4 個紫花苜蓿品種對低溫脅迫的生理響應及抗寒性評價
2021-05-12 14:09:06趙一航孟令東張曉萌王麗娜劉皓玥畢琳琳劉宏亮殷秀杰
草業科學 2021年4期
趙一航,孟令東,張曉萌,王麗娜,劉皓玥,畢琳琳,劉宏亮,殷秀杰
(1. 東北農業大學動物科學技術學院,黑龍江 哈爾濱 150030;2. 雀巢奶牛養殖培訓中心, 黑龍江 哈爾濱 150110;3. 雀巢谷物能力中心, 黑龍江 哈爾濱 150110)
紫花苜蓿 (Medicago sativa)是豆科苜蓿屬多年生草本植物,具有產量高、適應性強、再生能力強等特點[1]。‘肇東’、‘龍牧801’、‘東農1 號’和‘敖漢’4 個品種在我國北方被廣泛種植[2-3],而冬季低溫是制約我國北方地區苜蓿生產的主要因素,部分地區出現苜蓿凍害減產等問題,遭受巨大經濟損失[4]。低溫會影響紫花苜蓿的正常生長發育,不僅會導致植物物質代謝失調,甚至會導致植株死亡[5]。因此,提高苜蓿品種抗寒性、選育抗寒苜蓿品種對我國北方地區發展苜蓿產業具有重要意義。
植物在低溫脅迫下細胞膜被破壞,膜透性增加,相對電導率可以反映植物生物膜的透性,抗寒能力不同的植物電導率的變化會有顯著差異[6]。楊國柱等[7]通過測量低溫脅迫下12 個不同品系紫花苜蓿的相對電導率來比較其抗寒性。低溫脅迫下植物體內活性氧不斷積累最終導致膜脂過氧化產生丙二醛[8-10],植物通過增加可溶性糖和游離脯氨酸[11-16]等滲透調節物質的含量,以及增強保護酶活性,如過氧化物酶和超氧化物歧化酶[17-19],提高植物的抗寒能力。 因此可以通過測定低溫脅迫下植物相對電導率、丙二醛含量、可溶性糖含量、游離脯氨酸含量、過氧化物酶含量和超氧化物歧化酶的含量比較不同植物或者品種之間抗寒性的差異。葉片是植物代謝的主要器官,可以依據植物葉片的解剖結構特征和生長狀況判斷植物對其生長環境的適應性[20]。在低溫脅迫下,植物內部的解剖結構會發生不同程度的變化。研究表明,細胞結構越緊密的苜蓿品種抗寒性越強[21],抗寒性弱的苜蓿品種的葉片組織厚度大于抗寒性強的苜蓿品種[13]。
