement indicators

表2 實驗鱉組織、臟器質量與體質量的百分比(%)Tab.2 Percentage of tissue,viscera and body weight of T.sinensis(%)
2.2 相關分析
由表3 可見,BW 與本研究所涉各性狀間的相關系數均達到極顯著相關水平(P<0.01),且呈W4>W2>W1>W5>W3>W6,即與體質量相關性最強的為裙邊,最弱的為腹甲,結合W1、W2、W3與W4和W5間的相關系數均達到顯著水平(P<0.05),而W4與W5、W6間的相關系數均未達到顯著差異(P>0.05)的情形,既表明裙邊、背甲與各活性臟器間,以及各活性臟器之間在質量上均存在強相關性,而裙邊和背甲之間在質量上卻僅具弱相關性,也反映了裙邊質量較其它組織和臟器對體質量更具增重重要性。
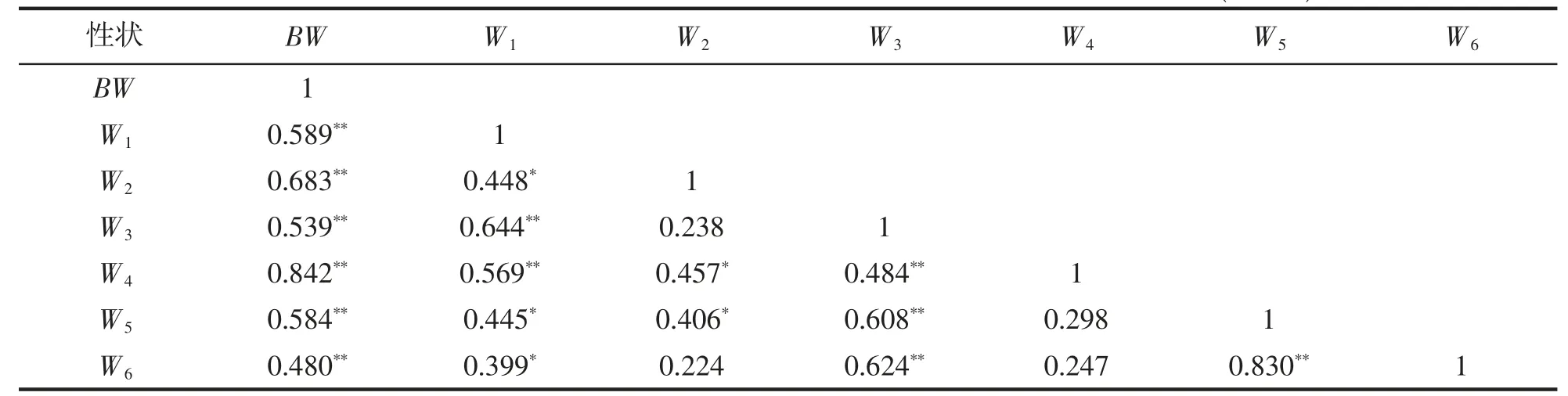
表3 實驗鱉各測定性狀間的相關系數(df=28)Tab.3 Correlation coefficients of measured characters of T.sinensis (df=28)
建立實驗鱉組織和臟器質量與體質量間的一元線性回歸方程得表4。由表4 可見,本研究所涉各性狀與BW 的回歸系數均達到極顯著水平(P<0.01)。從一元線性回歸方程的斜率呈W4>W5>W6>W2>W1>W3看,以裙邊和體甲為代表的機體惰性組織質量對體質量增速的貢獻率明顯大于以肝、腸、心為代表的機體活性臟器,尤以裙邊質量為甚。
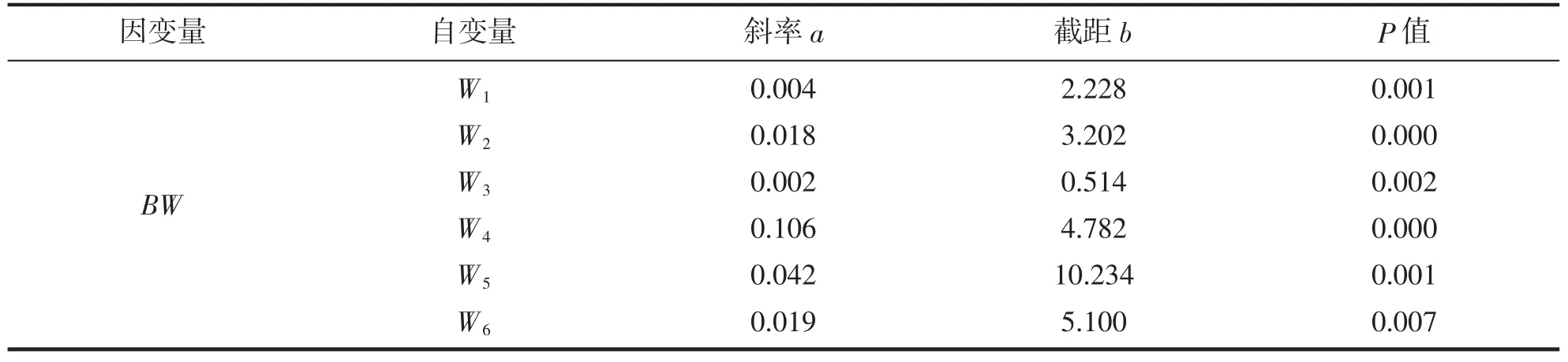
表4 實驗鱉組織和臟器質量與體質量間的一元線性回歸分析Tab.4 Unitary linear regression analysis of tissue and viscera weight and body weight of T.sinensis
2.3 通徑分析
由表5 可見,被保留性狀與BW 間的通徑系數均達到極顯著水平(P<0.01),方差膨脹因子也均遠小于經驗值(VIF<10),它們對BW 的直接作用和間接作用分別為W4>W5>W2和W2>W5>W4,其中直接作用大于間接作用的僅為W4,且W4對BW 的間接作用主要通過W2來實現,W5對BW 的間接作用主要通過W4來實現。由此可知,W4為影響BW 的關鍵核心變量,W2和W5則為影響BW 的相對重要變量。
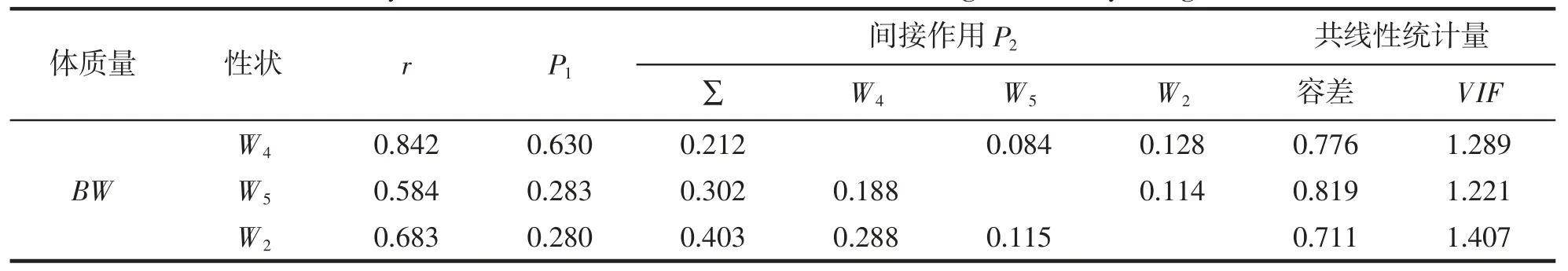
表5 實驗鱉組織和臟器質量對體質量影響的通徑分析Tab.5 Path analysis of influence of tissue and viscera weight on body weight of T.sinensis
