影響豬肌間脂肪沉積的主要因素及潛在調控機理
2021-04-13 02:40:56楊茜梓胡睿智賀建華伍樹松
動物營養(yǎng)學報 2021年3期
關鍵詞:水平
楊茜梓 胡睿智 賀建華 伍樹松,2*
(1.湖南農業(yè)大學動物科學技術學院,長沙410128;2.湖南省畜禽安全生產協同創(chuàng)新中心,長沙410128)
豬肉是我國居民日常膳食的主要肉類來源。2017年,我國人均豬肉消費量是世界其他國家平均水平的4.6倍,豬肉產量約占全球產量的48.19%[1]。為提高畜產品的產量,我國早期研究主要追求畜禽的生長速度、飼料轉化率和瘦肉率。長期的品種選育形成了目前以增長快、瘦肉率高為特征的畜禽品種。隨著經濟社會的發(fā)展和人民生活水平的提高,消費者在對豬肉及肉制品的需求量提高的同時,對肉品質的要求也越來越高。現如今瘦肉型豬品種的繁育已經很成熟,但瘦肉率的提高,伴隨著的是肌間脂肪的降低。而中國許多優(yōu)秀的地方品種由于具有更高的肌間脂肪含量,也有著更好的肉質。肌間脂肪在某些方面是比較特殊的脂肪組織,其含量的多少表現為肌肉間大理石花紋的評分,并且與豬肉的風味、嫩度和多汁性等性狀密切相關,是決定豬肉品質的重要指標之一[2]。因此,控制肌間脂肪沉積量對于提高肉品質有重要作用。肌肉的脂肪沉積受多種因素影響,如遺傳、營養(yǎng)、環(huán)境、外源激素等,外源激素一般指人工合成的或人工提取的與機體分泌的激素發(fā)揮相同作用的化學物質[3]。但目前關于靶向調控肌間脂肪沉積的研究較少,而肌間、肌內和皮下等不同的脂肪組織具有相似的代謝特征和沉積機理[4],因此探究各因素對肌間脂肪沉積的調控作用及相關機理尤為關鍵。本文結合國內外研究現狀對肌間脂肪沉積規(guī)律和營養(yǎng)調控手段進行綜述,以期為提高豬肉品質提供理論依據。
1 肌間脂肪概述
肌肉中的脂肪組織分為肌內脂肪和肌間脂肪[5],二者的功能與特點相似但又存在區(qū)別,但對于改善肉品質均有重要作用。肌內脂肪存在于肌內膜、肌外膜和肌束膜上[6]。肌間脂肪存在于肌纖維之間[7],主要在豬體重達到20 kg前發(fā)育[8]。雖然肌間脂肪和皮下脂肪的生長速率和組成相近[9],但有研究表明腹部肌間脂肪的沉積速率比皮下和腎周脂肪快[10-11],與皮下脂肪以及腎周脂肪相比,肌間脂肪的脂質含量最低,且不飽和程度高于腎周脂肪,低于皮下脂肪[10]。
研究表明肌間脂肪占總脂肪的20%~35%[8,11],肌間脂肪與總脂肪含量呈正相關關系[12]。肌間脂肪的含量直接影響肉的風味、組織的系水力和肉色[13]。雖然大量的研究集中在肌內脂肪對肉品質的影響[14-15],但在意大利杜洛克豬的育種計劃中,肌肉中的肌間脂肪含量已被作為其干腌火腿的品質特征之一[16]。肌間脂肪也是決定肉產品是否優(yōu)質的關鍵因素,因此研究影響肌間脂肪沉積的因素以及沉積的機理對于提高肉品質有著重要作用。
2 肌間脂肪的主要調控因素及潛在機理
2.1 肌間脂肪的種間差異
品種直接決定豬的遺傳性狀,不同品種豬的肌間脂肪含量不同。肌間脂肪含量在我國地方品種與國外選育品種豬之間存在明顯差異。有研究表明,在體重為20~90 kg的生長期內,合成系配套雜交豬、大白公豬、大白母豬和梅山豬的肌間脂肪含量分別為4.29、4.56、5.22和7.62 kg[17]。李盼等[18]對地方品種藏豬和滇南小耳豬與大白豬脂肪沉積差異進行的研究表明,藏豬和滇南小耳豬硬脂酰輔酶A去飽和酶(stearoyl-CoA desaturase,SCD)表達量顯著高于大白豬,SCD主要調節(jié)脂肪酸的生物合成。陶璇等[19]研究發(fā)現SCD基因在藏豬和杜洛克豬中表達量與肌內脂肪呈正相關。淮南豬脂肪組織中脂肪酸合酶(fatty acid synthase,FAS)的活性顯著高于長白豬[20]。可見我國地方品種豬具有優(yōu)良的肌間和肌內脂肪沉積能力。此外,Wood等[21]研究表明,大白豬生長速度快于杜洛克豬,且脂肪更薄,杜洛克豬背最長肌含有較高的脂質濃度和較低的水分含量。造成上述種間差異的主要原因之一是不同品種豬的脂肪沉積相關基因表達水平不同。
