幾種放射孢子蟲(chóng)的形態(tài)特征和分子鑒定
2021-04-01 07:56:16高志鵬習(xí)丙文
水生生物學(xué)報(bào) 2021年2期
高志鵬 楊 坤 陳 凱 習(xí)丙文, 謝 駿,
(1.南京農(nóng)業(yè)大學(xué)無(wú)錫漁業(yè)學(xué)院, 無(wú)錫 214081; 2.中國(guó)水產(chǎn)科學(xué)研究院淡水漁業(yè)研究中心,農(nóng)業(yè)農(nóng)村部淡水漁業(yè)和種質(zhì)資源利用重點(diǎn)實(shí)驗(yàn)室, 無(wú)錫 214081)
放射孢子蟲(chóng)是黏孢子蟲(chóng)生活史中寄生在無(wú)脊椎動(dòng)物宿主的發(fā)育階段; 魚(yú)體接觸有放射孢子蟲(chóng)水體是黏孢子蟲(chóng)病主要感染途徑之一[1]。自1899年Stolc[2]在顫蚓上首次發(fā)現(xiàn)放射孢子蟲(chóng)后的很長(zhǎng)一段時(shí)間, 放射孢子蟲(chóng)一直被認(rèn)為隸屬于黏體動(dòng)物門(mén)獨(dú)立的放射孢子蟲(chóng)綱[3]。1984年Wolf和Markiw首次證實(shí)虹鱒神經(jīng)系統(tǒng)寄生腦碘泡蟲(chóng)(Myxobolus cerebralis)生活史需要寡毛類(lèi)(正顫蚓)和魚(yú)類(lèi)兩替換宿主的發(fā)育過(guò)程[4]。該研究結(jié)果開(kāi)創(chuàng)性的將放射孢子蟲(chóng)和黏孢子蟲(chóng)的物種統(tǒng)一起來(lái)。此后, 黏體動(dòng)物分類(lèi)系統(tǒng)中放射孢子蟲(chóng)原有分類(lèi)階元被廢止(Suppress),不作為有效分類(lèi)類(lèi)群使用。為了容納此前報(bào)道大量未鑒定出對(duì)應(yīng)黏孢子蟲(chóng)的種類(lèi), 建議將原有屬名作為集合群(Collective group)區(qū)分不同形態(tài)的放射孢子蟲(chóng)[5,6]。目前共有20個(gè)放射孢子蟲(chóng)的集合類(lèi)群[3,7—11], 其中以三突放射孢子蟲(chóng)集合類(lèi)群報(bào)道種類(lèi)最多(約60種)[10]。由于放射孢子蟲(chóng)是魚(yú)類(lèi)黏孢子蟲(chóng)病的病原感染傳播重要途徑, 其相關(guān)研究受到越來(lái)越多關(guān)注。
目前, 魚(yú)類(lèi)寄生黏孢子蟲(chóng)報(bào)道有16科62屬2500多種[12—14], 其中一些種類(lèi)會(huì)導(dǎo)致嚴(yán)重的養(yǎng)殖或自然水體魚(yú)類(lèi)疾病: 如洪湖碘泡蟲(chóng)引起的鯽“喉孢子蟲(chóng)病”[15], 腦碘泡蟲(chóng)引起的鮭鱒的“旋轉(zhuǎn)病”[16],吳李碘泡蟲(chóng)引起的鯽“腹孢子蟲(chóng)病”等[17]。其中通過(guò)實(shí)驗(yàn)室感染或分子生物學(xué)方法闡析了約60種黏體動(dòng)物生活史[18]。王桂堂和姚衛(wèi)建[19]在2000年首次報(bào)道了國(guó)內(nèi)發(fā)現(xiàn)的1種三突放射孢子蟲(chóng)(Triactinomyxon); 習(xí)丙文等[3,20—22]報(bào)道了14種放射孢子蟲(chóng)并鑒定出了其中4種對(duì)應(yīng)的黏孢子蟲(chóng)種類(lèi): 武漢單極蟲(chóng)Thelohanellus wuhanensis)、吳李碘泡蟲(chóng)(Myxobolus wulii)、培養(yǎng)碘泡蟲(chóng)(Myxobolus cultus)和汪氏單極蟲(chóng)(Thelohanellus wangi)。趙丹丹等[23—25]報(bào)道了吉陶單極蟲(chóng)(Thelohanellus kitauei)、龜殼單極蟲(chóng)(Thelohanellus testudineus)和鲇楚克拉蟲(chóng)(Zschokkella parasiluri)的生活史。
