落葉生境蛭態輪蟲物種多樣性及四種中國新記錄種
2021-04-01 07:56:14汪文博楊宇峰
水生生物學報 2021年2期
關鍵詞:物種
汪文博 王 慶 李 瑩 曾 悅 楊宇峰
(暨南大學水生生物研究所, 廣州 510632)
輪蟲是一類水生或半水生小型無脊椎動物, 身體不分節, 兩側對稱, 具有假體腔, 已記載輪蟲2000余種[1]。輪蟲個體小、繁殖快, 能快速占領生態位, 并作為微食物網的重要環節, 將初級生產力(如藻類、細菌)轉化為次級消費者(如魚類、昆蟲幼蟲、枝角類和橈足類)可利用的形式, 在水生和濕地生態系統物質循環和能量流動中具有重要的生態學意義。
蛭態輪蟲是一類特殊的輪蟲類群, 具有可以套筒式收縮和舒張的身體; 除盤網輪科外均具有一對發達的輪盤, 兼有運動和攝食功能, 盤網輪科輪蟲頭冠扁平, 無輪盤, 腹面有一具纖毛的盤網; 兩對卵黃腺, 枝形咀嚼器。體長通常在200—500 μm, 某些種類如長足輪蟲(Rotaria neptunia.Ehrenberg, 1830)可長達1.6 mm。蛭態輪蟲具有兩大特點: 孤雌生殖和低濕休眠[2]。孤雌生殖使其在不利環境下, 免受種群豐度降低所帶來的滅絕風險, 并在環境條件適宜時迅速恢復種群數量。低濕休眠是指蟲體由于失水而進入休眠狀態, 形態特征和生理過程發生一系列變化, 從而在一定程度上抵御不利環境(干燥、低溫、紫外線)所造成的影響, 并在環境條件改善時復蘇[2]。因此, 蛭態輪蟲適應性極強, 分布十分廣泛, 大量存在于水體和陸生生境中。研究表明,生活在一般水體中的輪蟲, 20%—30%為蛭態輪蟲;在陸生生境中, 95%為蛭態輪蟲[2]。陸生生境包括苔蘚、地衣、土壤和落葉等。目前, 國內外蛭態輪蟲相關研究主要關注前3種陸生生境, 特別是對苔蘚中蛭態輪蟲研究最多。相比其他生境, 苔蘚中蛭態輪蟲物種豐富度和豐度均較高, 與其復雜的內部結構和高含水量有密切關系[3]。已有調查對落葉生境關注度較低[4,5], 偶見于部分蛭態輪蟲物種多樣性調查[6—8]。
落葉在森林生態系統中屬于凋落物層。凋落物是土壤生態系統物質和能量的主要來源, 對土壤質量維護和土壤生態功能發揮具有重要作用[9]。凋落物具有適合生物生存的微生境, 其中生活著大量土壤動物(如蜱螨目、彈尾目)。土壤動物在凋落物分解過程中發揮著十分重要的作用[10]。現有研究主要關注凋落物中大中型土壤動物群落結構, 低估甚至忽略了輪蟲現存量和生態作用。本研究旨在調查落葉生境蛭態輪蟲物種多樣性, 以期為進一步研究蛭態輪蟲在落葉凋落物分解中的作用提供基礎資料。