目前對不同品種苜蓿低溫脅迫下生理響應和抗寒性的研究較多[21-25],如通過對不同根型根系紫花苜蓿生理生化指標進行測定,從而對其進行抗寒性評價[23];通過8 個品種紫花苜蓿的相對電導率和根莖活力計算半致死溫度,從而對8 種苜蓿的抗寒性進行評價[24]。也有部分關于非生物脅迫下苜蓿葉片解剖結構的研究[21]。但是對不同品種苜蓿低溫脅迫下的生理生化各指標進行系統性的分析,并結合其葉片解剖結構對抗寒性進行評價的相關研究較少[13]。因此,本研究通過對低溫脅迫下4 種苜蓿生理生化指標和葉片解剖結構的觀測,綜合比較了4 種苜蓿的抗寒性,以期為我國北方地區篩選抗寒性強的紫花苜蓿品種提供理論依據。
1 材料與方法
1.1 試驗材料
試驗材料來源如表1 所列。
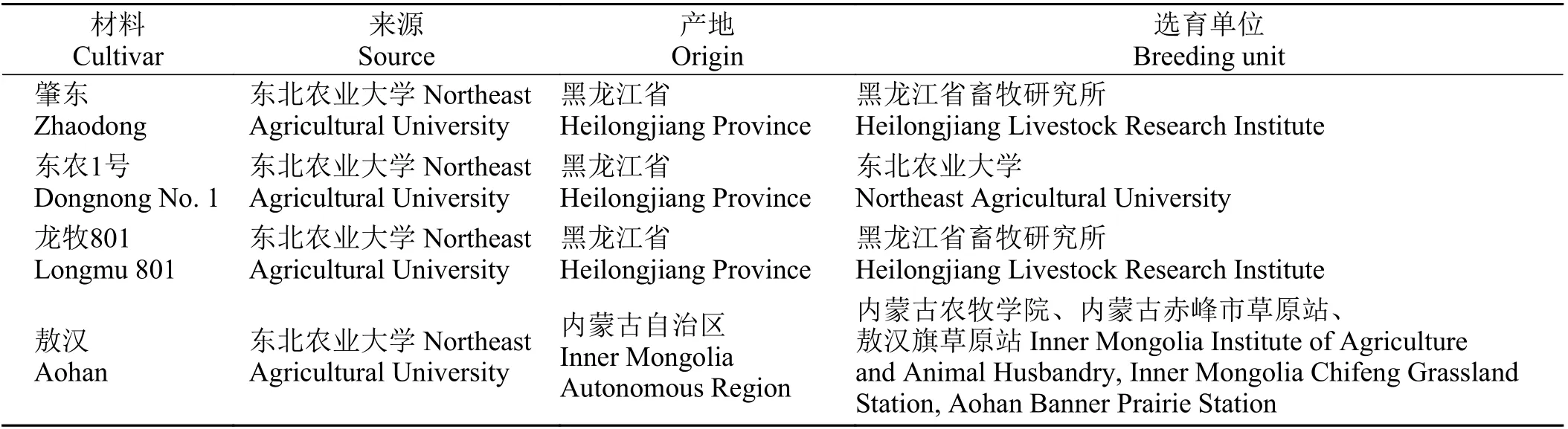
表 1 供試材料Table 1 Cultivars examined
1.2 試驗設計
選取大小均勻一致的4 種苜蓿種子播種于穴盤中(穴盤規格為540 mm × 280 mm,50 穴),每個穴內播種5~6 粒種子,將播種好的穴盤放置于培養箱中,每天按時澆水管理。培養條件如下:光暗周期為12 h/12 h,溫度為白天(20 ± 1) ℃/夜晚(17 ± 1) ℃,空氣相對濕度為65%。培養60 d 后,分別選取生長狀況良好且均一的同種苜蓿植株進行低溫脅迫處理,以20 ℃作為對照組 (CK),以溫度梯度4、0、-4 和-8 ℃進行低溫脅迫,每個溫度梯度的脅迫時間為24 h,對植株地上部分取樣并用錫紙包好,作為待測試驗樣品,放入-80 ℃冰箱中保存,每個樣品3 次重復[24]。
分別取4、0、-4 和-8 ℃處理組的苜蓿葉片和對照組的苜蓿葉片,切成5 mm × 5 mm 小塊,立即放入酒精、醋酸、福爾馬林混合固定液中固定24 h,采用常規石蠟切片法,經過取樣、固定、脫水、透明、浸蠟、包埋、切片(厚度8 μm)、粘片、去蠟、染色(蘇木精染色)、中性樹膠封片等步驟永久制片[25]。
1.3 測定指標與方法