本研究在對(duì)江蘇地區(qū)發(fā)病鯽塘口的放射孢子蟲(chóng)調(diào)查過(guò)程中, 發(fā)現(xiàn)了多種放射孢子蟲(chóng)。本文報(bào)道和描述了其中新發(fā)現(xiàn)的4個(gè)集合類(lèi)群的6種放射孢子蟲(chóng)。
1 材料與方法
1.1 池塘底泥寡毛類(lèi)采集和暫養(yǎng)
通過(guò)自制采泥器采集養(yǎng)殖池塘底泥(10—15 cm),用鋼篩(40目)篩除污泥, 收集寡毛類(lèi)帶回實(shí)驗(yàn)室在曝氣自來(lái)水中暫養(yǎng), 室溫(25±2)℃。
1.2 放射孢子蟲(chóng)的檢查和鑒定
吸取暫養(yǎng)水樣, 用浮游計(jì)數(shù)框在顯微鏡下(Olympus CX-31, 10倍物鏡)檢查是否有放射孢子蟲(chóng)。將由單條寡毛類(lèi)釋放的放射孢子蟲(chóng)在顯微鏡下拍照, 形態(tài)學(xué)數(shù)據(jù)測(cè)量采用系統(tǒng)自帶軟件(TAC-9.0), 單位為微米(μm)。測(cè)量后的放射孢子蟲(chóng)水樣收集到離心管中, -20℃保存?zhèn)溆谩?/p>
1.3 SSU rDNA序列擴(kuò)增
將保存的放射孢子蟲(chóng)水樣離心(6000 r/min,5min)富集蟲(chóng)體到1.5 mL離心管中, 用Lysis buffer for microorganism to direct PCR (TaKaRa) 粗提基因組-20℃保存。放射孢子蟲(chóng)SSU rDNA采用巢式PCR方法擴(kuò)增, 外引物ERIB1和ERIB10[26], 內(nèi)引物Myxosp-F和Myxosp-R[27]。第一輪PCR擴(kuò)增程序: 94℃預(yù)變性5min; 30個(gè)循環(huán), 94℃變性30s, 50℃退火45s, 72℃延伸1min; 最后延伸7min; 第二輪PCR擴(kuò)增程序:94℃預(yù)變性5min; 35個(gè)循環(huán), 94℃變性30s, 55℃退火45s, 72℃延伸45s; 最后延伸7min。PCR反應(yīng)體系: Ex Taq酶0.5 μL, 10×ExTaqbuffer 5 μL, dNTP 4 μL, 正反引物(10 μmol/L)各1 μL, 模板2—5 μL, 加雙蒸水至50 μL。PCR擴(kuò)增產(chǎn)物經(jīng)1.2%瓊脂糖凝膠電泳分離、純化回收, 送至生工生物工程(上海)測(cè)序。
1.4 DNA序列分析
測(cè)序獲得的SSU rDNA序列使用SeqMan (Lasergen package, DNAStar Inc.) 進(jìn)行拼接, 通過(guò)BLASTN(http://blast.ncbi.nlm.nih.gov/) 在NCBI GenBank數(shù)據(jù)庫(kù)中檢索同源性較高序列進(jìn)行比對(duì)和系統(tǒng)發(fā)育分析。序列多重比對(duì)采用Clustal X軟件[28]; 序列一致性和堿基替換最佳模型分析使用MEGA 7.0軟件[29]。系統(tǒng)發(fā)育分析采用MrBayes 3.1.2軟件的貝葉斯推論法(Bayesian Inference, BI)和MEGA 7.0軟件的最大似然法(Maximum likelihood, ML)。
2 結(jié)果