2.4 決定系數分析與復相關系數分析
由表6 可見,單一性狀對BW 的決定系數呈W4>W5>W2,性狀兩兩交互對BW 的決定系數呈W2-W4>W4-W5>W2-W5;經計算,W2-W4-W5組合對BW 的總決定系數為0.887。由表7 可見,W2-W4-W5組合與BW間復相關系數達到極顯著水平(P<0.01),所示復相關指數亦為0.887。無疑,上述結果進一步印證了表4 所示W4為影響BW 的關鍵核心變量,以及W2和W5為影響BW 的相對重要變量的可靠性。
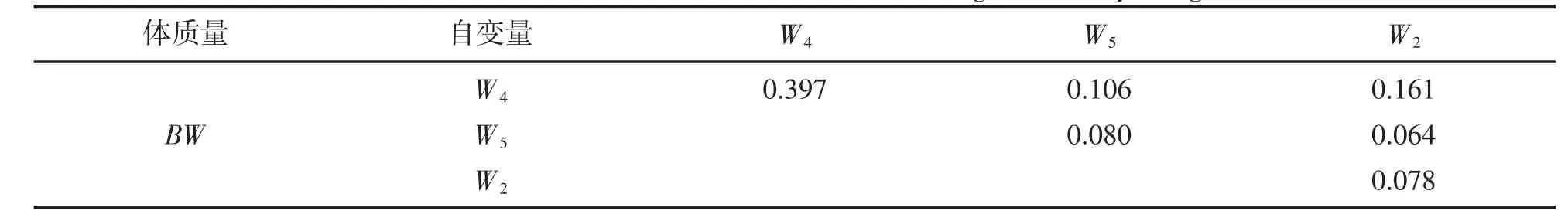
表6 實驗鱉組織和臟器質量對體質量的決定系數Tab.6 Determination coefficients of tissue and viscera weight on body weight of T.sinensis

表7 實驗鱉組織和臟器質量與體質量的復相關系數Tab.7 Complex correlation coefficients between tissue and viscera weight and body weight of experimental T.sinensis
2.5 多元回歸方程構建與檢驗
由表8 可見,所有被保留性狀的偏回歸系數均達到極顯著水平(P<0.01),用于估算BW 的線性回歸方程為BW=10.73W2+5.02W4+3.94W5-57.45;方差分析顯示,該方程的回歸系數亦達到極顯著水平(P<0.01)(表9);經回歸預測,估算值與實測值間無顯著差異(P>0.05),表明該方程能夠精確反映實驗鱉組織和臟器質量與體質量間的真實關系。
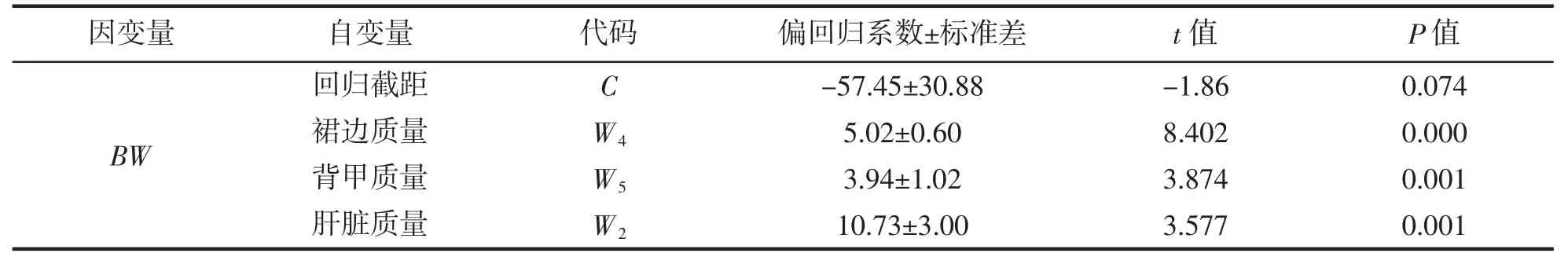
表8 實驗鱉組織和臟器質量與體質量的偏回歸分析Tab.8 Partial regression analysis of tissue and viscera weight and body weight of T.sinensis

表9 實驗鱉組織和臟器質量與體質量的多元回歸方程的方差分析Tab.9 Analysis of variance of multiple regression equation of tissue and viscera weight and body weight of T.sinensis
3 討論
3.1 W2-W4-W5 成為影響BW 關鍵性狀組合的原因分析
由表6 和表7 可知,W2-W4-W5對BW 的總決定系數和復相關指數均為0.887,均大于劉小林等[8-9]所認為的影響因變量的主要自變量組合已被找到的閾值0.85,表明W2-W4-W5確實為影響BW 的關鍵性狀組合。