1 材料與方法
1.1 采樣地點與采樣方法
2018年11月至2019年5月, 共采集中國8省12地區28個落葉樣品, 采樣范圍涵蓋我國熱帶、亞熱帶及溫帶地區, 海拔跨度為3至1500 m(表1)。
落葉樣品通過手撿法采集, 隨即放入牛皮種子袋中, 并在種子袋表面標注落葉種類、地點、日期和海拔高度等信息。樣品帶回實驗室后, 室溫下自然風干后保存。
1.2 蛭態輪蟲的分離與提取
采用震蕩復蘇法分離落葉中蛭態輪蟲, 取部分風干后的落葉樣品放入250 mL錐形瓶中, 加入超純水至沒過樣品, 在培養箱中20℃恒溫黑暗培養24h使其復蘇。在培養完畢后, 將樣品放在渦旋振蕩儀上震蕩15s, 隨即將液體部分倒入30 μm孔徑網篩中過濾, 以上操作重復3次。用超純水沖洗網內殘留物至培養皿或5 mL浮游生物計數框內進行鏡檢觀察[11]。
1.3 形態學鑒定
將盛有蛭態輪蟲樣品的培養皿或浮游動物計數框置于體視顯微鏡下, 用玻璃毛細管挑取單個蟲體于載玻片上并添加少量超純水, 加蓋玻片后于具拍照系統的正立熒光顯微鏡下觀察其形態學特征,拍照作為鑒定依據, 通過TCapture成像軟件對蛭態輪蟲體長、體寬和頭冠寬等重要性狀進行測量, 測量系統和方式依據Yakovenko等[12]提出的形態學測量模型, 包括總長(TL)、體寬(BW)、頭冠寬(CW)、頭長(HL)、頭寬(HW)、頸長(NL)、最大頸寬(MxNW)、最小頸寬(MinNW)、咀嚼器長(RaL)、咀嚼器寬(RaW)、臀長(RL)、臀寬(RW)、腿長(FL)、腿寬(FW)、足刺長(SL)和足刺間隔寬(SSW)。使用Adobe Illustrator CC 2014繪制蛭態輪蟲形態模式圖。鑒定主要參考依據為王家楫[13]和Donner[14]的輪蟲分類專著及分類數據庫www.rotifera.hausdernatur.at。
1.4 新記錄種跛足突盤輪蟲Bradyscela clauda的COⅠ基因擴增與系統發育進化樹分析
本文采用HotSHOT法提取單只蛭態輪蟲DNA[15], 用毛細管挑取跛足突盤輪蟲B.clauda若干只于載玻片上, 超純水清洗2—3次至體表無雜質,將單個蟲體放入0.2 mL PCR管中, 加入堿性裂解液10 μL, 用顯微鑷破碎蟲體, 95℃裂解0.5h后取出立即放入-20℃冰箱內冷卻10—15min, 加入中和液10 μL,即完成DNA提取。