2018年4月至2019年6月在江蘇溧陽(yáng)、江陰、大豐和洪澤4個(gè)地區(qū)的鯽苗種塘和成魚(yú)塘采集的底棲寡毛類(lèi)水蚯蚓主要是蘇氏尾鰓蚓(Branchiura sowerbyiBeddard, 1892)。在檢查的3000多條蘇氏尾鰓蚓中共發(fā)現(xiàn)10種放射孢子蟲(chóng), 分別隸屬于6個(gè)集合類(lèi)群: 三突放射孢子蟲(chóng)(Triactinomyxon)2種、橘瓣放射孢子蟲(chóng)(Aurantiactinomyxon)2種、棘放射孢子蟲(chóng)(Echinactinomyxon)2種、新型放射孢子蟲(chóng)(Neoactinomyxum)2種、匈牙利放射孢子蟲(chóng)(Hungactinomyxon)1種和雷氏放射孢子蟲(chóng)(Raabeia)1種。其中4種與作者實(shí)驗(yàn)室前期報(bào)道描述種類(lèi)一致, 本文主要對(duì)其中新發(fā)現(xiàn)的6種放射孢子蟲(chóng)進(jìn)行鑒定和描述。
2.1 三突放射孢子蟲(chóng)DF
該類(lèi)型放射孢子蟲(chóng)發(fā)現(xiàn)于江蘇大豐(Dafeng),遵循放射孢子蟲(chóng)命名習(xí)慣將其命名為三突放射孢子蟲(chóng)DF(圖1和表1)。
形態(tài)學(xué)特征: 孢子具有長(zhǎng)橢圓形孢體, 有孢柄和3個(gè)錨狀尾突。孢體長(zhǎng)(22.62±1.58) μm (19.05—24.79 μm), 孢體寬(9.96±0.91) μm (19.05—12.34 μm);3個(gè)等長(zhǎng)的梨形極囊位于孢體頂端, 極囊長(zhǎng)(4.56±0.48) μm (3.96—5.79 μm), 極囊寬(2.06±052) μm(1.22—2.73 μm); 孢柄長(zhǎng)(72.35±1.00) μm (68.87—77.84 μm); 3個(gè)幾乎等長(zhǎng)錨狀尾突長(zhǎng)(142.99±7.39) μm(132.13—156.66 μm)。
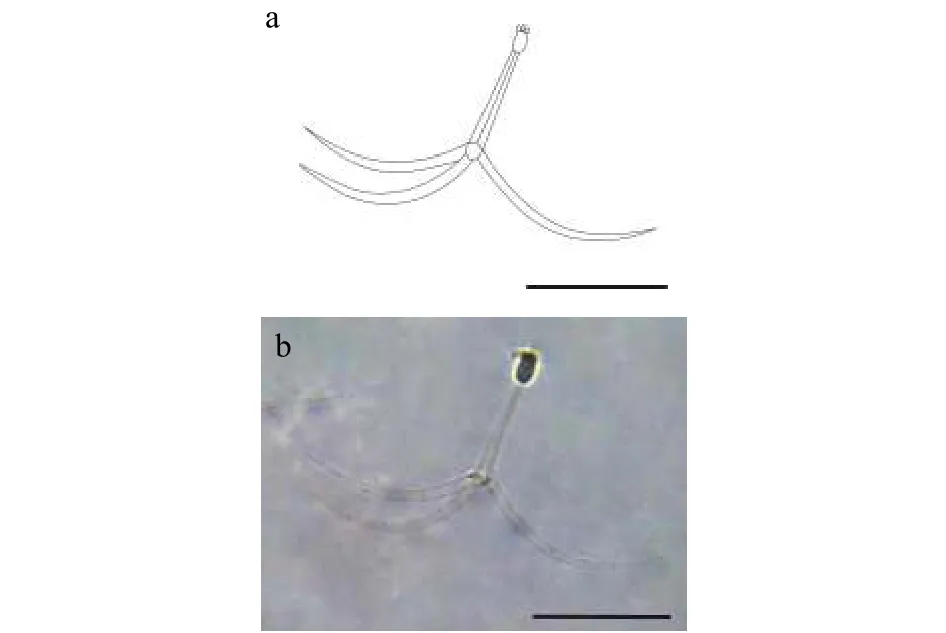
圖1 三突放射孢子蟲(chóng)DFFig.1 Triactinomyxon-type DF
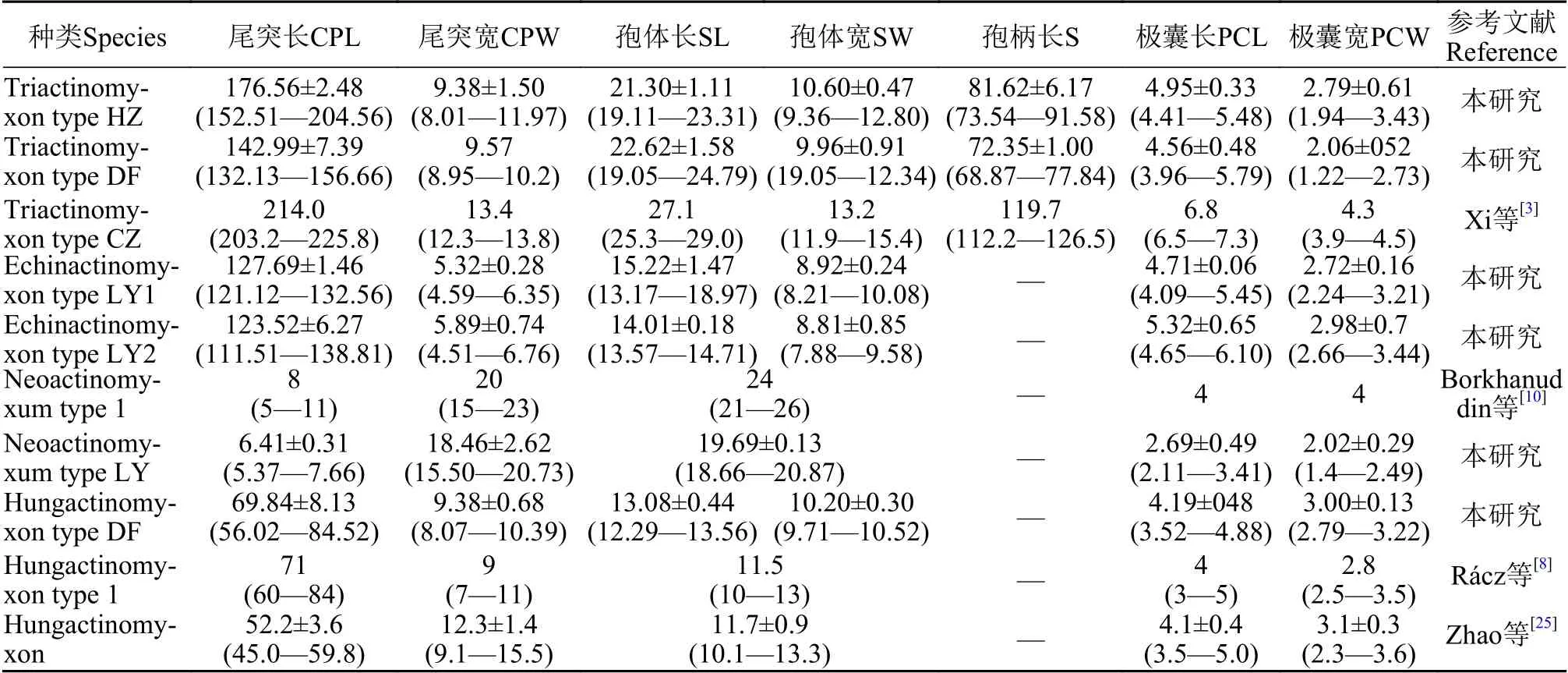
表1 本研究發(fā)現(xiàn)放射孢子蟲(chóng)及其相似種類(lèi)的形態(tài)測(cè)量數(shù)值(均值±標(biāo)準(zhǔn)差, 最小值—最大值)Tab.1 Morphormetric data of freshly actinosporeans released from the oligochaete Branchiura sowerbyi (mean±SD with ranges in parentheses; μm)
宿主: 蘇氏尾鰓蚓(Branchiura sowerbyi)
采樣地點(diǎn): 江蘇省鹽城市大豐區(qū)海豐農(nóng)場(chǎng)
感染率: 0.6%(3/500)
DNA序列: GenBank登錄號(hào)MN294777
2.2 三突放射孢子蟲(chóng)HZ
該類(lèi)型放射孢子蟲(chóng)發(fā)現(xiàn)于江蘇洪澤(Hongze),遵循放射孢子蟲(chóng)命名習(xí)慣將其命名為三突放射孢子蟲(chóng)HZ(圖2和表1)。
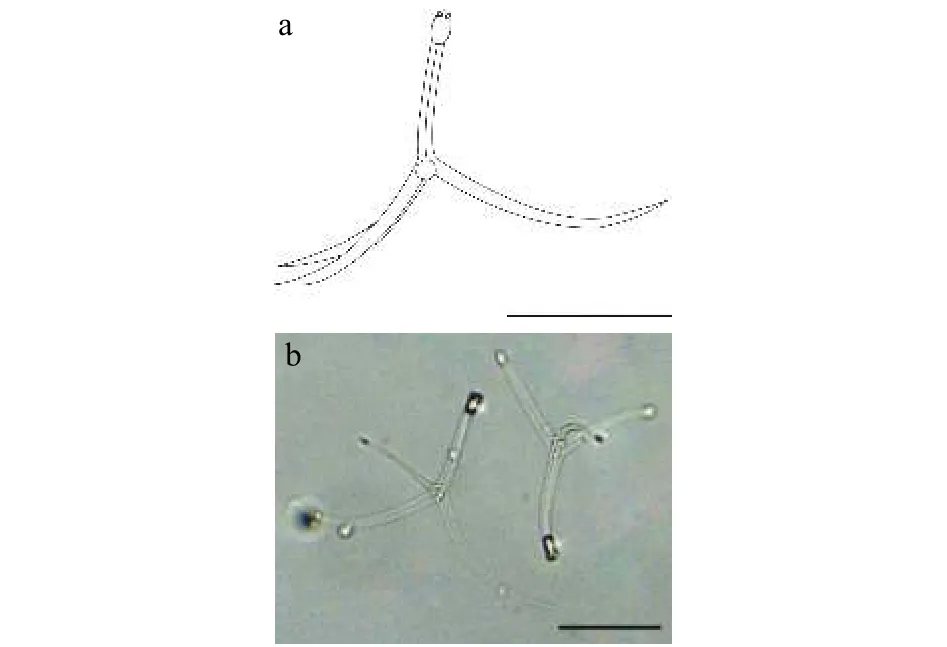
圖2 三突放射孢子蟲(chóng)HZFig.2 Triactinomyxon-type HZ