先將所取樣品進行液氮粗磨,再通過研磨機細磨后進行各指標測定。相對電導率采用電導法測定[26],丙二醛含量采用硫代巴比妥酸比色法測定[27],可溶性糖含量采用蒽酮比色法測定[27],游離脯氨酸含量采用酸性茚三酮法測定[27],超氧化物歧化酶活性采用氮藍四唑法測定[27],過氧化物酶活性采用愈創木酚比色法測定[28]。
1.4 解剖結構測定
用顯微鏡觀察葉片橫切面的形態結構和變化情況,測量并記錄葉片厚度(μm)、柵欄組織厚度(μm)、海綿組織厚度(μm)、維管束直徑(μm)等結構。
柵海比 = 柵欄組織厚度/海綿組織厚度[29];
葉片組織結構緊密度 = (柵欄組織厚度/葉片厚度) × 100%;
葉片組織結構疏松度 = (海綿組織厚度/葉片厚度) × 100%。
1.5 統計方法
所有抗寒性生理指標和解剖結構數據應用SPSS 19.1 軟件進行方差顯著性分析,應用Excel 2013 對試驗數據進行數據處理和圖表制作。
1.6 隸屬函數分析方法
用于分析的隸屬函數值[ X1, X2]計算方程如下:
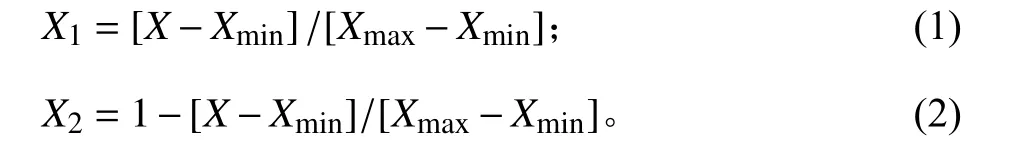
式中: X 為某種苜蓿某一指標的測定值; Xmax為4 種苜蓿此指標的最大值; Xmin為4 種苜蓿此指標的最小值。若所測指標與植物的抗寒性呈正相關關系,則采用式(1)計算隸屬值,負相關用式(2)。累加4 種苜蓿各指標的具體隸屬值,并求出平均值后進行比較,平均值越大則表明植物的抗寒性越強[30-31]。
2 結果與分析
2.1 相對電導率
在4 ℃處理下4 種苜蓿的相對電導率沒有顯著差異(P > 0.05) (圖1)。而-4 和-8 ℃處理下4 種苜蓿的相對電導率相比對照組均有大幅提升。與對照組相比,-4 ℃處理下4 種苜蓿的相對電導率上升幅度為‘敖漢’ > ‘肇東’ > ‘龍牧801’ > ‘東農1 號’;-8 ℃處理下4 種苜蓿的相對電導率上升幅度為‘敖漢’ >‘龍牧801’ > ‘東農1 號’ > ‘肇東’。
2.2 丙二醛含量