使用引物(ZplankF1_t1: 5′-TGTAAAACGACGGCCAGTTCTASWAATCATAARGATATTGG-3′; ZplankR1_t1: 5′-CAGGAAA CAGCTATGACTTCAGGRTGRCCRAARAATCA-3′)進行PCR擴增, 擴增反應體系25 μL, 包含2×Gflex PCR Buffer (Mg2+, dNTP plus) 12.5 μL, 引物各0.5 μL,模板5 μL, Tks Gflex DNA Polymerase 0.5 μL,ddH2O 6 μL。引物反應程序: 95℃預變性1min,94℃變性40s→45℃退火40s→72℃延伸45s, 循環5次, 94℃變性40s→51℃循環退火40s→72℃循環延伸45s, 循環35次, 72℃延伸5min→保存[16]。1%瓊脂糖凝膠電泳檢測PCR產物, 若目標條帶較淺, 則將模版稀釋95%后用Bdell_COI_FW: 5′-CGTACWGAGTTAGGAATRGTA-3′和 Bdell_COI_Rev: 5′-CCAAAATTWCGATCTAAYA-3′引物進行巢式PCR擴增, 1%瓊脂糖凝膠電泳檢測PCR產物后測序。將本研究獲得序列于NCBI進行Blast比對, 下載得分最高的參考序列, 使用MEGA7軟件采用鄰接法(Neighbor-Joining method)構建系統發育進化樹。
2 結果
本研究共記錄蛭態輪蟲3科8屬29種(表2), 占全球已發現蛭態輪蟲物種多樣性的6.1%, 其中1屬(Bradyscela)和4種(B.clauda,Macrotrachela aculeataMilne, 1886,Philodina scabraMilne, 1916和Scepanotrocha corniculataBryce, 1910)為我國首次記錄。

表2 蛭態輪蟲物種名錄Tab.2 The species list of bdelloid rotifers
2.1 新記錄種形態學描述
跛足突盤輪蟲, 新記錄種Bradyscela clauda
身體淺褐色, 皮膚表面有顆粒狀點紋, 軀干側面有縱向皮膚褶, 形似蠕蟲狀。喙片二裂片, 其寬度約為最大頭寬的一半。頭長與頭寬相近, 爬行時,最大頭寬略小于軀干最寬處。兩側耙各具6個齒狀缺刻, 攝食時, 跛足突盤輪蟲的耙較靈活, 可以自由張合刮取食物。咀嚼器齒式為2/2, 背觸手短, 無眼點。足部短, 末端形成一吸盤狀圓盤, 第一偽足節中間位置有一環形腫脹, 第二偽足節末端有一圈21個短的、瘤狀的乳突。具三個短且粗壯的趾, 無刺戟。卵生。跛足突盤輪蟲足部伸展緩慢, 爬行運動遲鈍, 不具有如盤網輪屬輪蟲快速向前滑行的運動能力(圖1)。