形態(tài)學(xué)特征: 該放射孢子蟲(chóng)形態(tài)與三突放射孢子蟲(chóng)DF較為接近。孢體正面觀呈長(zhǎng)柱形, 孢體長(zhǎng)(21.30±1.11) μm (19.11—23.31 μm), 孢體寬 (10.60±0.47) μm (9.36—12.80 μm); 3個(gè)等大極囊均勻分布在孢體頂端, 極囊長(zhǎng)(4.95±0.33) μm (4.41—5.48 μm),極囊寬 (2.79±0.61) μm (1.94—3.43 μm); 孢柄長(zhǎng)(81.62±6.17) μm (73.54—91.58 μm); 3個(gè)等長(zhǎng)的錨狀尾突長(zhǎng) (176.56±2.48) μm (152.51—204.56 μm)。
宿主: 蘇氏尾鰓蚓(Branchiura sowerbyi)
采樣地點(diǎn): 江蘇省淮安市洪澤區(qū)
感染率: 0.77%(5/650)
DNA序列: GenBank登錄號(hào)MN294778
2.3 棘放射孢子蟲(chóng)LY1
該類(lèi)型放射孢子蟲(chóng)發(fā)現(xiàn)于江蘇溧陽(yáng)(Liyang),遵循放射孢子蟲(chóng)命名習(xí)慣將其命名為棘放射孢子蟲(chóng)LY1(圖3和表1)。
形態(tài)學(xué)特征: 孢子具近卵圓形孢體和3個(gè)直棘尾突, 整體呈三角支架狀。孢體長(zhǎng) (15.22±1.47) μm(13.17—18.97 μm), 孢體寬 (8.92±0.24) μm (8.21—10.08 μm); 極囊長(zhǎng) (4.71±0.06) μm (4.09—5.45 μm),極囊寬 (2.72±0.16) μm (2.24—3.21 μm); 尾突長(zhǎng)(127.69±1.46) μm (121.12—132.56 μm), 尾突寬(5.32±0.28) μm (4.59—6.35 μm)。
宿主: 蘇氏尾鰓蚓(Branchiura sowerbyi)
采樣地點(diǎn): 江蘇省溧陽(yáng)前馬鎮(zhèn)
感染率: 0.15%(1/650)
DNA序列: GenBank登錄號(hào)MN294775
2.4 棘放射孢子蟲(chóng)LY2
該類(lèi)型放射孢子蟲(chóng)發(fā)現(xiàn)于江蘇溧陽(yáng)(Liyang),遵循放射孢子蟲(chóng)命名習(xí)慣將其命名為棘放射孢子蟲(chóng)LY2(圖4和表1)。
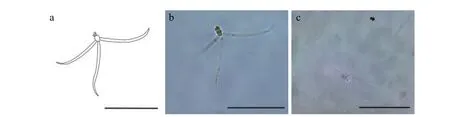
圖3 棘放射孢子蟲(chóng)LY1Fig.3 Echinactinomyxon-type LY1

圖4 棘放射孢子蟲(chóng)LY2Fig.4 Echinactinomyxon-type LY2
形態(tài)學(xué)特征: 孢子形態(tài)和測(cè)量學(xué)數(shù)據(jù)與棘放射孢子蟲(chóng)LY1相似; 孢體長(zhǎng) (14.01±0.18) μm (13.57—14.71 μm), 孢體寬 (8.81±0.85) μm (7.88—9.58 μm);3個(gè)等長(zhǎng)的極囊長(zhǎng) (5.32±0.65) μm (4.65—6.10 μm),極囊寬 (2.98±0.7) μm (2.66—3.44 μm); 3個(gè)等長(zhǎng)尾突長(zhǎng) (123.52±6.27) μm (111.51—138.81 μm), 尾突寬 (5.89±0.74) μm (4.51—6.76 μm)。
宿主: 蘇氏尾鰓蚓(Branchiura sowerbyi)
采樣地點(diǎn): 江蘇省溧陽(yáng)前馬鎮(zhèn)
感染率: 1.5%(10/650)
DNA序列: GenBank登錄號(hào)MN294776
2.5 新型放射孢子蟲(chóng)LY
