IPA 模式對鯉生長、肌肉和血漿中游離氨基酸含量的影響
2021-03-31 08:26:06李曉東胡建勇
浙江海洋大學學報(自然科學版) 2021年6期
李曉東,胡建勇,高 攀,李 林,劉 晶
(新疆維吾爾自治區水產科學研究所,農業部西北地區漁業資源環境科學觀測實驗站,新疆烏魯木齊 830000)
新疆地處我國西北地區,水資源時空分配不均,水產養殖受水資源的制約和用水政策影響較大,尤其是池塘養殖業受到的不利影響更為深遠;提升漁業養殖用水效率,實施高效節水養殖已成為新疆池塘養殖業從業者所面臨的難題。近年來,本地區開展了以節水、高效為重點的設施化池塘養殖技術的試驗,相比傳統池塘養殖,集約化池塘內循環流水養殖系統(intensive pond aquaculture,IPA)因水資源利用效率高、環境友好備受行業從業者關注。而養殖效益作為衡量水產養殖業的主要指標,受技術、管理等影響較大,研究IPA 模式水流刺激對養殖魚類生理生化的影響,對于揭示養殖品種的能量利用及提升經濟效益具有重要意義。現有證據表明西伯利亞鱘Acipenser baerii 和俄羅斯鱘A.gueldenstaedti 在循環水養殖條件下均可快速生長,但西伯利亞鱘具有更強的抗氧化能力和消化能力[1];而在池塘養殖模式下,大口黑鱸Micropterus salmoides 等肉營養和肉質的表現比循環水養殖的差[2]。
蛋白質在魚類營養中具有重要作用,而氨基酸作為蛋白質的基本組成成分,其在調控魚類生理活動和行為習性中起關鍵作用,其含量亦受攝食及魚體自身蛋白質代謝的影響。血液循環系統通過紅細胞與血漿的相互作用在氨基酸轉運中起關鍵作用,其與多種組織細胞內氨基酸含量有密切關系,而肌肉作為魚體的主要組織,是游離氨基酸的主要儲存場所,尤其以白色肌肉更為甚之[3-4]。鑒于此,本研究以鯉C.carpio 為對象,通過基于IPA 原理的浮式池塘內循環系統和普通池塘養殖試驗,比較不同養殖模式下鯉魚肌肉和血漿中游離氨基酸變化情況,探討IPA 模式水流對魚類的影響,在理論上豐富了魚類運動與營養代謝方面的內容,實踐上可為今后改善養殖條件、提高養殖魚類的生長速度、完善品質和營養價值提供科學依據。
1 材料與方法
1.1 試驗條件
試驗地點位于新疆水生野生動物救護中心。以普通標準化的土池塘作為對照塘,各2 口,水面面積均為0.3 hm2;以IPA 浮式養殖系統為試驗塘,由2 口面積均為0.3 hm2的池塘(A 池、B 池)組成,如圖1 所示,構建浮式養殖平臺系統(以下簡稱“平臺系統”)用于養殖試驗,其所需水源從B 池抽取,排放尾水依次進入A 池、B 池;在A、B 池中投放一定數量的鰱鳙等濾食性水生動物;設置植物浮床種植水生植物,實現水體凈化,經過一定的流程后實現養殖水體循環利用。