尖刺粗頸輪蟲, 新記錄種Macrotrachela aculeata
體型呈紡錘形, 軀干皮膚表面規律排列有許多體刺, 該特征與多刺粗頸輪蟲M.multispinosa相似,但存在不同。尖刺粗頸輪蟲體刺較短, 規律排布在軀干每一偽體節上: 軀干第一和第二偽體節處各具一組體刺; 第三和第四偽體節處各具一組體刺, 且其中一組體刺位于軀干最寬處; 第五和第六偽體節處即臀部具三組體刺, 其中兩組體刺位于第六偽體節。尖刺粗頸輪蟲軀干部皮膚表面具有橫向和縱向的皮膚褶, 上述體刺按照皮膚褶分布規律排布,均朝向足部方向。喙長, 半圓形二裂片。在攝食狀態下, 輪盤寬度與頭寬一致, 具有發達的纖毛區和感覺剛毛; 瓣環發達, 從背面觀察, 兩側明顯凸出于頭部; 上唇片形成一“V”形缺口(圖2C和2D)。背觸手發達。咀嚼器齒式為2/2。足部第二偽體節背部中央有一明顯增厚凸出的附屬物。刺戟寬度較窄,逐漸變尖細, 有間距。無眼點。趾3個。卵生。
粗糙旋輪蟲, 新記錄種Philodina scabra
體型較小, 呈紡錘形。軀干皮膚粗糙, 有縱向皮膚褶, 呈灰褐色, 不透明。喙發達, 二裂片。攝食狀態下, 輪盤寬度小于頭部寬度, 兩個輪柱緊密靠在一起, 輪槽很窄; 上唇片形狀為平緩的弓形, 中間位置有一三角形突起。最突出的特征是倒數第二偽足節處著生有上下兩對刺戟, 上方一對刺戟基部較寬, 后逐漸變尖細, 無足間距, 下方一對刺戟較細,有足間距。這是迄今為止有記錄的唯一具有兩對刺戟的蛭態輪蟲。趾4個。咀嚼器齒式為1+2/2+1(圖3)。
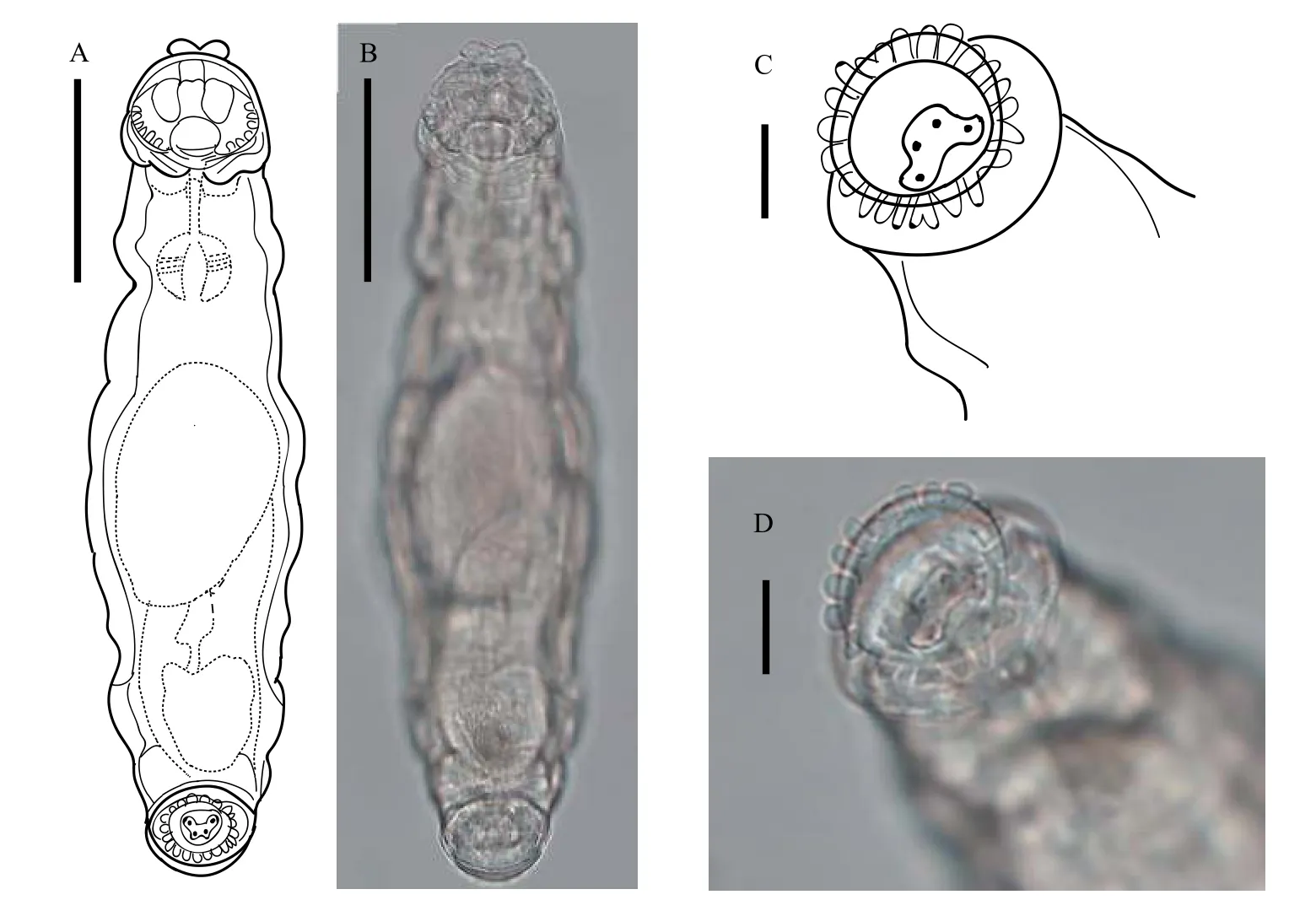
圖1 跛足突盤輪蟲Fig.1 Bradyscela clauda Bryce, 1893
角萼圍輪蟲, 新記錄種Scepanotrocha corniculata
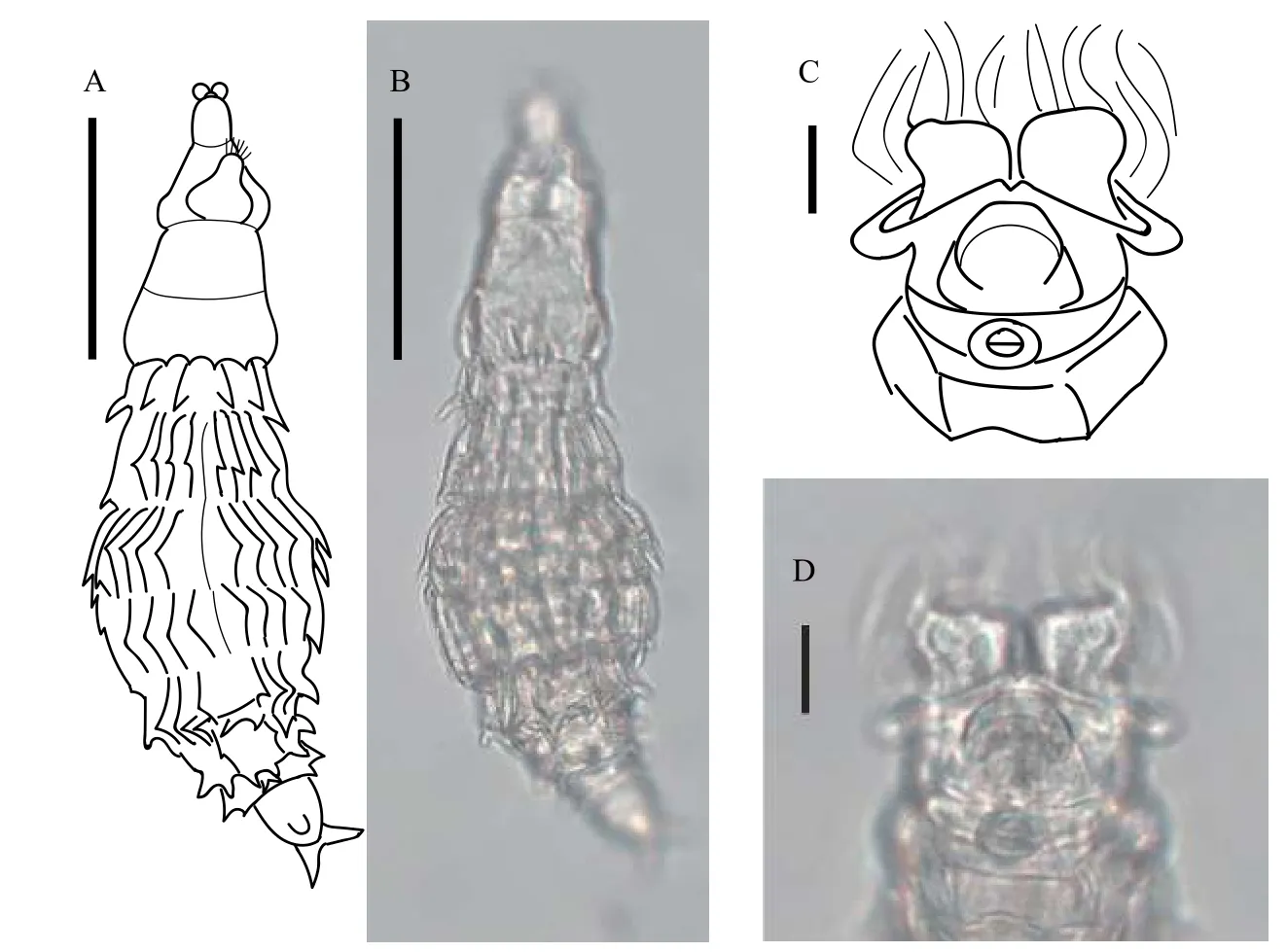
圖2 尖刺粗頸輪蟲Fig.2 Macrotrachela aculeata Milne, 1886
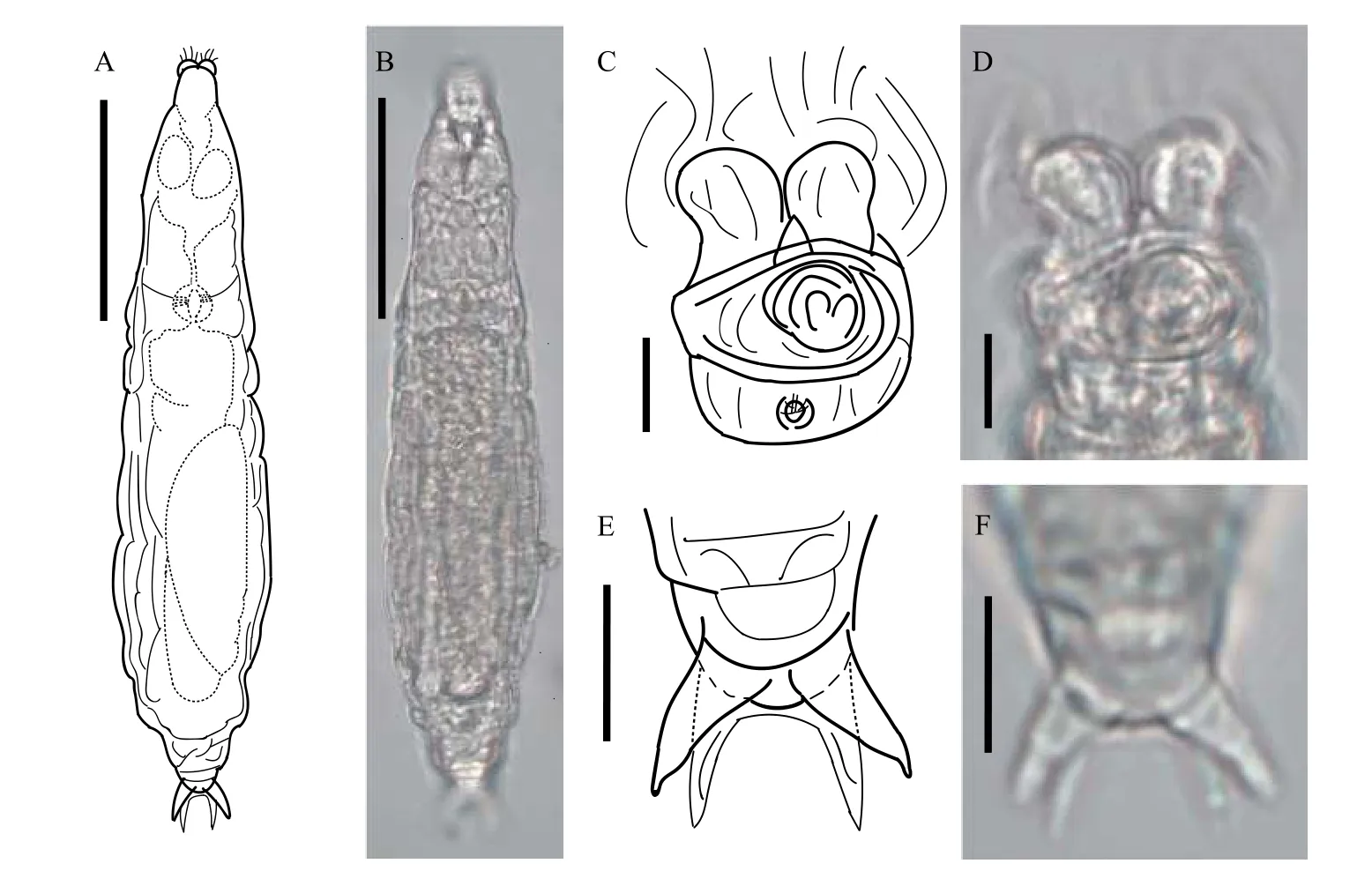
圖3 粗糙旋輪蟲Fig.3 Philodina scabra Milne, 1916