該類(lèi)型放射孢子蟲(chóng)發(fā)現(xiàn)于江蘇溧陽(yáng)(Liyang),遵循放射孢子蟲(chóng)命名習(xí)慣將其命名為棘放射孢子蟲(chóng)LY(圖5和表1)。
形態(tài)學(xué)特征: 孢子頂面觀呈近似三角形, 尾突月牙形, 孢體呈球形, 3個(gè)極囊聚集在孢體頂點(diǎn)處凸起。孢體直徑長(zhǎng) (19.69±0.13) μm (18.66—20.87 μm);3個(gè)極囊長(zhǎng)(2.69±0.49) μm (2.11—3.41 μm); 月牙狀尾突長(zhǎng)約占孢體直徑的1/3, 長(zhǎng)(6.41±0.31) μm (5.37—7.66 μm), 寬 (18.46±2.62) μm (15.50—20.73 μm)。
宿主: 蘇氏尾鰓蚓(Branchiura sowerbyi)
采樣地點(diǎn): 江蘇省溧陽(yáng)市前馬鎮(zhèn)
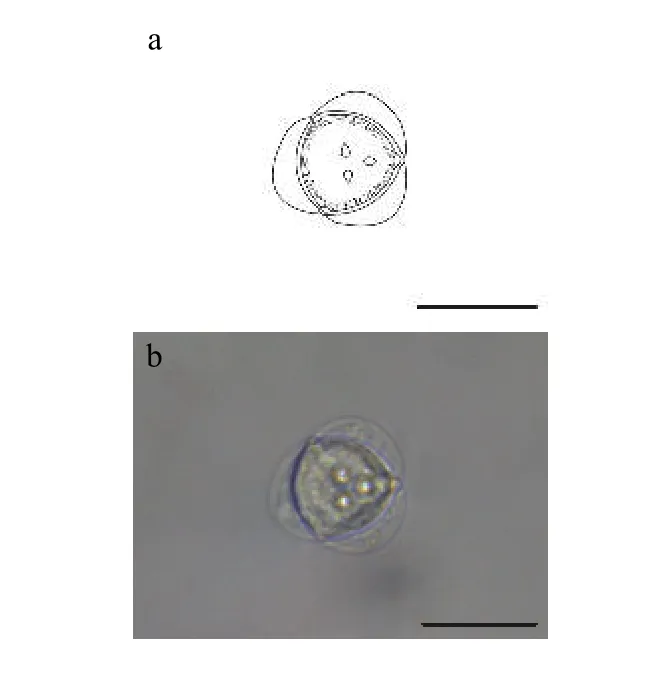
圖5 新放射孢子蟲(chóng)LYFig.5 Neoactinomyxum-type LY
感染率: 0.17%(1/600)
DNA序列: GenBank登錄號(hào)MN294779
2.6 匈牙利放射孢子蟲(chóng)DF
該類(lèi)型放射孢子蟲(chóng)發(fā)現(xiàn)于江蘇大豐(Dafeng),遵循放射孢子蟲(chóng)命名習(xí)慣將其命名為匈牙利放射孢子蟲(chóng)DF(圖6和表1)。
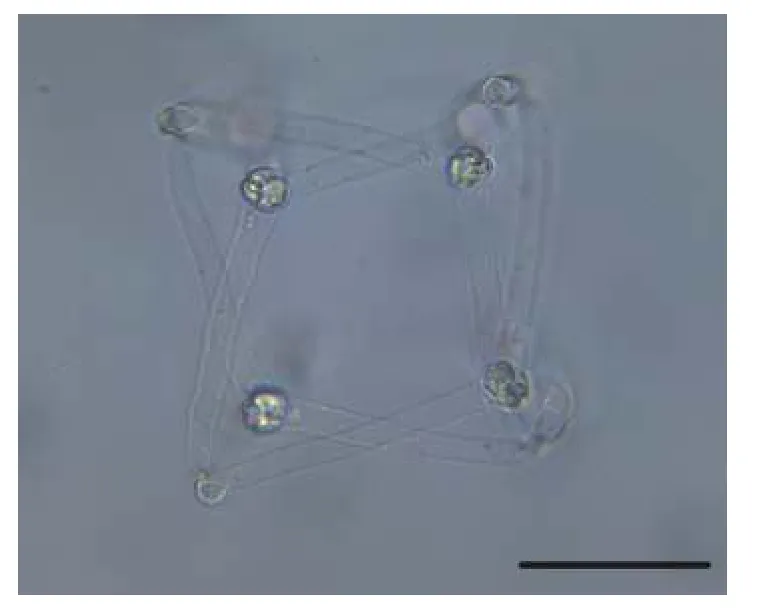
圖6 水體懸浮的匈牙利放射孢子蟲(chóng)DF, 比例尺=50 μmFig.6 Fresh floating Hungactinomyxon-type DF released by B.sowerbyi.Scale bars=50 μm
形態(tài)學(xué)特征: 由4個(gè)近似橘瓣放射孢子蟲(chóng)的尾突末端連接組成立方體, 與另一個(gè)由4個(gè)孢子組成的立方體交織形成網(wǎng)狀結(jié)構(gòu); 孢體近球形, 無(wú)孢柄,尾突直棘狀。單個(gè)孢子的孢體長(zhǎng)(13.08±0.44) μm(12.29—13.56 μm), 孢體寬(10.20±0.30) μm (9.71—10.52 μm); 不含有孢柄; 3個(gè)等長(zhǎng)的尾突(69.84±8.13) μm (56.02—84.52 μm), 尾突寬(9.38±0.68) μm(8.07—10.39 μm); 3個(gè)等大的極囊長(zhǎng)(4.19±048) μm(3.52—4.88 μm) , 極囊寬(3.00±0.13) μm (2.79—3.22 μm) 。
宿主: 蘇氏尾鰓蚓(Branchiura sowerbyi)
采樣地點(diǎn): 江蘇省鹽城市大豐區(qū)海豐農(nóng)場(chǎng)
感染率: 0.8%(4/500)
DNA序列: GenBank登錄號(hào)MN334159
2.7 SSU rDNA序列比對(duì)和系統(tǒng)發(fā)育分析
PCR擴(kuò)增獲得了6種新發(fā)現(xiàn)放射孢子蟲(chóng)的SSU rDNA序列片段, 其中三突放射孢子蟲(chóng)DF 845 bp(GenBank登錄號(hào)MN294777)、三突放射孢子蟲(chóng)HZ 856 bp (GenBank登錄號(hào)MN294778)、棘放射孢子蟲(chóng)LY1 830 bp (GenBank登錄號(hào)MN294775)、棘放射孢子蟲(chóng)LY2 842 bp (GenBank登錄號(hào)MN294776)、新放射孢子蟲(chóng)LY 830 bp (GenBank登錄號(hào)MN29-4779)、匈牙利放射孢子蟲(chóng)DF 843bp (GenBank登錄號(hào)MN334159)。