2.2 表觀遺傳對肌間脂肪沉積的影響
表觀遺傳學是指在基因的DNA序列沒有發(fā)生改變的情況下,基因的功能發(fā)生可遺傳的變化,并最終導致表型的變化。基因修飾、蛋白質與蛋白質、DNA和其他分子的相互作用會影響和調控基因的表達、功能和特性,這在一定程度上解釋了為什么同一品種豬的不同組織、不同部位的脂肪含量不相同。DNA甲基化是調控基因轉錄活性和表達的典型因素之一,其與基因的表達水平呈負相關[22]。已有報道指出,DNA的甲基化能影響脂肪的沉積[23]。例如瘦肉型的長白豬背脂與ATP合酶活性相關基因甲基化水平高于中國地方品種榮昌豬[24],表明較低能量代謝水平可能導致脂質合成效率降低。瘦肉型(杜洛克)豬與肥胖型(陸川)豬脂肪組織基因組差異甲基化區(qū)域比較后發(fā)現,瘦肉型豬的DNA甲基化水平較肥胖型豬高[25]。肥胖小鼠的過氧化物酶體增殖物激活受體(peroxisome proliferators-activated receptor,PPAR)α DNA甲基化水平顯著高于正常小鼠[26]。此外,還有研究表明豬淺表脂肪組織中與脂質代謝相關的基因相比與深層脂肪組織,DNA甲基化程度更高[27],這證明不同的脂肪組織甲基化程度也存在不同。
2.3 肌間脂肪沉積的相關分子機制
研究表明,肌間脂肪的含量與總脂肪量密切相關[17],而直接影響脂肪沉積的主要原因是脂肪酸的運轉、脂肪合成代謝以及分解代謝的相關蛋白表達[28]。大量研究表明,腺苷酸活化蛋白激酶(AMP-activated protein kinase,AMPK)信號通路是調控脂肪沉積與分解的關鍵通路。
AMPK是一種絲氨酸(Ser)/蘇氨酸(Thr)激酶[29],關鍵功能是調節(jié)細胞內的能量平衡[30]。AMPK包括α催化亞基、β和γ調節(jié)亞基,α亞基的N端包含Ser/Thr激酶區(qū)和1個需要被磷酸化的Thr172位點,C端主要負責聯系β亞基;β亞基包含1個糖原結合區(qū)和1個聯系α、γ亞基的C末端結合區(qū);與AMP、ADP結合的γ亞基,能夠促進α亞基的Thr172位點被上游激酶磷酸化,從而激活AMPK[31]。AMPK的激活受到AMP與ATP比值的調節(jié),當細胞的ATP水平降低,通過高濃度5’-AMP和AMPK的γ亞基相互作用來激活AMPK,AMPK會通過降低生物合成來保存ATP[32],并通過響應激素和營養(yǎng)信號來參與全身能量平衡的調節(jié),其激活可促進ATP生成機制,如誘導脂肪酸氧化,同時抑制脂肪生成和脂肪細胞分化,并刺激肌肉對葡萄糖的攝取[33-34]。而蛋白磷酸酯酶-1(PP1)、蛋白磷酸酯酶-2A(PP2A)和蛋白磷酸酯酶-2C(PP2C)3種蛋白激酶則能使Thr172殘基去磷酸化,抑制AMPK的激活。中國地方豬民豬相較于大白豬,因為AMPK的低水平表達,從而有更高的脂肪沉積[35],可見AMPK表達水平與肌間脂肪沉積量存在潛在負相關關系。