研究表明,蛋白質是生物機體代謝的最終能源物質[14-16]。無疑,就生物有機體而言,蛋白質較脂肪、糖類等其它代謝能源利用物質不僅更具儲藏價值,而且對體質量的穩定增長更具生理重要性。Ca 既是構成機體骨骼的基礎性元素,也是機體生長不可或缺的大量元素。據報道,中華鱉裙邊富含具生理活性的膠原蛋白[17],其蛋白質含量遠高于肌肉,而脂肪則明顯低于肌肉[18];背甲中的礦物元素含量以Ca 為最高,達22 961.30 mg·100-1·g-1,遠高于肌肉中的75.70 mg·100-1·g-1[19]。由此,基于本研究中裙體比和甲體比(背甲)均顯著高于其它組織和臟器(P<0.05)的情形(表2),以及W1、W2、W3間均呈強相關,而W4與W5和W6則均僅具弱相關(表3)的結果,可將裙邊和背甲分別認定為中華鱉機體生理活性蛋白與Ca 元素的儲藏庫,按“器官代謝活性假說”[1-2],兩者分別為儲藏生理活性蛋白和Ca 元素的惰性組織。肝臟與機體蛋白質、脂肪和碳水化合物等基本能源物質的分解與合成,組織和臟器所需氨基酸的合成、搭配與輸送等環節,以及生成可促進鈣元素吸收、代謝、轉運及骨礦化等過程均有著極為密切的關聯[20-21],是中華鱉的能量代謝中樞和機體內最重要的活性器官。故,結合W4對BW 的間接作用主要通過W2來實現和W5對BW 的間接作用主要通過W4來實現(表5)的結果,可認定肝臟為支持裙邊生理活性蛋白和背甲Ca 元素儲藏的重要轉移庫。按“器官代謝活性假說”[1-2],肝臟為與生理活性蛋白和Ca 代謝密切關聯的活性臟器。
盡管本研究中W1、W2、W3與BW 均呈極顯著正相關(P<0.01)的結果(表3),與黃姑魚Nibea albiflora和羅非魚Oreochromis niloticus 活性臟器的質量與其體質量間均呈顯著正相關的情形完全相符[22-23],卻無法動搖W4和W5對BW 的直接作用和單一決定系數均大于W2(表5、表6)的事實,表明實驗鱉體質量的增長更倚重裙邊和背甲等惰性部位的物質積累,這與前述“器官代謝活性假說”及其它相關文獻所反映的動物體質量增長既依賴活性臟器代謝能耗的持續增加,更表露為惰性部位對代謝產物的不斷積累的觀點相符[1-7]。結合本研究BW 與W4間的相關系數和一元線性回歸方程的斜率分別為BW 與W5間的1.44 倍(表3)和2.52倍(表4),以及W2與W4間的相關系數為W2與W5間的1.28 倍(表3)的結果,本文作者認為W4為影響BW的關鍵核心變量,而W5僅為影響BW 的相對重要變量的原因,一方面應與蛋白質分子量明顯大于鈣元素有關,另一方面則可能與生理活性蛋白產物的外溢效應明顯強于鈣質有關。“器官代謝活性假說”認為,活性器官質量與體質量的比例與其代謝水平呈正相關[1-2]。由此,W1和W3未能入選影響BW 關鍵性狀組合的原因,因其與體質量的比例顯著低于W2有關(P<0.05) (表2)。