除4 ℃處理下‘肇東’苜蓿的丙二醛含量低于對照外,其他處理下4 種紫花苜蓿的丙二醛含量均高于對照(圖1)。隨著溫度的降低,‘東農1 號’和‘敖漢’的丙二醛含量呈先上升后下降的趨勢,‘肇東’則呈上升趨勢。與對照組相比,‘肇東’和‘龍牧801’苜蓿在-8 ℃處理下丙二醛含量最高,分別上升52.96%和59.90%。‘東農1 號’苜蓿在-4 ℃處理下丙二醛含量最高,與對照組相比上升幅度為66.12%;‘敖漢’在0 ℃處理下丙二醛含量最高,與對照組相比上升幅度為126.68%。4 種苜蓿受到低溫脅迫后丙二醛含量總體表現為‘敖漢’ > ‘東農1 號’ > ‘龍牧801’ >‘肇東’。
2.3 可溶性糖含量
低溫處理下4 種苜蓿的可溶性糖含量與對照組相比有所上升,-8 ℃處理下‘東農1 號’和‘敖漢’的可溶性糖含量與對照組相比略有下降(圖1)。隨著溫度的降低,‘肇東’、‘龍牧801’和‘東農1 號’的可溶性糖含量呈先上升后下降的趨勢,并且在0 ℃處理下最高。‘敖漢’在4 ℃處理下可溶性糖含量最高。與對照相比,‘肇東’和‘龍牧801’的可溶性糖含量總體上升最為明顯,‘敖漢’無明顯變化。
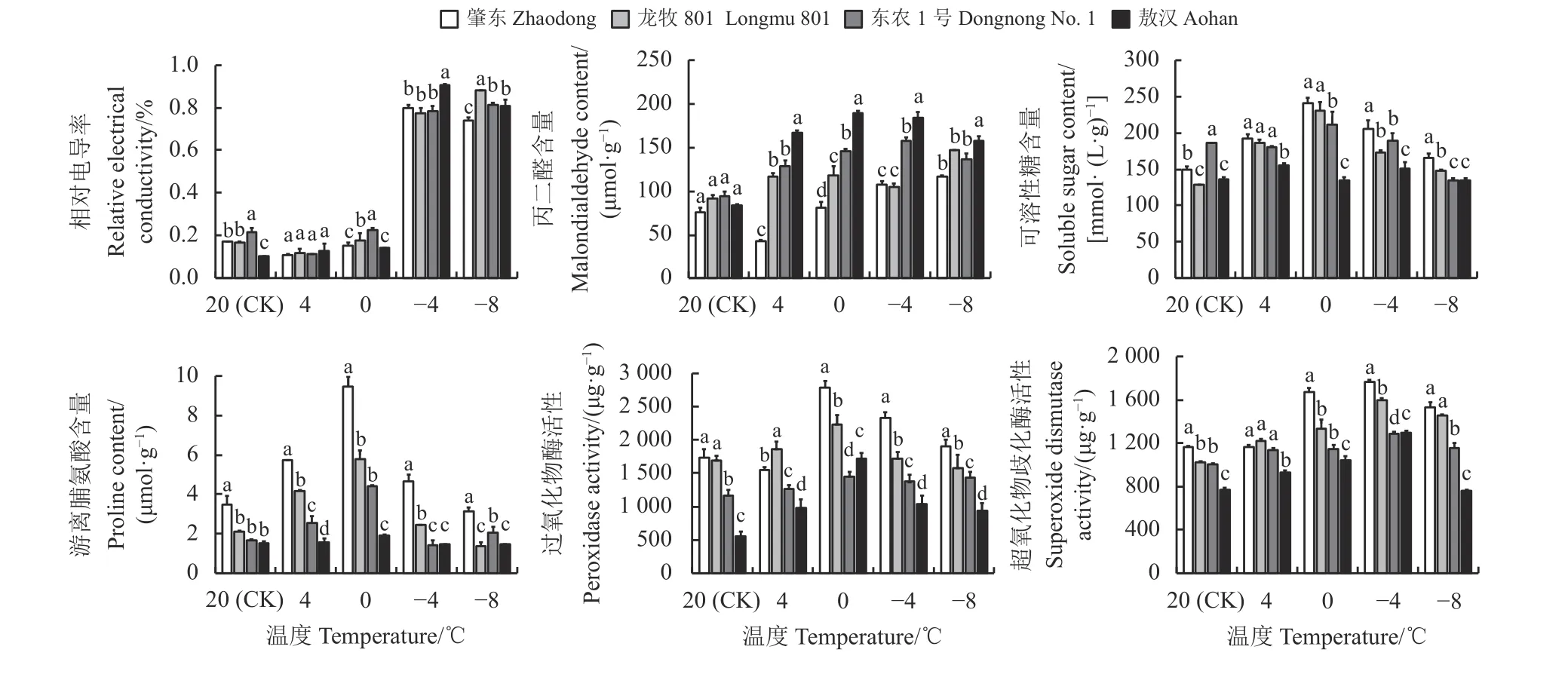
圖 1 低溫脅迫下紫花苜蓿的生理生化指標變化Figure 1 Physiological and biochemical indexes of alfalfa under low temperature treatments
2.4 游離脯氨酸含量
低溫處理下4 種苜蓿的游離脯氨酸含量與對照組相比均有所提升,并且隨著溫度降低,4 種苜蓿的游離脯氨酸含量呈先上升后下降的趨勢,4 種苜蓿的脯氨酸含量均在0 ℃處理下最高(圖1)。與對照組相比,‘肇東’的游離脯氨酸含量上升最為明顯,上升幅度達到179.3%;‘敖漢’的變化較小,上升幅度僅為23.7%。4 種苜蓿受到低溫脅迫后游離脯氨酸含量總體上升幅度為‘肇東’ > ‘龍牧801’ > ‘東農1 號’ >‘敖漢’。
2.5 過氧化物酶活性