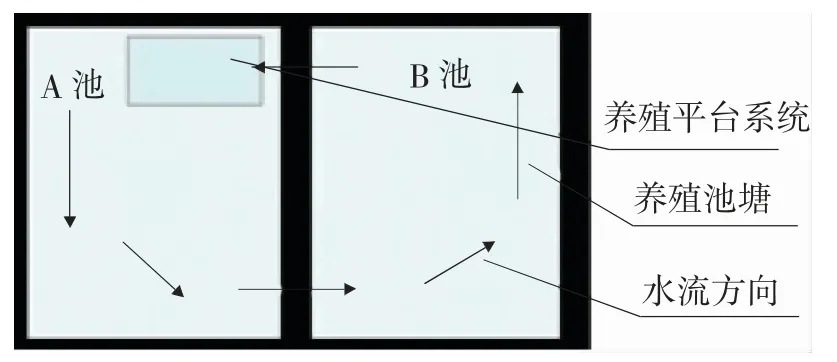
圖1 基于IPA 原理的養殖系統Fig.1 Breeding system based on IPA principle
平臺系統操作平臺包括浮體單元結構和養殖單元結構。浮體單元框架由40 mm×40 mm×3 mm 的鍍鋅角鐵焊接而成,并鋪設格柵板,浮體為厚度25 cm 的泡沫板。平臺系統養殖池為自制帆布池,直徑3 m,圓柱體墻高1 m,底部圓錐體高0.5 m,在圓錐體最低處設置直徑為60 mm 的排污口。在帆布池圓柱體與圓錐體連接處鋪設微孔曝氣管,通過羅茨風機供氣持續增氧,使池內水體形成由四周向中心的渦旋狀態,以便將殘餌和糞便集中到底部圓錐排污口,再通過排污管道到達集污池,沉淀過濾后,尾水進入池塘,通過池塘內凈化后再次進入帆布池,形成循環。
1.2 試驗魚放養及管理
易捕鯉C.carpio var.Yibu 為從中國水產科學研究院黑龍江水產研究所引進的水花苗種培育而成,建鯉C.carpio var.Jian 為親本自繁水花培育而成。對照池放養模式分別為:主養易捕鯉(1.5×104尾·hm-2)、套養鰱鳙(鰱:1.5×103尾·hm-2,鳙:7.5×102尾·hm-2);主養建鯉(1.5×104尾·hm-2)、套養鰱鳙(鰱:1.5×103尾·hm-2,鳙:7.5×102尾·hm-2)。平臺系統試驗池分別單養易捕鯉、建鯉,每種魚使用3 個養殖池,放養密度均為10 kg·m-3水體,即每個養殖池投放60 kg,外塘放養適量鰱鳙魚,利用植物浮床種植水蕹菜等水生植物。養殖時間56 d,養殖期間,投喂浮性顆粒飼料,飼料來源于成都海大生物科技有限公司生產的魚種專用料。其營養成分如表1 所示。
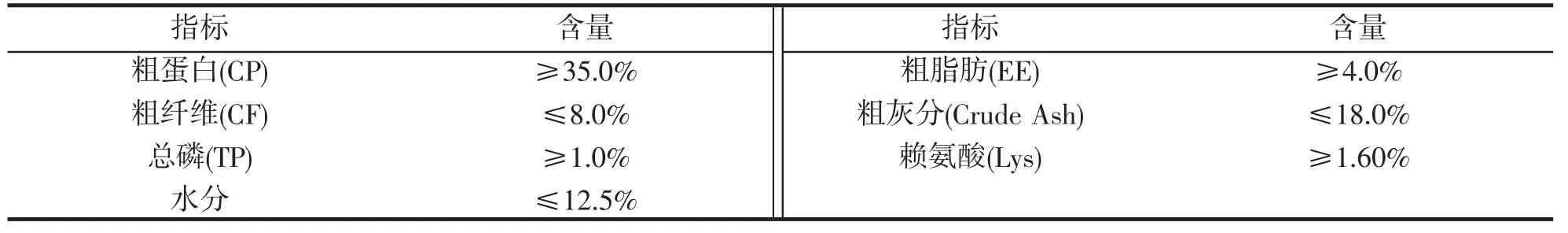
表1 飼料主要營養成分Tab.1 Main nutritional components of feed
養殖期間采用飽食投喂。養殖平臺系統保持持續進水和增氧狀態,增氧的強度以使養殖池中的糞便等顆粒物向魚池中心聚集為準,以便其及時通過底部排污口排出;養殖平臺外塘的增氧設施為微孔曝氣增氧系統,每天根據安裝的物聯網系統設置的水中溶氧情況,適時自動增氧。養殖期間,水溫維持在23~28 ℃,pH 7.3~7.8,溶氧≥5.8 mg·L-1,氨氮≤0.1 mg·L-1。
1.3 樣品采集與指標測定
1.3.1 樣品采集
試驗結束,饑餓24 h,分別從對照池塘隨機選取15 尾魚,作為對照組樣本,然后從平臺系統每個養殖池隨機選取5 尾試驗魚,分別混合后獲得易捕鯉和建鯉的試驗組樣本,測量其體重、體長、全長等指標,有關計算公式如下:

式中:WGR 為增重率,%;SR 為成活率,%;SGR 為特定生長率,%;FER 為飼料轉化效率,%;Wi為試驗開始時魚體重,g;Wf為試驗結束時魚體重,g;t 為養殖試驗天數,d;F 為攝食量,g;P 為粗蛋白含量,%;Wt為魚體重量,g;Wh為肝臟質量,g;Wb為內臟質量,g;We為腸系膜脂肪重量,g。
分別從每個養殖池隨機選取3 尾魚,利用浸潤肝素鈉(5 mL)采血管尾靜脈采血法抽取血液1 mL,置于潔凈的1.5 mL 離心管中,以常溫4 000 r·min-1離心8 min,取上層血漿于-80 ℃冰箱保存備用。在采血結束后,分別取下魚體背部兩側白色肌肉,攪碎混勻后取混合樣品50 g,在-80 ℃條件下作短暫保存。
1.3.2 游離氨基酸含量測定
委托上海百趣生物醫學科技有限公司進行血漿和肌肉中游離氨基酸含量的測定。稱取樣品于EP 管中,加入1 000 μL 提取液(體積比,乙腈-甲醇-水,2:2:1,-20 ℃),渦旋30 s 混勻;加入鋼珠,45 Hz 研磨處理4 min,冰水浴條件下超聲5 min,重復此步驟3 次;-20 ℃靜置1 h;取出樣品4 ℃,12 000 r·min-1離心15 min;取80 μL 上清液至LC 進樣瓶中,用于LC-MS/MS 分析。另取上清液分別稀釋10 倍、100 倍后用于LC-MS/MS 分析。
1.4 數據處理與分析
實驗數據用Excel 2010 和SPSS 24.0 軟件進行統計分析,顯著性方差分析法采用單因素方差分析進行顯著性檢驗。
2 結果與分析
2.1 不同養殖模式對2 種鯉生長及餌料利用的影響
如表2,與對照組比較,IPA 模式下易捕鯉、建鯉體增重均不同程度增加(P<0.05)。其中,易捕鯉提增重率達到了110.57%,建鯉增重率達到了92.98%。在特定生長率方面,試驗組均顯著高于對照組(P<0.05),且IPA 模式下易捕鯉較建鯉高13.73%(P<0.05);各組養殖成活率均達到97%以上;從飼料轉化效率看,對照組均高于試驗組(P<0.05),而2 種魚在同一模式下,差異不明顯。
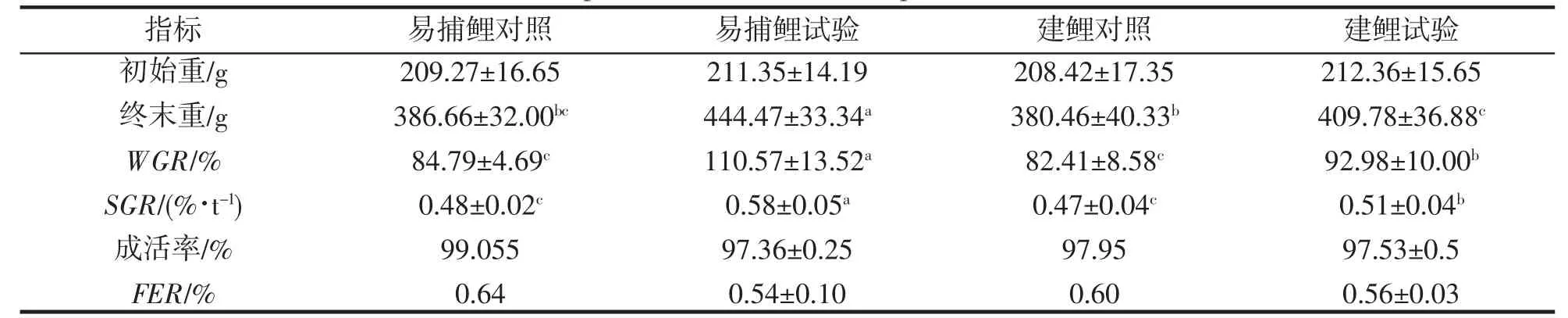
表2 不同養殖模式對2 種鯉生長與攝食的影響Tab.2 Effects of different culture modes on growth and feeding of two species of C.carpio var.Yibu and C.carpio var.Jian
2.2 不同養殖模式下,2 種鯉魚白色肌肉中游離氨基酸含量變化