角萼圍輪蟲以食物泡為消化方式。體型小, 皮膚透明, 體表著生有許多小刺毛。圍輪屬的顯著分類特征是其輪盤周圍有一兜帽狀結構, 背面觀能觀察到兩個輪盤上伸出的發達纖毛, 兜帽結構可能是上唇片的一種變形。角萼圍輪蟲的兜帽狀結構寬度大于頭寬, 中央有一向下較深的凹陷, 且兜帽結構兩側中間位置有一水平向外的凸出。刺戟短, 足間距窄。趾2個, 短。咀嚼器齒式為4/4。卵生。爬行速度慢 (圖4)。
2.2 跛足突盤輪蟲, 新記錄種Bradyscela clauda進化樹分析及分子分類學地位
從GenBank中下載與本研究所獲得跛足突盤輪蟲B.clauda COⅠ基因序列相似度高的物種基因序列, 以旋輪科間盤輪屬Dissotrocha aculeataEhrenberg, 1830作為跛足突盤輪蟲B.clauda的外類群,盤網輪科盤網輪屬A.vaga和A.ricciae作為同科物種, 使用鄰接法構建系統發育進化樹。由圖5可知,本研究所獲得的兩條序列(N1-N2)與來自英國的跛足突盤輪蟲序列(EF173179.1)相似度最高, 聚為一支, 后與同科Adineta屬兩種輪蟲序列聚集在一起。聚類分析結果表明該輪蟲屬于蛭態亞綱盤網輪科突盤輪屬, 以COⅠ基因條形碼技術驗證了形態學鑒定的結果。
3 討論
3.1 新記錄種分類討論
跛足突盤輪蟲Bradycela clauda突盤輪屬在中國屬首次記錄。該屬共記錄輪蟲3種, 分別是B.clauda、B.granulosaDe Koning, 1947和B.hoonsooiSong, 2015, 三種形態學特征相似, 主要區別在于耙上齒狀缺刻數目、第二偽足節處有無乳突及乳突大小和數量[17]。跛足突盤輪蟲最早是Bryce(1893)在苔蘚生境中發現, 當時未將該種從盤網輪屬中分離出來, 僅作為盤網輪屬的一個新種記錄下來[18]。本研究所觀察的跛足突盤輪蟲與Bryce描述的種類存在差異, Bryce描述的種類第二偽足節處僅有10個乳突[18], 而本研究描述的種類第二偽足節處有23個乳突, 與Song[17]在韓國所觀察的種類乳突數量相同。Bryce[18]認為該輪蟲有許多變種。因此需要進一步觀察和比較, 并應用DNA條形碼等技術, 以確定其是否為同一種類的不同變種。
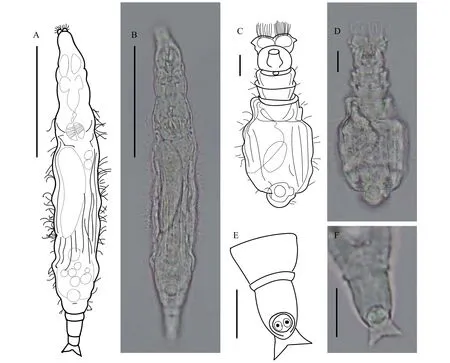
圖4 角萼圍輪蟲Fig.4 Scepanotrocha corniculata Bryce, 1910
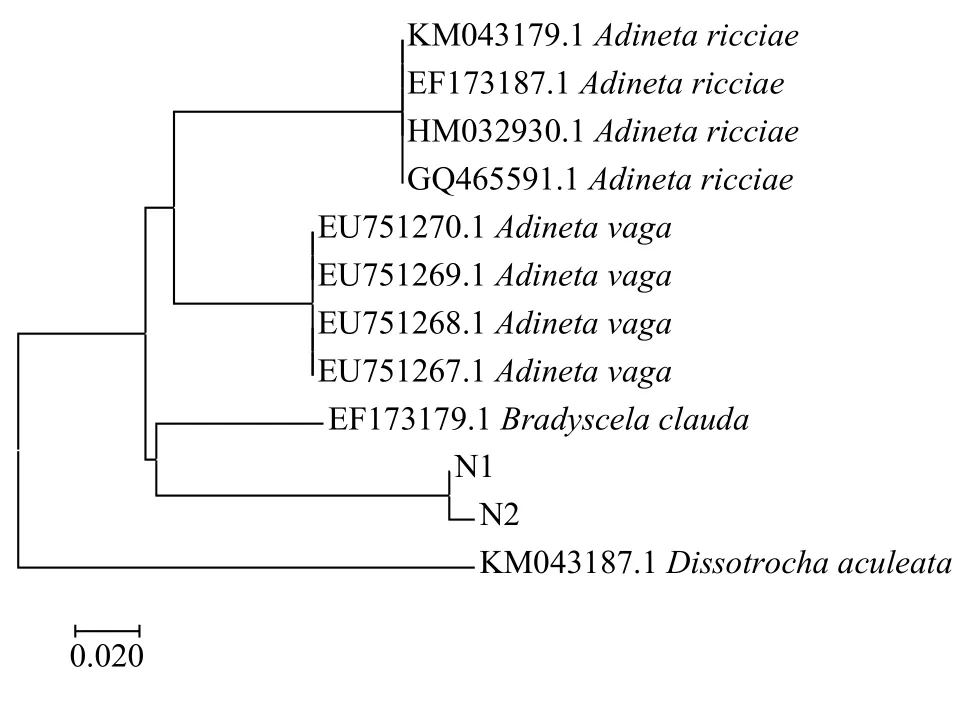