在0、-4、-8 ℃處理下4 種苜蓿的過氧化物酶活性與對照組相比大幅提高,而4 ℃處理下4 種苜蓿的過氧化物酶活性與對照組相比變化不明顯(圖1)。4 種苜蓿在0 ℃處理下過氧化物酶活性最高,-4 和-8 ℃處理下的過氧化物酶活性比0 ℃處理下低。總體來看,低溫處理下‘肇東’的過氧化物酶活性上升幅度最高,‘東農1 號’最低。
2.6 超氧化物歧化酶活性
低溫處理下4 種苜蓿的超氧化物歧化酶活性與對照組相比有所上升(圖1)。隨著處理溫度的降低,4 種苜蓿的超氧化物歧化酶活性均呈現先上升后下降的趨勢,4 種苜蓿均在-4 ℃處理下酶活性最高,隨后活性變弱,-8 ℃處理下酶活性下降明顯,其中敖漢下降幅度最大,‘肇東’和‘龍牧801’的酶活性下降幅度較小。總體來看,低溫處理下超氧化物歧化酶活性的上升幅度為‘肇東’ > ‘龍牧801’ > ‘敖漢’ >‘東農1 號’。
2.7 隸屬函數綜合分析
僅憑單一指標難以對植物抗寒性進行評價,因此通過隸屬函數綜合分析4 種苜蓿的6 項生理生化指標,從而對抗寒性進行綜合評價。結果顯示(表2),‘肇東’和‘龍牧801’的抗寒能力較強,‘敖漢’的抗寒能力最弱,4 種苜蓿的抗寒性為‘肇東’ > ‘龍牧801’ >‘東農1 號’ > ‘敖漢’。
2.8 低溫脅迫對苜蓿葉片解剖結構的影響
正常生長下4 種苜蓿的葉片厚度在96.14~129.9 μm,其中‘肇東’和‘龍牧801’差異不顯著(P > 0.05)(表3)。‘敖漢’與‘東農1 號’差異不顯著,但顯著大于‘肇東’和龍牧801 的葉片厚度 (P < 0.05)。4 種苜蓿的葉片厚度為‘敖漢’ > ‘東農1 號’ > ‘龍牧801’ >‘肇東’。4 種苜蓿的柵欄組織厚度在47.04~53.99 μm,‘敖漢’的柵欄組織厚度最大,顯著高于‘龍牧801’和‘東農1 號’,后兩者差異不顯著,‘肇東’顯著低于其他3 種苜蓿(P < 0.05)。4 種苜蓿的海綿組織厚度在37.41~65.70 μm,‘肇東’和‘龍牧801’的海綿組織厚度顯著小于‘敖漢’。4 種苜蓿的維管束直徑在62.36~78.59 μm,其中‘肇東’和‘東農1 號’差異不顯著,‘龍牧801’與‘敖漢’差異不顯著。