本試驗共檢測了色氨酸(Trp)、苯丙氨酸(Phe)、纈氨酸(Val)、甲硫氨酸(Met)、異亮氨酸(Ile)、精氨酸(Arg)、組氨酸(His)、亮氨酸(Leu)、蘇氨酸(Thr)、賴氨酸(Lys)、酪氨酸(Tyr)、脯氨酸(Pro)、甘氨酸(Gly)、谷氨酸(Glu)、天冬氨酸(Asp)、天冬酰胺(Asn)、丙氨酸(Ala)、谷氨酰胺(Gln)、絲氨酸(Ser)等19 種游離氨基酸。由圖2 可知,建鯉白色肌肉游離氨基酸中蘇氨酸、甘氨酸、絲氨酸、賴氨酸、天冬氨酸、天冬酰胺在試驗組中含量與對照組中含量差異顯著(P<0.05),其中前3 種呈顯著降低,后3 種呈顯著增加;而易捕鯉肌肉游離氨基酸中僅賴氨酸和谷氨酸在試驗組中含量與對照組中含量差異顯著(P<0.05),其中賴氨酸呈現顯著增加,谷氨酸呈現顯著降低。通過計算肌肉中的總游離氨基酸(total free amino acid,TFAA)和游離氨基酸中必需氨基酸含量(essential amino acid,EAA)、支鏈氨基酸(branched-chain amino acids,BCAA)、芳香族氨基酸(aromatic amino acid,AAA)含量,2 種鯉魚TFAA、EAA、BCAA、AAA 均表現出差異不顯著(圖3)。

圖2 池塘與IPA 養殖模式下2 種鯉魚肌肉中游離氨基酸含量Fig.2 Contents of free amino acids in white muscle of C.carpio var.Yibu and C.carpio var.Jian under pond and IPA culture mode
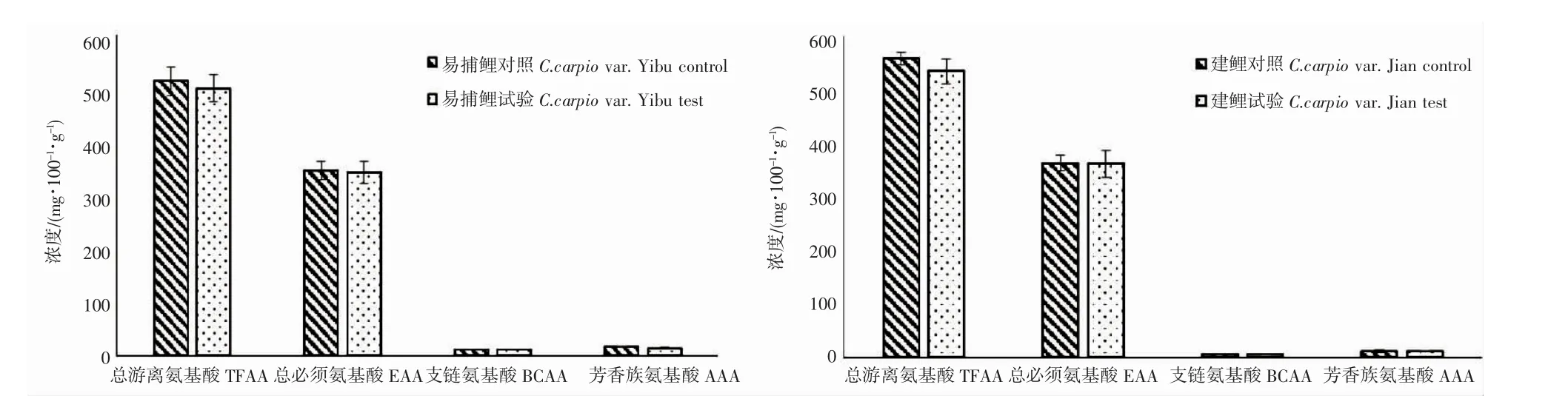
圖3 池塘與IPA 養殖模式下2 種鯉魚白色肌肉中TFAA、EAA、BCAA、AAA 含量Fig.3 Contents of TFAA、EAA、BCAA、AAA in white muscle of C.carpio var.Yibu and C.carpio var.Jian under pond and IPA culture mode
2.3 不同養殖模式下,2 種鯉魚血漿中游離氨基酸含量變化
本試驗在2 種鯉魚血漿檢測的前述19 種游離氨基酸中,建鯉僅有丙氨酸差異不顯著,其他18 種氨基酸試驗組含量較對照組含量呈現顯著增加(P<0.05);而易捕鯉中除天冬氨酸、天冬酰胺、丙氨酸外其他16種氨基酸試驗組含量較對照組含量呈現顯著降低(P<0.05)(圖4)。通過計算肌肉中的總游離氨基酸(total free amino acid,TFAA)和游離氨基酸中必需氨基酸含量(essential amino acid,EAA)、支鏈氨基酸(branched-chain amino acids,BCAA)、芳香族氨基酸(aromatic amino acid,AAA)含量,2 種鯉魚TFAA、EAA、BCAA、AAA 均表現出差異顯著(P<0.05),其中建鯉呈顯著增加而易捕鯉呈顯著降低(圖5)。