序列同源性比對(duì)分析發(fā)現(xiàn), 三突放射孢子蟲(chóng)HZ和DF與文獻(xiàn)報(bào)道的Triactinomyxon type CZ (GenBank登錄號(hào)JX477771)同源性最高, 分別為93.39%和94.48%; 棘放射孢子蟲(chóng)LY1與Myxobolus lentisuturalis(GenBank登錄號(hào)AY119688)同源性最高(96.88%); 棘放射孢子蟲(chóng)LY2與Hungactinhomyxon type 1 (GenBank登錄號(hào)AY779062)相似性最高(97.51%); 新放射孢子蟲(chóng)LY與Thelohanellussp.相似性最高(92.48%)。匈牙利放射孢子蟲(chóng)DF和國(guó)外文獻(xiàn)報(bào)道的Hungactinomyxon序列相似度為99.41%—99.64%, 應(yīng)被鑒定為同一物種。
系統(tǒng)發(fā)育分析顯示棘放射孢子蟲(chóng)LY1和棘放射孢子蟲(chóng)LY2位于碘泡蟲(chóng)支, 其中棘放射孢子蟲(chóng)LY1與Myxobolus lentisuturalis親緣關(guān)系最近, 聚為姊妹枝; 三突放射孢子蟲(chóng)DF和三突放射孢子蟲(chóng)HZ位于四極蟲(chóng)屬分支; 新放射孢子蟲(chóng)LY聚在單極蟲(chóng)一枝。
3 討論
黏孢子蟲(chóng)病是當(dāng)前魚(yú)類(lèi)養(yǎng)殖中重要的寄生蟲(chóng)病之一, 如異育銀鯽洪湖碘泡蟲(chóng)病、鯉吉陶單極蟲(chóng)病等。由于缺乏明確有效治療藥物, 通過(guò)切斷病原生活史中魚(yú)體外感染傳播途徑是該類(lèi)疾病可行的有效防控措施。放射孢子蟲(chóng)作為黏孢子蟲(chóng)在無(wú)脊椎動(dòng)物替換宿主內(nèi)發(fā)育的重要階段, 受到魚(yú)病學(xué)和寄生蟲(chóng)學(xué)研究的廣泛關(guān)注[30]。早期在研究黏孢子蟲(chóng)生活史過(guò)程中需要在實(shí)驗(yàn)室完成無(wú)脊椎動(dòng)物感染[31]。由于實(shí)驗(yàn)復(fù)雜和耗時(shí)使得進(jìn)展比較緩慢, 而且對(duì)于部分種類(lèi)存在混合感染的孢子使結(jié)果的可靠性顯著降低[32—35]。近年來(lái), 分子生物學(xué)技術(shù)手段, 特別是SSU rDNA序列比對(duì)分析, 對(duì)黏孢子蟲(chóng)生活史的研究發(fā)揮重要促進(jìn)作用[18], 共計(jì)約50多種黏孢子蟲(chóng)在兩替換宿主體內(nèi)發(fā)育階段被準(zhǔn)確鑒定。如碘泡蟲(chóng)(Myxobolus)、單極蟲(chóng)(Thelohanellus)、尾孢蟲(chóng)(Henneguya)、擬尾孢蟲(chóng)(Myxobilatus)、球孢蟲(chóng)(Sphaerospora)、新角形蟲(chóng)(Ceratonova)、四極蟲(chóng)(Chloromyxum)、兩極蟲(chóng)(Myxidium)、楚克拉蟲(chóng)(Zschokkella)、霍氏蟲(chóng)(Hoferellus)以及微囊蟲(chóng)(Parvicapsula)等屬內(nèi)的不同種類(lèi)[18]。本研究在對(duì)放射孢子蟲(chóng)鑒定中也采用了SSU rDNA序列比對(duì)分析,然而并沒(méi)能確定所新發(fā)現(xiàn)的6種類(lèi)型所對(duì)應(yīng)的魚(yú)體寄生黏孢子蟲(chóng)種類(lèi)。可能的原有主要有兩個(gè)方面,一是魚(yú)體寄生黏孢子蟲(chóng)類(lèi)有2500多種, 目前NCBI數(shù)據(jù)庫(kù)中可供參考的序列非常有限; 或者由于黏孢子蟲(chóng)物種多樣性非常高, 有些種類(lèi)在魚(yú)體還沒(méi)有被發(fā)現(xiàn)和報(bào)道。隨著魚(yú)類(lèi)黏孢子蟲(chóng)物種多樣性研究的深度和廣度的不斷增加, 本研究所報(bào)道放射孢子蟲(chóng)及其DNA序列將提供重要的參考依據(jù)。