4 種苜蓿的柵欄組織和海綿組織之比在1.04~1.31,其中‘肇東’和‘龍牧801’柵海比大于‘東農1 號’和‘敖漢’,并且‘肇東’和‘龍牧801’與‘東農1 號’和‘敖漢’之間差異顯著(表4)。4種苜蓿的細胞結構緊密度在41.56%~52.79%,由大到小依次為‘龍牧801’ > ‘肇東’ > ‘東農1 號’ > ‘敖漢’。
4 種苜蓿在不同溫度處理下,葉片厚度、柵欄組織厚度和海綿組織厚度與對照組相比均有所減小(圖2)。其中‘肇東’的葉片厚度與對照組相比減小幅度為27.50%;‘敖漢’的減小幅度為39.47%;‘龍牧801’的減小幅度為12.05%;‘東農1 號’的減小幅度為34.61%。變化幅度為‘敖漢’ > ‘東農1 號’ > ‘肇東’ > ‘龍牧801’。
3 討論
低溫脅迫后植物細胞膜被破壞,膜透性增加,相對電導率可以反映植物生物膜的透性[32-33]。對照組中,4 種苜蓿的相對電導率存在顯著差異,推測是由于不同品種的基因型不同而造成的。4 種苜蓿的相對電導率與對照組相比,‘肇東’的增幅最小,‘敖漢’的增幅最大。說明‘敖漢’在受到低溫脅迫后其細胞膜的受損程度最嚴重。隨著脅迫溫度的下降,4 種苜蓿的相對電導率均呈現先平穩、再上升、最后平穩的趨勢,4 種苜蓿的相對電導率均在-4 ℃時急劇上升,相對電導率呈“S”型變化趨勢,這一結果與曾憲海等[34]應用電導率法及Logistic 方程評價引種海南儋州油棕(Elaeis guineensis)新品種抗寒性研究的結論相一致。研究表明,當植物受到低溫脅迫時體內丙二醛含量均有不同程度的上升,可以反映植物細胞膜的過氧化程度[35]。本研究結果表明,低溫脅迫下4 種苜蓿的丙二醛含量為‘敖漢’ > ‘龍牧801’ > ‘東農1 號’ > ‘肇東’。這說明4 種苜蓿中,‘肇東’的抗氧化能力最強,‘敖漢’的抗氧化能力最弱,‘龍牧801’和‘東農1 號’的抗氧化能力介于兩者之間。
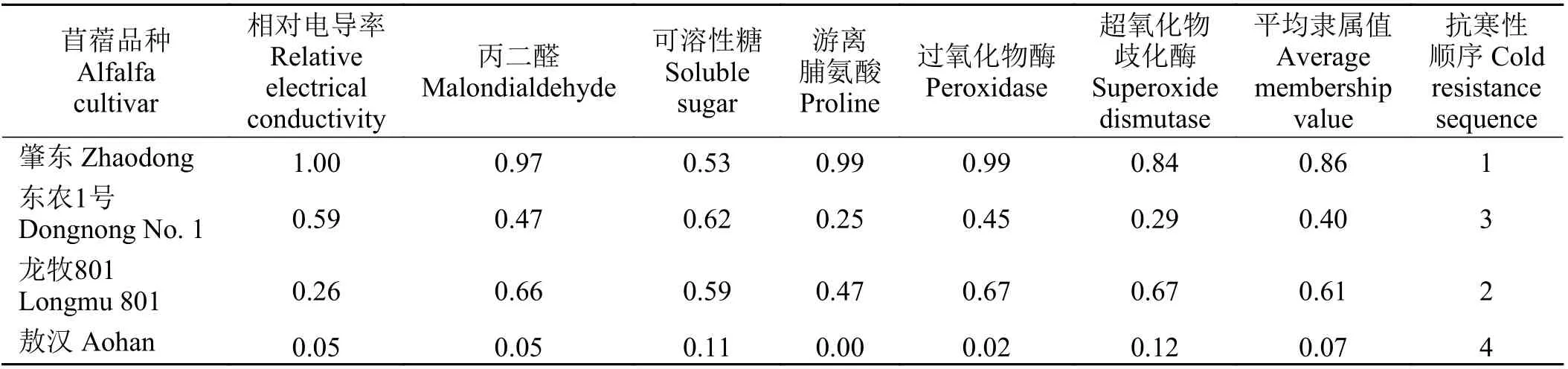