PPAR是核激素受體家族中的配體激活受體,與配體結合激活后,與視黃醇X受體(retinoid X receptor,RXR)形成異二聚體,形成的PPAR-RXR異二聚體與靶基因啟動子上游的PPAR反應元件結合,最終調節(jié)靶基因的轉錄。AMPK能提高PPARαmRNA的表達、降低PPARγmRNA的表達。其中PPARα是β-氧化的重要調節(jié)劑[36],表達提高可以促進脂質氧化[37]。有研究發(fā)現,PPARα缺失會導致小鼠的肥胖和脂肪肝加重[38],而PPARαmRNA表達水平升高,能減少肝臟中甘油三酯含量[39]。且激活PPARα能降低高脂飼養(yǎng)的大鼠肌肉脂肪含量[40]。PPARγ能誘導間充質干細胞分化為前脂肪細胞,參與脂質代謝相關基因的表達調控,促進脂肪酸合成。使用PPARγ激活劑后,能上調豬皮下和肌肉脂肪細胞中有關脂肪細胞分化的標志基因的表達[41]。Cui等[42]研究表明,PPARγmRNA表達量與萊蕪豬、魯萊黑豬以及大白豬肌內脂肪沉積均呈顯著正相關。活化的AMPK有抑制p38絲裂原活化蛋白激酶(MAPK)磷酸化的作用,p38 MAPK是細胞外信號轉導的樞紐,磷酸化的p38 MAPK在信號轉導通路中可以調控PPARγ的表達[43]。AMPK還能通過調節(jié)下游的乙酰輔酶A羧化酶(acetyl CoA carboxylase,ACC)和肉毒堿棕櫚酰轉移酶-1(carnitine palmitoyl transterase-1,CPT-1)來調控脂肪的代謝[35]。Merrill等[44]對SD大鼠的研究顯示,AMPK的表達增加會導致大鼠肌肉ACC活性減少,促進脂質氧化以及葡萄糖的攝取[44]。ACC是長鏈脂肪酸從頭合成的限速酶,催化脂肪酸合成的第一步反應,可以通過磷酸化和去磷酸化發(fā)揮調節(jié)脂肪酸合成的作用[45]。AMPK在Ser 79處直接磷酸化ACC,這種磷酸化會抑制ACC的活性[46],進而降低細胞內丙二酰輔酶A的含量,后者是CPT-1的生理抑制劑,因此解除了CPT-1的抑制作用[47]。CPT-1本身還受PPARα調節(jié),PPARα可以直接作用于CPT-1啟動子,在轉錄水平上影響CPT-1的表達[48],使CPT-1表達水平提高,促進脂肪酸的β-氧化[49]。CPT-1是脂肪酸β-氧化的關鍵限速酶,對脂肪氧化的整個過程都有重要的調控作用。CPT-1主要負責催化肉毒堿和乙酰輔酶A合成乙酰肉堿[50],將脂肪酸轉運到線粒體中進行氧化[51]。AMPK還可以抑制膽固醇調節(jié)元件結合蛋白-1c(sterol-regulatory element binding protein-1c,SREBP-1c)的表達。SREBP-1c是固醇原件結合蛋白家族的成員,能調節(jié)脂肪酸合成所需的基因的表達,促進細胞生成脂肪[52]。敲除C57BL/6J小鼠SREBP-1c基因,或SREBP-1c基因表達水平下降,都會導致脂肪含量顯著降低[53-54]。過表達SREBP-1c可以增加3T3-L1前體脂肪細胞中ACC等脂肪沉積相關基因的表達[55],還能與FAS靶基因的啟動子結合[56],FAS與肌肉和脂肪中脂肪酸含量密切相關,FAS表達量升高,體內甘油三酯含量增加,導致脂肪沉積[57]。AMPK還能通過下調CCAAT增強子結合蛋白α(CCAAT/enhancer binding protein α,C/EBPα)和脂肪酸結合蛋白(fatty acid-binding protein,FABP)表達水平,抑制脂肪合成[1,58]。有研究表明,C/EBPα能活化PPARγ,保持分化細胞的表型[59],從而調控脂肪細胞分化和脂肪沉積[9],但只有在PPARγ表達的前提下,過表達C/EBPα才能誘導成纖維細胞向脂肪細胞分化[60]。有研究表明,在3T3-L1脂肪細胞中,能通過下調C/EBPα和PPARγmRNA的表達水平來抑制脂肪生成[61]。存在于脂肪組織中的FABP稱為A-FABP或FABP4,能將脂肪酸從刷狀膜向光滑內質網位移,在脂類的代謝和運轉中起著重要作用,并能在脂肪細胞中沉積甘油三酯,從而改善肉質[62-63]。總而言之,大量研究都證明了AMPK處于脂肪調控通路的中心位置,其表達水平對各類脂肪沉積均有重要影響。圖1總結概述了AMPK信號通路在脂肪代謝中的調控作用。