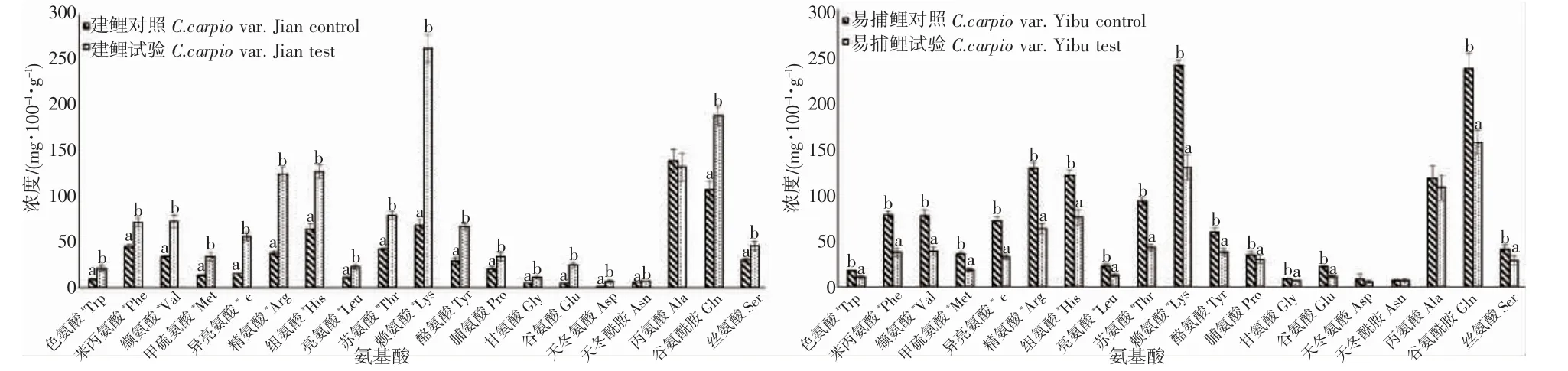
圖4 池塘與IPA 養殖模式下2 種鯉魚血漿中游離氨基酸含量Fig.4 Contents of free amino acids in plasma of C.carpio var.Yibu and C.carpio var.Jian under pond and IPA culture mode
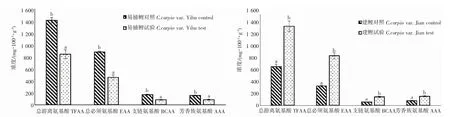
圖5 池塘與IPA 養殖模式下2 種鯉魚血漿中游離氨基酸含量Fig.5 Contents of TFAA、EAA、BCAA、AAA in plasma of C.carpio var.Yibu and C.carpio var.Jian under pond and IPA culture mode
3 討論
3.1 養殖模式對2 種鯉生長的影響
魚類通過攝食獲取的蛋白質消化為游離氨基酸后通過腸道吸收經血液循環系統運輸至各個器官用于多種組織蛋白合成、糖異生、脂類合成及參與能量代謝[4-6]。但對于養殖而言,如何使攝食蛋白最大化的用于魚類生長進而提升養殖效益是生產者所關注的焦點,而魚體肌肉蛋白作為魚體肌肉的主要組成部分其與評價魚類經濟價值的重要指標含肉率及營養成分密切相關,受養殖模式、養殖品種影響[7-9]。
養殖環境或條件能顯著影響魚類的生長性能,在集約化水產養殖生產中,由于養殖密度增加,帶來養殖對象擁擠脅迫,從而影響生長[10-11]。本試驗結果顯示,IPA 模式下,易捕鯉和建鯉的生長性能均顯著高于池塘養殖模式,這與大口黑鱸M.salmoides 等[12-13]魚類研究結果一致;而與黃顙魚(Pelteobagrus fulvidraco)等[10]魚類研究結果相反;另外,本研究中,無論IPA 模式還是池塘模式,易捕鯉生長性能均不同程度高于建鯉,這可能與品種自身特性有關。
3.2 養殖模式對兩種鯉肌肉游離氨基酸含量的影響
游離氨基酸作為合成肌肉蛋白的條件因子,同樣受品種、養殖模式的影響,筆者發現,在IPA 和靜水池塘養殖模式下,2 種鯉魚肌肉游離氨基酸總含量均差異不顯著,這與Piaractu mesopotamicus[14]在較低流速下(流速等于體長)所獲得的結果一致,但該魚在高流速情況下(流速等于2 倍體長)肌肉中游離氨基酸總含量比對照組顯著增加,這從另一方面說明魚類肌肉中的游離氨基酸含量與魚類生存環境密切相關,這在異育銀鯽Carassius auratus gibelio[15]的研究中有類似的結果。