表 2 不同苜蓿品種的隸屬函數值Table 2 Subordinate function values of different alfalfa cultivars
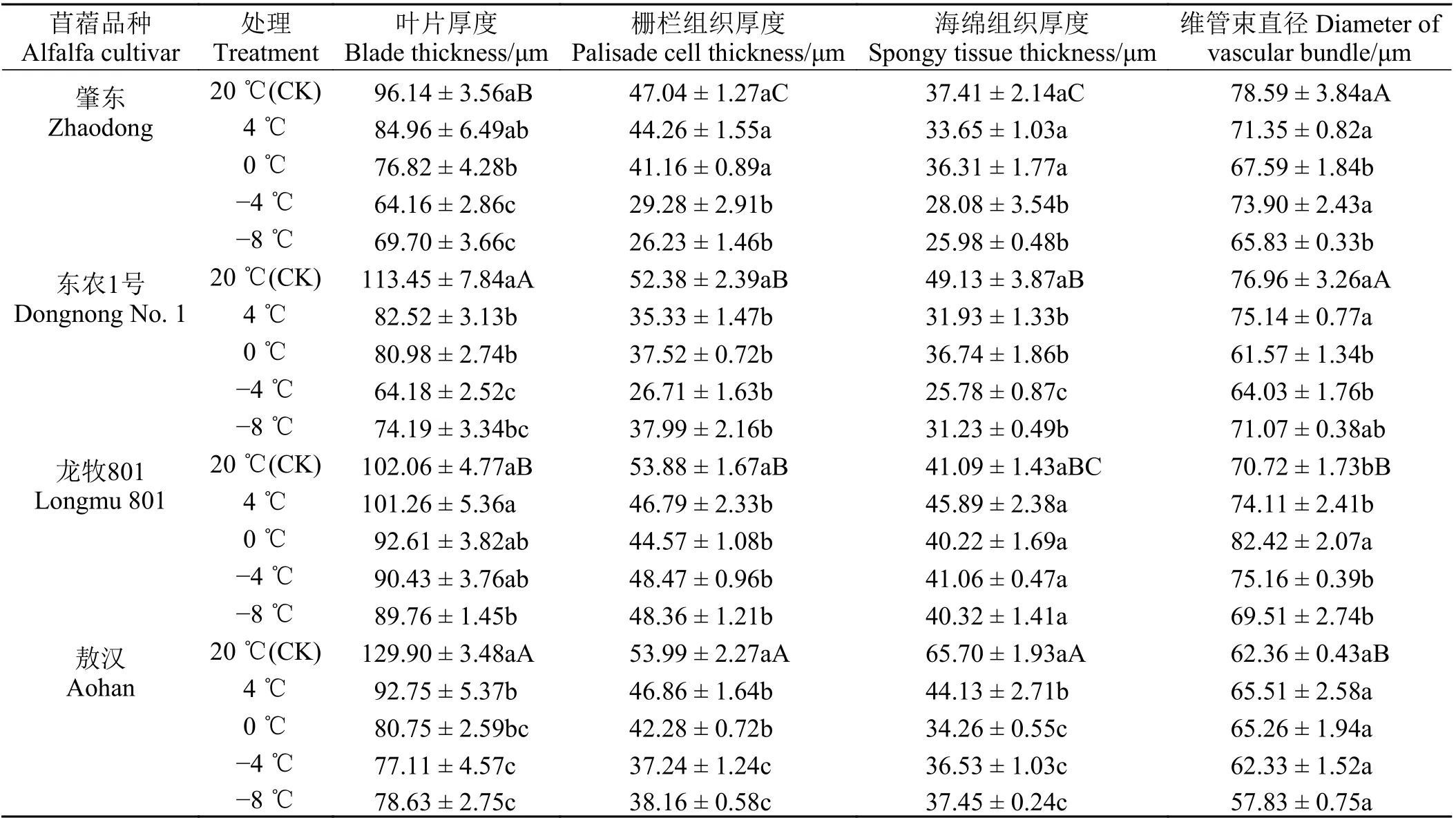
表 3 低溫脅迫下葉片解剖結構的比較Table 3 Comparison of leaf anatomical structure under low temperature treatments
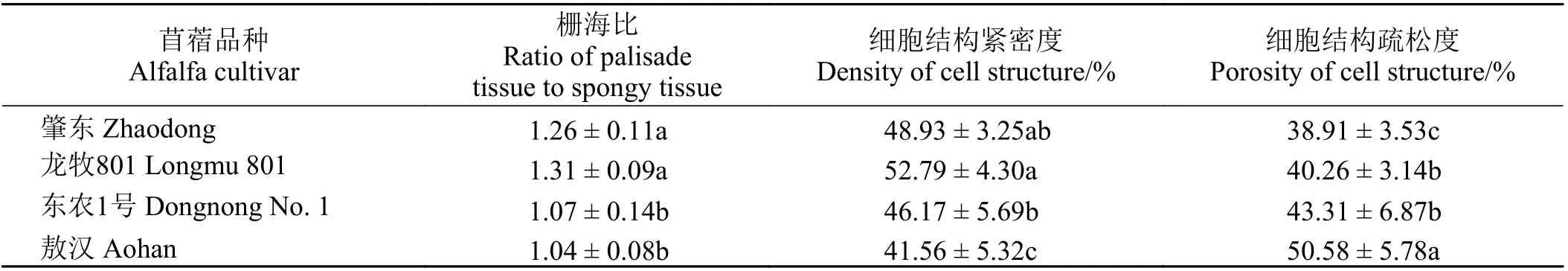
表 4 苜蓿葉片解剖結構的參數測定結果Table 4 Determination of anatomical structure parameters of alfalfa leaves
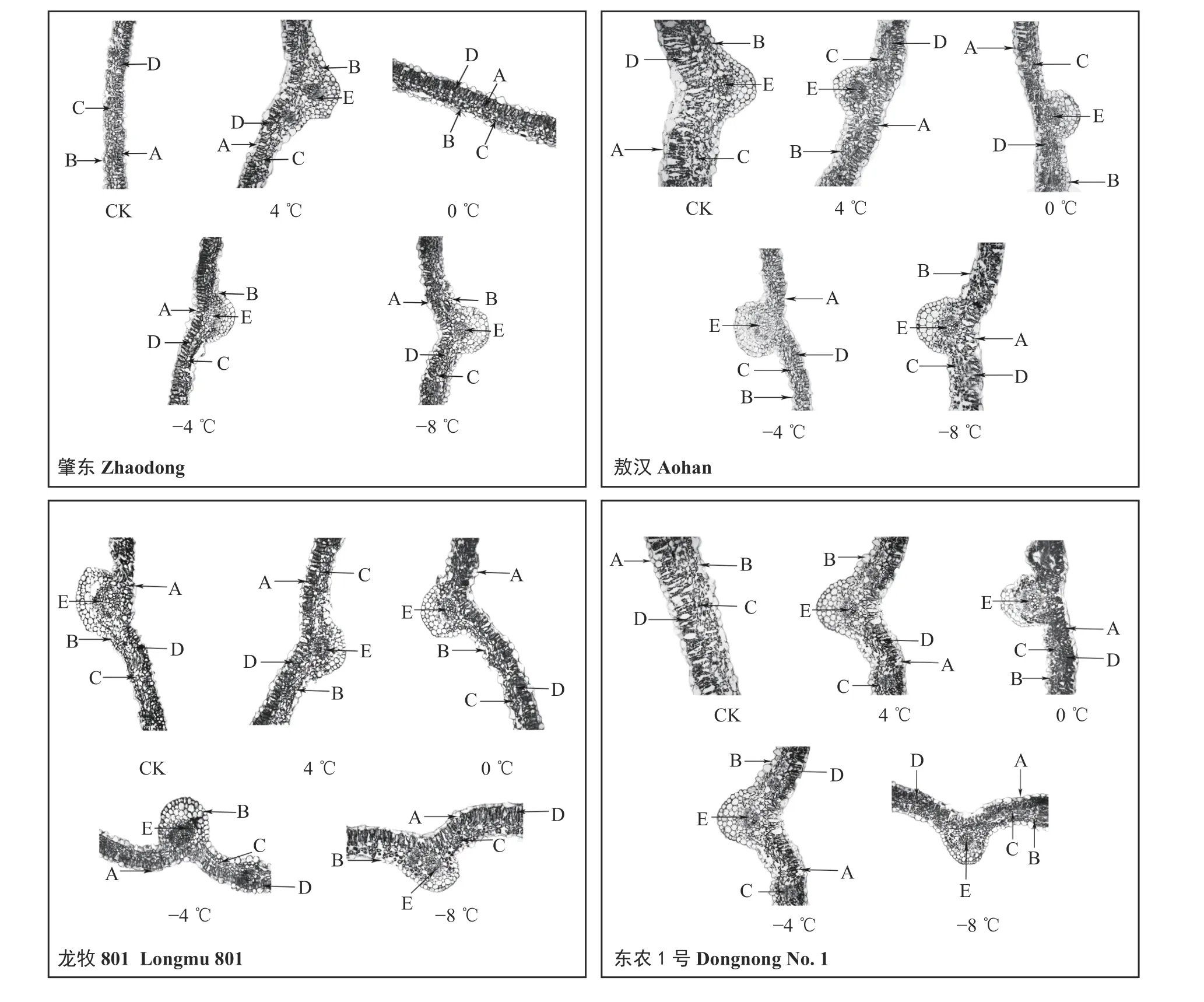
圖 2 4 種紫花苜蓿葉片的解剖結構Figure 2 Anatomical structure of leaves of four alfalfa cultivars
植物可以通過積累可溶性糖提高自身保水能力,低溫脅迫下植物的可溶性糖含量越高,植物的抗寒能力就越強[36]。對照組中,‘東農1 號’和‘肇東’的可溶性糖含量較高;在低溫脅迫下,‘肇東’和‘龍牧801’的可溶性糖含量上升且高于‘東農1 號’和‘敖漢’的可溶性糖含量,說明‘肇東’和‘龍牧801’在受到低溫脅迫時,能夠調節自身生理機能,使可溶性糖合成能力提高,從而更好地抵御低溫。低溫脅迫下4 種苜蓿的可溶性糖含量均呈先上升后下降的趨勢,并且低溫脅迫下抗寒性強的品種,其可溶性糖含量的上升幅度較大,這一結果與黑龍江省主要栽培葡萄(Vitis vinifera)品種抗寒性研究的結論[37]相一致。游離脯氨酸是植物體內一種重要的滲透調節物質,游離脯氨酸含量是衡量苜蓿抗寒性的一個重要指標[38]。隨著脅迫溫度降低,4 種苜蓿的游離脯氨酸含量在0 ℃處理下達到最高,隨后含量降低。游離脯氨酸含量上升幅度為‘肇東’ > ‘龍牧801’ >‘東農1 號’ > ‘敖漢’。4 種苜蓿中抗寒性強的品種低溫脅迫下游離脯氨酸含量上升幅度大于抗寒性弱的品種,這一結果與張榮華等[39]關于脯氨酸含量對苜蓿抗寒性影響研究的結論相一致。