放射孢子蟲(chóng)的形態(tài)鑒定主要依據(jù)孢體形狀、孢柄有無(wú)、尾突發(fā)生位置和形狀及大小。目前三突放射孢子蟲(chóng)是已報(bào)道種類(lèi)最多的集合類(lèi)群。本研究所發(fā)現(xiàn)的Triactinomyxon HZ與文獻(xiàn)中江蘇常州養(yǎng)殖池塘發(fā)現(xiàn)的Triactinomyxon type CZ形態(tài)非常相似, 但在孢子大小上存在差異; 前者尾突和孢柄明顯短于后者(176.56 μmvs.214.0 μm, 81.62 μmvs.119.7 μm, 表1)。本研究所報(bào)道的Triactinomyxon DF在孢子大小上顯著小于Triactinomyxon HZ(表1)。SSU rDNA分析發(fā)現(xiàn)序列一致性低于95%,進(jìn)一步確認(rèn)這些孢子大小不同的三突放射孢子蟲(chóng)屬于不同物種。本研究所發(fā)現(xiàn)的Neoactinomyxum LY與該集合類(lèi)群中其他種類(lèi)主要區(qū)別特征在于月牙狀尾突, 但與Borkhanuddin等[10]報(bào)道的Neoactinomyxum type 1形態(tài)較為相似; 二者主要區(qū)別在于孢子尾突長(zhǎng)和寬大小(6.41 μmvs.8 μm, 18.46 μmvs.20 μm, 表1)。本研究發(fā)現(xiàn)的Echinactinomyxon LY1和LY2的尾突的末端都有明顯的分叉結(jié)構(gòu), 可以與文獻(xiàn)中報(bào)道的Echinactinomyxon種類(lèi)明顯區(qū)分開(kāi)。Echinactinomyxon LY1和LY2的形態(tài)數(shù)值非常接近很難區(qū)分, 然而SSU rDNA序列(MN294775和MN294776, 96.7%)卻存在很大差異, 表明這2種放射孢子蟲(chóng)為不同物種。該結(jié)果進(jìn)一步說(shuō)明由于放射孢子蟲(chóng)形態(tài)結(jié)構(gòu)簡(jiǎn)單, 僅僅依賴形態(tài)特征容易存在錯(cuò)誤鑒定, 分子生物學(xué)手段是物種鑒定的有力佐證。
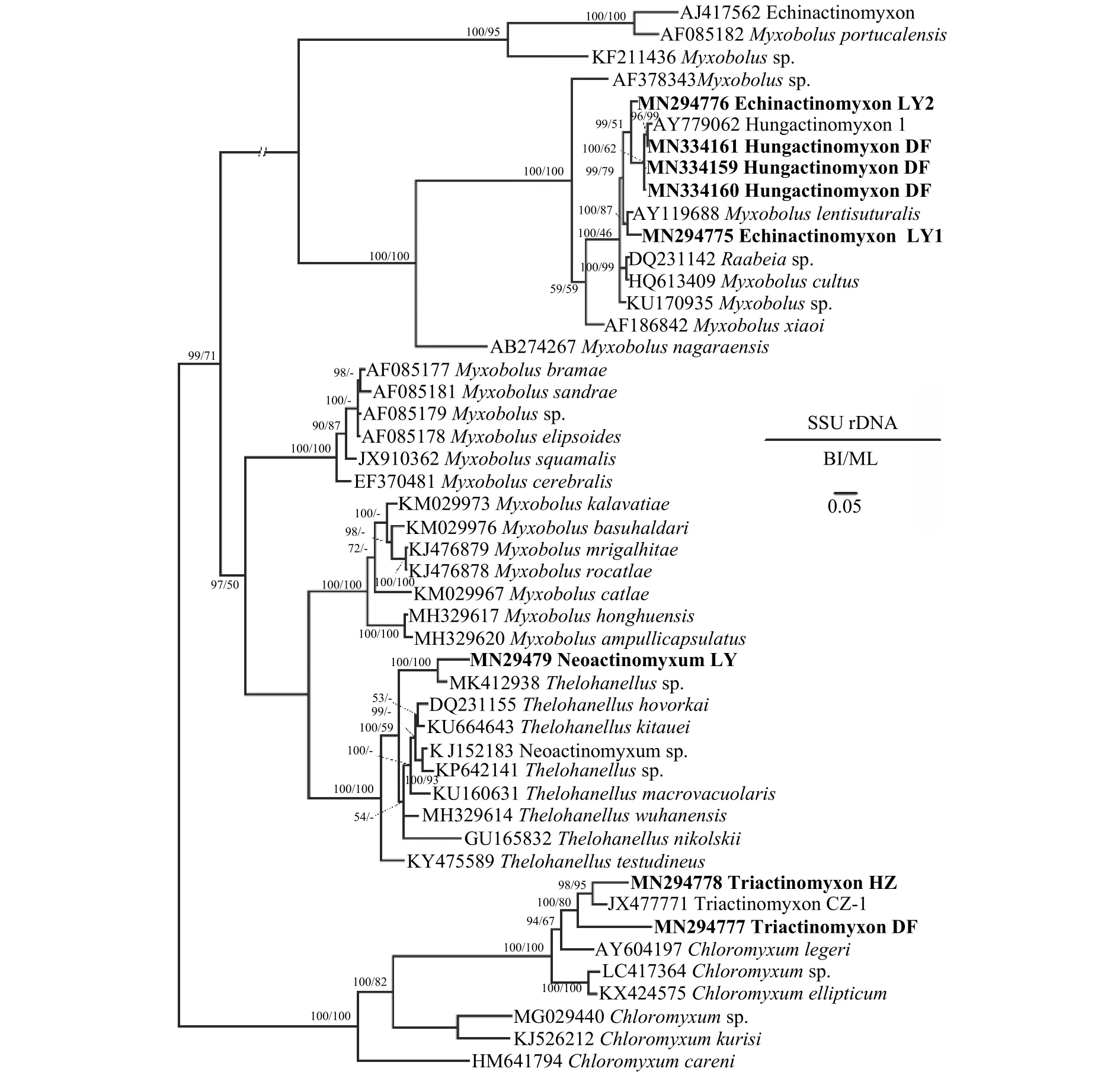
圖7 基于黏體動(dòng)物SSU rDNA序列建立的系統(tǒng)發(fā)育樹(shù)Fig.7 Phylogenetic tree of myxozoans based on the SSU rDNA sequences
本研究發(fā)現(xiàn)的Hungactinomyxon DF, 其形態(tài)與Ráczdeng等[8]在匈牙利蘇氏尾鰓蚓中發(fā)現(xiàn)的Hungactinomyxon type 1及趙丹丹等[25]在湖北大同湖和東西湖異育銀鯽苗種塘發(fā)現(xiàn)的Hungactinomyxon蟲(chóng)體形態(tài)上非常相近。本研究發(fā)現(xiàn)的Hungactinomyxon蟲(chóng)體與趙丹丹等[25]描述的放射孢子蟲(chóng)在形態(tài)上有些不同, 如尾突長(zhǎng)比趙丹丹等[25]描述的更長(zhǎng)(69.84 μmvs.52 μm), 尾突更細(xì)(9.38 μmvs.12.3 μm); 與Rácz描述的匈牙利放射孢子蟲(chóng)形態(tài)更接近。