3 營養(yǎng)因素對肌間脂肪的影響
近年來大量研究表明,營養(yǎng)能影響動物的表觀遺傳,通過改變飼糧能量、蛋白質水平,或者在飼糧中添加營養(yǎng)素來調控脂肪沉積,從而改善豬的胴體品質及肉品質,已成為畜牧學研究的熱點之一。
3.1 能量
從飲食中獲得的能量水平很大程度影響著機體的脂肪沉積量。豬在不同生長階段所需能量水平不同,通過調控飼糧能量水平可有效改善豬的胴體品質及肉品質。飼糧中消化能(DE)水平能通過調控以PPAR為中心的脂肪代謝通路來提高脂肪沉積量[64]。飼糧DE為16 MJ/kg時可以顯著降低豬PPARαmRNA在肌肉中的表達[65]。DE為13.85 MJ/kg相比于DE為12.75 MJ/kg,有提高60 kg“杜長大”三元雜交豬FABP基因表達量的趨勢,并且能顯著提高背脂厚度與肌內脂肪含量[66]。
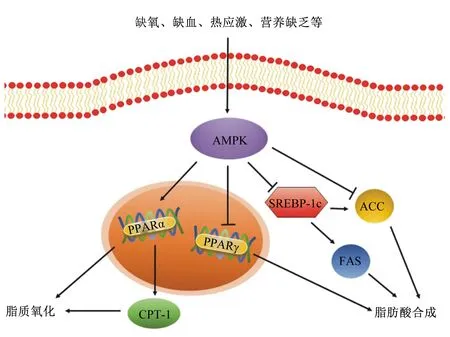
AMPK:腺苷酸活化蛋白激酶 AMP-activated protein kinase;PPAR:過氧化物酶體增殖物激活受體 peroxisome proliferators-activated receptor;CPT-1:肉毒堿棕櫚酰轉移酶-1 carnitine palmitoyl transterase-1;SREBP-1c:膽固醇調節(jié)元件結合蛋白-1c sterol-regulatory element binding protein-1c;ACC:乙酰輔酶A羧化酶 acetyl CoA carboxylase;FAS:脂肪酸合酶 fatty acid synthase;↓:促進 promote;⊥:抑制 inhibit。
3.2 蛋白質
蛋白質能調節(jié)豬的肌肉發(fā)育和脂肪沉積,因此飼糧的蛋白質水平對改善肉品質有重要作用[67]。降低飼糧蛋白質水平可以促進脂肪沉積。相比于粗蛋白質水平為18%的“杜長大”三元雜交豬,當飼糧粗蛋白質水平降低3個百分點時,其肌肉內ACCmRNA水平顯著提高,肌內脂肪含量也顯著增加[68]。低蛋白質組(16%粗蛋白質)的杜洛克豬和塔姆沃思豬肌間脂肪含量顯著大于對照組(20%粗蛋白質)[69],并且隨著理想蛋白質水平的提高,大白×長白雜交豬的大理石紋評分顯著下降,瘦肉率趨于增加等[70],這可能也與肌間脂肪的沉積存在關聯。還有研究表明,母豬在哺乳期限飼蛋白質,可以顯著降低后代肌纖維面積,從而影響肉品質[71]。鑒于當飼糧中蛋白質水平低于NRC(1998)標準不超過4個百分點時,其對豬的生長性能不會產生顯著影響[72],且有助于肌間脂肪沉積,提高豬肉品質,因此在生產過程中可以適當降低飼糧蛋白質水平。
3.3 氨基酸