圖5 基于B.clauda COⅠ基因序列構建的NJ系統發育樹Fig.5 The neighbor-joining tree based on COⅠgene sequences of B.clauda
尖刺粗頸輪蟲Macrotrachela aculeata尖刺粗頸輪蟲最早由Milne[19]發現, 但由于樣本數量少, 故描述比較簡單, 并且對體刺數量和位置描述不夠準確, 與后來文獻記載存在出入。Murray[20,21]分別在南非和愛爾蘭發現了該種及形態特征不同的幾個變種并對它們進行了詳細描述, 其中南非的種類與本研究所觀察到的種類較相似, 均具7組體刺, 且排布方式相同。Barto?[22]在捷克境內也發現了該輪蟲。在他看來, 不同研究者對該輪蟲的描述存在很大出入, 這其中可能蘊藏著不同種類, 或者說該輪蟲具有很多變種。Donner[14]編寫的蛭態輪蟲檢索表中, 將粗頸輪屬中體表具有附屬物的輪蟲編輯在一起, 根據附屬物的形狀、大小、數量和在軀體上的排列進行分類。本研究是尖刺粗頸輪蟲在亞洲區系的首次記錄, 以往的研究主要集中在歐洲和南非。
粗糙旋輪蟲Philodina scabraMilne[23]在南非的苔蘚生境中首次觀察到粗糙旋輪蟲并對其進行了詳細描述, 形容其軀干皮膚粗糙有溝痕, 像樹皮一樣。該輪蟲到底存在幾個趾, Milne[23]尚無定論。同時, 該輪蟲無論體型還是輪盤、足部等細節特征, 都與粗頸輪屬相似。因此, Milne[23]對該輪蟲的分類歸屬保留懷疑態度, 暫且將其歸入旋輪屬。粗糙旋輪蟲較稀有, 很長一段時間世界上都沒有該輪蟲的相關報道。直到2017年, Song等[24]在韓國觀察到該輪蟲及其4趾, 最終確定了其分類歸屬。不過, 對于粗糙旋輪蟲兩對刺戟位置目前還存在爭議: Song等[24]記錄的種類較細一對刺戟位于上方,而較粗一對刺戟位于下方, 與Milne[23]原始文獻中描述情況相反。本研究觀察到粗糙旋輪蟲兩對刺戟排列位置與Milne[23]描述相同。這可能是由于顯微鏡下光與影的錯覺, 使Song等[24]得出了不同結論, 也可能是粗糙旋輪蟲存在不同變種。需要我們獲得更多樣本, 使用更先進的成像技術觀察并運用分子生物學手段比較。