就魚類肌肉中的不同種類游離氨基酸含量而言,亦受養殖模式、品種影響,羅非魚Oreochromis mossambicus 在池塘和水庫養殖模式下,前者肌肉游離氨基酸中天冬氨酸、絲氨酸、谷氨酸、甘氨酸、丙氨酸含量顯著高于后者[16];草魚Ctenopharyngodon idella 經投喂蠶豆脆化養殖后肌肉游離氨基酸中的丙氨酸、異亮氨酸、賴氨酸含量高于普通草魚但甘氨酸、亮氨酸含量低于普通草魚[17];同在池塘養殖模式下,南方大口鯰Silurus soldatovi meridionalis 和鯰魚S.asotus 肌肉中的游離氨基酸含量具有差異性,前者蘇氨酸、甘氨酸、丙氨酸含量高于后者而異亮氨酸、亮氨酸低于后者[18]。同樣,筆者發現在2 種養殖模式下2 種鯉魚肌肉中的不同種類游離氨基酸含量受養殖模式及養殖品種影響,建鯉和易捕鯉雖在養殖中均當做鯉魚新品種,但二者遺傳背景差異極大[19-20],這種差異可能是導致同一養殖模式下2 種鯉魚肌肉游離氨基酸種類及含量彼此差異的內在因素。
3.3 養殖模式對2 種鯉血漿游離氨基酸含量的影響
血液循環系統作為動物營養物質運輸的唯一通道,其血漿中游離氨基酸水平與魚類新陳代謝水平密切相關,在魚類能量代謝中扮演著重要的角色。在堿性脅迫下,瓦氏雅羅魚Leuciscus waleckii 通過調控機體代謝提升血漿中的游離氨基酸水平以滿足機體所需能量,以維持機體正常生理機能[21]。本試驗結果顯示,在血漿中,建鯉在IPA 模式下除丙氨酸含量與對照差異不顯著外其他氨基酸含量均顯著增加,而易捕鯉除丙氨酸、天冬氨酸、天冬酰胺含量與對照差異不顯著外其他氨基酸含量均顯著下降。因此,我們推測建鯉在池塘模式下而易捕鯉在IPA 養殖模式下所需能量較高,血漿中游離氨基酸中的丙氨酸、天冬氨酸轉運至肝臟參與糖異生而滿足其生理能量需要[3],現有研究發現老鼠經長時間運動后其血漿中游離的丙氨酸和酸性氨基酸及其酰胺類氨基酸濃度下降[22];并且我們通過二者的選育背景可知[19-20],大頭鯉C.pellegrini作為選育易捕鯉的親本,其為中上層魚類且易捕鯉在親緣關系中偏向該魚而建鯉是底棲魚類[23-25],因此,這種生態位的差異可能是2 種鯉魚在不同養殖模式下能量利用差異的根本原因,因為流水模式能否促進魚類生長及提升能量利用效率在多種魚類的研究中結果大相徑庭[26]。
4 結論
IPA 養殖模式下,易捕鯉和建鯉均表現出更好的生長性能,但易捕鯉表現得更優;在游離氨基酸含量方面,2 種鯉魚肌肉中僅有少部分種類的氨基酸含量受養殖模式的影響,而在血漿中則絕大多數種類的游離氨基酸含量受養殖模式的影響,并且血漿中的絕大多數游離氨基酸含量的變化在2 種鯉魚中呈現出相反的趨勢。