氨基酸不僅能在機體內合成蛋白質,還可以作為信號分子調控脂肪沉積,改善豬肉風味和品質[73]。不同種類氨基酸對脂肪沉積的調控作用不同,大量研究表明賴氨酸、精氨酸、亮氨酸等氨基酸能通過調節(jié)脂肪代謝關鍵基因例如AMPK及其下游PPAR等基因的表達來影響脂肪沉積。當降低飼糧中賴氨酸與可消化能比值后,可以提高豬肌肉組織中PPARγ基因的表達水平[74],促進肌肉脂肪的沉積。雖然有研究報道補充精氨酸導致體脂降低8%[75],但在“杜長大”三元雜交豬的飼糧中添加1%的精氨酸能上調骨骼肌中PPARγmRNA表達水平[76],背最長肌脂肪含量也顯著提高[77],這證明精氨酸能在降低總脂肪的同時提高肌肉脂肪,對于提高肉品質有重要作用。飼糧中添加1%亮氨酸可以顯著提高“杜長大”三元雜交豬背最長肌脂肪含量[78]。但在高脂C57BL/6J小鼠中添加亮氨酸會顯著降低ACC在脂肪組織中的表達,而AMPK表達水平顯著升高,從而降低脂肪的沉積[79],這可能是由于亮氨酸在正常情況下能促進脂肪沉積,但在高脂情況下反而會促進脂肪的氧化分解代謝。
3.4 共軛亞油酸(conjugated linoleic acid,CLA)
CLA參與機體脂肪代謝,且對豬體況以及肉品質有積極影響,近年來開始被廣泛研究和報道。CLA的生物學作用之一是與脂肪沉積和養(yǎng)分代謝有關[80]。飼糧中額外添加CLA可以改善機體生長性能,提高肌間脂肪含量[81],減少皮下脂肪含量[82]。4%CLA可以提高長白×杜洛克雜交豬背最長肌肌間脂肪的PPARγ表達水平,促進肌間脂肪的沉積,有利于提高豬肉品質[83],這與Cordero等[84]和Zhong等[85]研究結果一致。
3.5 微量元素
在飲食中添加微量元素是改善豬肉品質的飲食干預之一,當在豬飼糧中添加有機硒后,能明顯降低脂肪氧化,提高豬肉品質[28]。高銅對于豬有顯著的促生長作用,但當飼糧中添加250 mg/kg銅之后,會降低背膘厚4.7%,對肉品質產生負面影響[86]。雖然有關微量元素調控豬脂肪沉積機理的報道較少,但大量研究表明微量元素對豬肉品質有重要調控作用,而豬的脂肪沉積也能影響肉品質,因此飼糧添加微量元素與機體脂肪沉積之間也必定存在著密不可分的關系。
3.6 植物提取物
除了幾大營養(yǎng)素對脂肪沉積有重要影響,大量研究還表明一些植物提取物具有降脂作用[87]。植物提取物中的生物活性成分能影響表觀遺傳,起到修飾基因的作用,從而改變DNA表達并調控脂肪沉積,例如紫檀芪能逆轉肥胖飲食引起的大鼠FAS基因DNA甲基化升高[88],緩解飲食引起的肥胖癥狀;姜黃素能顯著逆轉高脂大鼠PPARαDNA甲基化水平的升高[89],降低脂肪沉積;白藜蘆醇能顯著降低脂肪細胞中PPARγ表達水平,抑制脂肪的沉積[90]。植物提取物在降低總脂肪沉積量的同時,是否能提高肌間脂肪含量以及品質有待進一步研究。
4 小 結
肌間脂肪是肉品質的重要評判標準,如何降低皮下和腹部脂肪,同時提高肌間和肌內脂肪比例是改善胴體品質和肉品質的重點和難點所在,因此闡明脂肪沉積的分子機理顯得尤為關鍵。近年來研究表明,AMPK及其下游PPAR等相關基因的表達與脂肪沉積有著密切的關聯,而營養(yǎng)因素可以通過甲基化和去甲基化等途徑調控基因的表達水平,但具體的調控機制仍然不明確。從調控脂肪代謝的信號通路入手,明確不同組織中脂肪形成的分子機理,篩選獲得脂肪沉積的關鍵調控位點,對精準調控脂肪沉積改善肉品質至關重要。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年6期)2019-10-08 08:55:48
人大建設(2019年12期)2019-05-21 02:55:32
雜文月刊(2018年21期)2019-01-05 05:55:28
人大建設(2017年6期)2017-09-26 11:50:44
學苑創(chuàng)造·A版(2015年11期)2016-01-14 09:03:27
俄羅斯問題研究(2012年1期)2012-03-25 09:54:45
中國火炬(2010年12期)2010-07-25 13:26:22
中國火炬(2010年8期)2010-07-25 11:34:30