角萼圍輪蟲Scepanotrocha corniculataBryce[25]在英國一苔蘚生境中最早發現角萼圍輪蟲。他認為該輪蟲一般特征與其他輪蟲并無區別, 只有在攝食狀態下, 才能觀察到其特有的兜帽結構。目前共記載圍輪屬蛭態輪蟲12種, 頭部均具有兜帽結構,彼此之間通過該結構的形狀、大小等特征加以區分[14]。其中, 角萼圍輪蟲S.corniculata與S.HaueriDonner, 1962,S.simplexDe Koning, 1947和S.parvaSchulte, 1954等相似。S.corniculata與S.simplex的區別在于前者兜帽結構前端中央存在一向下的凹陷, 而S.simplex不具有此凹陷; 與S.haveri之間區別在于S.corniculata的兜帽結構完全覆蓋住輪盤, 中央凹陷較深, 而S.haveri的兜帽結構剛好覆蓋住輪盤, 其邊緣與輪盤邊緣很接近, 但并不貼合, 且中央凹陷較淺; 至于S.parva, 該輪蟲兜帽結構兩側邊緣的中間位置無水平擴展[14]。另外, 本文所觀察的種類體表具有許多小刺毛, 這在Byrce[25]的報道和Donner[14]編寫的檢索表中皆未提到。推測本研究發現該輪蟲的采樣地點(浙江舟山)風力較大,S.corniculata為適應多風環境, 增大自身固著能力而形成這一變種。本研究是該輪蟲在亞洲區系的首次記錄。
3.2 蛭態輪蟲棲息生境研究現狀
關于蛭態輪蟲棲息地, 主要有兩種類型, 一是湖泊、河流、溪流等水生生境, 二是苔蘚、地衣、落葉和土壤等陸生生境。除此之外, 一些特殊生境也引起研究者們的極大興趣, 如種子、蘑菇等。就物種多樣性豐富程度而言, 與其他類群輪蟲不同的是, 95%的蛭態輪蟲生活在不穩定的陸生生境中,而棲息于淡水生境中的蛭態輪蟲, 常見的只有10種左右[13], 這可能與蛭態輪蟲低濕休眠和孤雌生殖的生活史策略有關。有研究者認為, 蛭態輪蟲經歷低濕休眠后, 繁殖力會顯著提高, 有助于保持適合度成分(Fitness components)的穩定[26]。并且, 嚴格孤雌生殖使它們能夠快速大量繁殖和傳播, 迅速占領棲息地, 快速適應多變或不穩定的陸生環境。
在陸生生境方面, 對苔蘚生境蛭態輪蟲研究最多(表3)。有研究者調查了2種不同生長年齡苔蘚與蛭態輪蟲物種組成之間的關系, 發現物種豐富度與苔蘚的生長年齡呈正相關, 且那些含水量高和結構復雜的苔蘚種類有更高的蛭態輪蟲豐度和物種多樣性[3]。目前, 蛭態輪蟲物種組成與其棲息的苔蘚物種特異性關系還未明晰, 物種對棲息生境植物種類的偏好不明顯[27]。其次, 研究較多的生境是土壤。Devetter[28]曾調查捷克境內克列特山保護區林下土壤蛭態輪蟲群落的時空變化, 發現輪蟲種類、豐度及物種多樣性與季節變化和土壤深度有密切關系; 他們認為土壤蛭態輪蟲群落是一原生群落,包含一些適應土壤生境的特有種。