此外, 盡管中國(guó)和匈牙利地理相隔很遠(yuǎn)且屬于不同動(dòng)物地理區(qū)系, 但SSU rDNA序列分析表明它們具有>99%的序列一致性。因此, 本文報(bào)道的Hungactinomyxon和匈牙利報(bào)道的Hungactinomyxon type 1應(yīng)被鑒定為同一物種的不同地理種群。20世紀(jì)歐洲曾從亞洲地區(qū)引進(jìn)鯉科魚(yú)類(lèi)(鯉、鰱、鳙等)用于養(yǎng)殖和水環(huán)境調(diào)控。Hungactinomyxon type 1[8]在匈牙利的出現(xiàn)極有可能是黏孢子蟲(chóng)隨魚(yú)類(lèi)宿主種群擴(kuò)散的結(jié)果, 同時(shí)其生活史中所需的替換宿主蘇氏尾鰓蚓也是世界廣布種。
系統(tǒng)發(fā)育分析表明本研究所發(fā)現(xiàn)的Echinactinomyxon和Hungactinomyxon與Raabeia類(lèi)型放射孢子蟲(chóng)有較近的親緣關(guān)系且都聚集在碘泡蟲(chóng)(Myxobolusspp.)分支中(圖7)。此前的研究也發(fā)現(xiàn)碘泡蟲(chóng)屬種類(lèi)對(duì)應(yīng)的放射孢子蟲(chóng)集合類(lèi)群具有很高的多樣性[36]。此外, Rocha等[37]認(rèn)為Raabeia類(lèi)群的尾突形態(tài)具有很高的可塑性, 尾突的直和彎曲不能作為Echinactinomyxon和Raabeia類(lèi)群的區(qū)分特征; 建議將Echinactinomyxon和Raabeia合并。本研究認(rèn)為目前放射孢子蟲(chóng)集合類(lèi)群名稱(chēng)不是有效的分類(lèi)階元, 不具有實(shí)際分類(lèi)學(xué)意義; 為了避免描述上的混亂仍然保留Echinactinomyxon集合類(lèi)群。
由于本研究所報(bào)道的放射孢子蟲(chóng)DNA序列與目前可用于比對(duì)分析的魚(yú)體寄生黏孢子蟲(chóng)序列一致性都低于99.0%, 超出普遍認(rèn)為的種內(nèi)差異1%;因此, 未能鑒定出對(duì)應(yīng)的黏孢子蟲(chóng)物種。系統(tǒng)發(fā)育分析表明三突放射孢子蟲(chóng)Triactinomyxon HZ和DF聚在四極蟲(chóng)屬分支、Neoactinomyxum LY聚在單極蟲(chóng)Thelohanellus分支、Echinactinomyxon LY1和LY2及Hungactinomyxon DF聚類(lèi)在碘泡蟲(chóng)分支;這些放射孢子蟲(chóng)應(yīng)分別對(duì)應(yīng)了四極蟲(chóng)、單極蟲(chóng)和碘泡蟲(chóng)屬的6種不同黏孢子蟲(chóng)種類(lèi)。四極蟲(chóng)通常為腔寄生, 感染魚(yú)的膽囊和膀胱[38]; 碘泡蟲(chóng)和單極蟲(chóng)可感染魚(yú)幾乎所有的組織器官[39]。進(jìn)一步開(kāi)展池塘養(yǎng)殖魚(yú)類(lèi)寄生黏孢子蟲(chóng)多樣性調(diào)查有助于揭示本研究所發(fā)現(xiàn)放射孢子蟲(chóng)感染傳播途徑。
總之, 本研究基于形態(tài)和分子生物學(xué)特征描述了6種新發(fā)現(xiàn)的放射孢子蟲(chóng), 進(jìn)一步豐富了黏孢子蟲(chóng)物種多樣性的研究?jī)?nèi)容, 為揭示魚(yú)類(lèi)寄生黏孢子蟲(chóng)病原的生活史提供基礎(chǔ)數(shù)據(jù)。