3.2 實驗鱉基于形態構造的體質量增長對策分析
牟恩鏇等[13]以本研究實驗鱉為研究對象,采用通徑分析方法找到了影響實驗鱉BW 的關鍵體尺性狀組合為“頸長—后肢長—體寬”,并從形態與所在區域臟器和組織功能相維系的視角,將該關鍵性狀組合衍生為“搶食功能—運動功能—消化功能”,由此提出了在池塘專養條件下捕食運動能力較臟器消化能力在影響實驗鱉BW 上更具重要性的觀點。
一般而言,在相同生境下惰性部位的質量大小可作為表征機體體質強弱的顯性指標。故,按此可將本研究所示影響實驗鱉BW 的關鍵組織和臟器質量組合中的W4(裙邊質量)和W5(背甲質量)表征為體質強度,W2(肝質量)表征為代謝強度。頭足類肉鰭具輔助運動及在游泳中保持平衡的作用[24],實驗鱉裙邊所處部位和功能與其類似,故亦可將W4(裙邊質量)表征為攝食運動輔助強度。由此,體尺性狀關鍵組合“頸長—后肢長—體寬”和組織與臟器質量性狀關鍵組合“裙邊質量—背甲質量—肝質量”在影響BW 的作用機制上獲得了互證,即實驗鱉基于其形態構造形成了強化攝食運動、提高代謝水平和提升機體體質三維有機互動的體質量增長對策。其中,強化攝食運動強度對提高活性部位的代謝水平具牽引作用,提升機體體質對強化攝食運動強度具促進作用,提高活性部位的代謝水平對提升機體體質具支持作用。
4 結語
4.1 以裙邊和體甲為代表的機體惰性組織質量對體質量及其增速的貢獻率,均明顯大于以肝、腸和心為代表的機體活性臟器質量,尤以裙邊質量為甚。
4.2 實驗鱉裙邊與體甲和各活性臟器間,以及各活性臟器之間在質量上均具強相關性,而裙邊與背甲之間在質量上則僅具弱相關性。其中,裙邊和背甲分別為儲藏生理活性蛋白和Ca 元素的惰性組織,肝臟為支持裙邊生理活性蛋白和背甲Ca 元素儲藏的活性臟器。
4.3 裙邊質量為影響體質量的關鍵核心變量,肝質量和背甲質量為影響體質量的相對重要變量,它們對體質量的總決定系數和復相關指數均為0.887。三者與體質量的線性多元回歸方程為BW=10.73W2+5.02W4+3.94W5-57.45。
4.4 實驗鱉基于其形態構造形成了強化攝食運動、提高代謝水平和提升機體體質三維有機互動的體質量增長對策。其中,強化攝食運動強度對提高活性部位的代謝水平具牽引作用,提升機體體質對強化攝食運動強度具促進作用,提高活性部位的代謝水平對提升機體體質具支持作用。
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
中國生殖健康(2019年2期)2019-08-23 08:12:08
產品可靠性報告(2017年7期)2017-09-05 09:49:12
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
汽車觀察(2016年3期)2016-02-28 13:16:26