相比之下, 有關落葉生境中蛭態輪蟲群落結構時空變化的研究則較少, 大部分研究僅限于落葉生境蛭態輪蟲的定性觀察。主要原因有: (1)相較其他兩種陸地生境, 落葉生境不穩定。由于落葉脫離植物體, 喪失保水能力等自我保護和調節機制, 相較于苔蘚生境更易受外界環境變化的影響, 溫度、含水量等對生命活動至關重要的環境因子波動較大。(2)落葉本身在形態、結構、質量等方面差別較大, 即便是同種植物落葉, 其質量等特性隨腐化程度不同, 存在很大差別。相比之下, 同一樣點內土壤生境環境因子差別不大。目前對落葉生境中蛭態輪蟲群落結構的研究, 尚無科學定量標準。
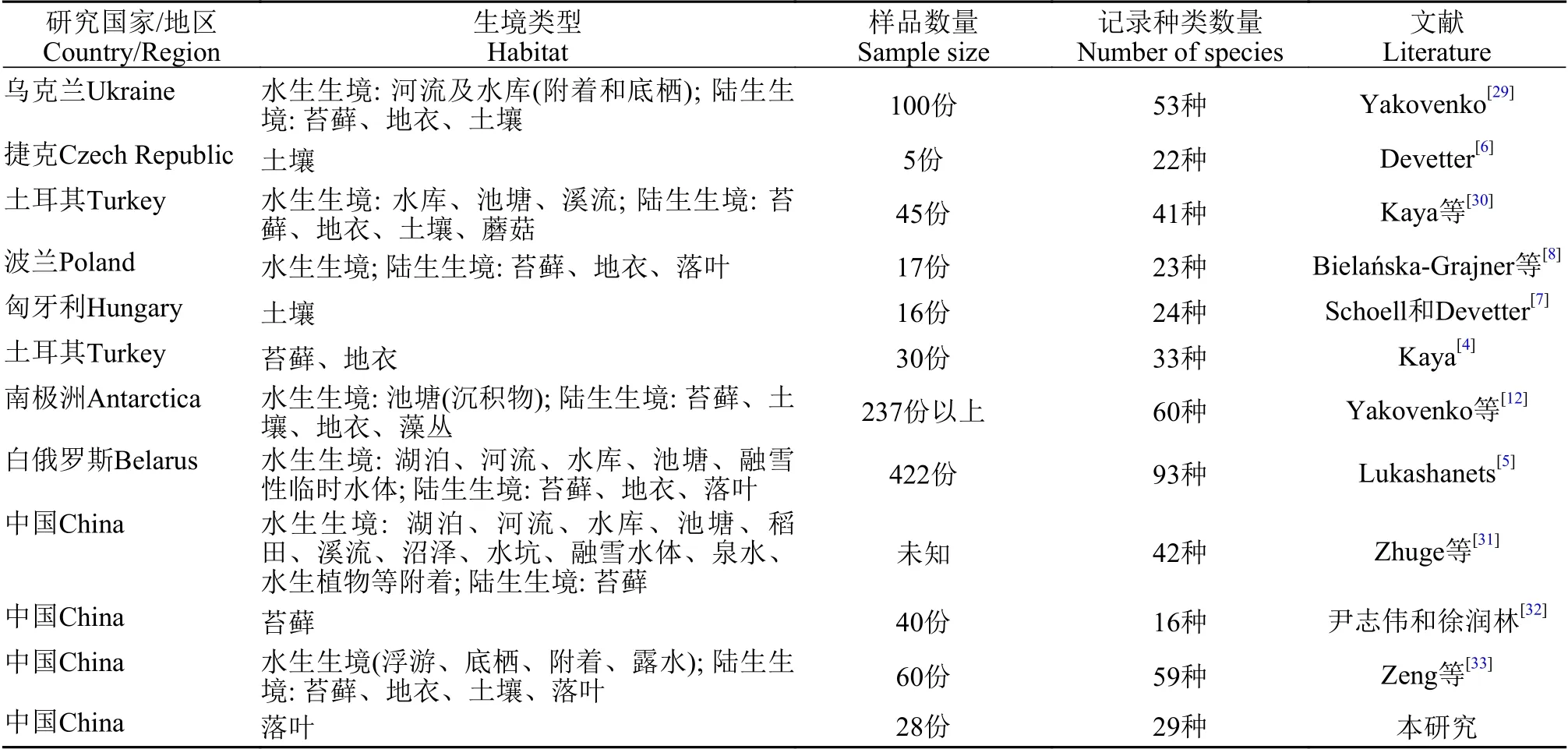
表3 不同生境蛭態輪蟲種類數量的比較Tab.3 Comparison of species number of bdelloid rotifers in the different habitats
3.3 落葉生境蛭態輪蟲研究展望
落葉作為一種特殊生境, 具有較高的研究價值。第一, 落葉及其他凋落物是森林生態系統物質循環中不可或缺的部分, 營養物質豐富, 是土壤肥力的重要來源之一[9]。研究表明, 凋落物組成對土壤化學性狀有顯著影響。第二, 落葉在分解過程中為土壤生物的生命活動提供了營養物質, 是腐食食物鏈的重要物質來源[34]。有研究發現, 凋落物分解過程對微生物有顯著影響, 植物未腐爛之前已有微生物存在, 并且真菌和細菌生物量會隨分解過程升高, 從而引起一些以微生物為食的小型和微型土壤動物群落結構發生變化, 為蛭態輪蟲生存和繁殖提供豐富的食物來源。同時, 包括蛭態輪蟲在內的土壤動物攝食活動也會對土壤細菌群落產生影響, 它們的排泄物促進微生物增殖, 進而推動落葉生境和森林生態系統的物質循環和能量流動。第三, 目前關于落葉生境中蛭態輪蟲群落結構及其時空變化的研究較少, 特別是落葉生境中蛭態輪蟲與微生物及其他土壤動物相互作用關系, 落葉腐化過程中蛭態輪蟲群落結構的變化等方面亟待深入研究。第四, 本研究對中國8省12地區落葉生境中蛭態輪蟲物種多樣性進行初步調查, 共鑒定出蛭態輪蟲29種,表明落葉生境蛭態輪蟲物種豐富。因此, 應加大對落葉生境蛭態輪蟲的調查力度, 以期發現更多新記錄種、特有種和新種。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