超氧化物歧化酶(SOD)和過氧化物酶(POD)作為抗氧化防御系統可以清除氧自由基,對保護植物正常生理功能和提升植物抵御逆境的能力具有重要作用[40]。低溫脅迫下,4 種苜蓿SOD 和POD 活性的變化規律較為一致,隨溫度降低SOD 和POD 活性均呈現先上升后下降的趨勢,上升幅度為‘肇東’ >‘龍牧801’ > ‘東農1 號’ > ‘敖漢’;SOD 和POD 活性上升達到最高后活性下降,這可能是因為處理溫度低于苜蓿所能承受的最低溫度,導致酶活性逐漸降低。低溫脅迫下植物體內的SOD 和POD 含量均有所上升,并且抗寒性越強的植物酶活性越強,上升的幅度越大,但低溫脅迫過于嚴重時,植物體內的酶活性降低。這與梁坤倫等[41]和鐘海霞等[42]在垂穗披堿草(Elymus nutans)和葡萄的抗寒性研究中得出的結果一致,輕度低溫脅迫會使植物體內SOD和POD 活性上升,但重度低溫脅迫下,植物體內SOD和POD 活性會降低。
4 種苜蓿的細胞結構緊密度呈現顯著差異,并且抗寒性強的品種細胞結構緊密度大于抗寒性弱的品種;但是細胞結構緊密度是否與其抗寒性有關仍需進一步研究。通過比較4 種苜蓿的維管束直徑發現,‘肇東’和‘東農1 號’之間差異不顯著,‘龍牧801’與‘敖漢’之間差異不顯著;但是不同處理溫度下4 種苜蓿與對照組相比,葉片厚度、柵欄組織厚度、海綿組織厚度與對照組相比均有所減小,并且呈顯著差異。4 種苜蓿在不同處理溫度下維管束直徑均無明顯變化,且與對照組相比差異不顯著,這說明維管束直徑不能作為比較抗寒性的指標。通過研究4 種苜蓿對照組的解剖結構發現,抗寒性弱的品種的葉片厚度、柵欄組織厚度、海綿組織厚度均大于抗寒性強的品種。
4 結論
低溫脅迫下,4 種苜蓿中‘敖漢’的相對電導率和丙二醛含量上升幅度最大,細胞膜損傷最為嚴重。低溫脅迫下,4 種苜蓿的可溶性糖含量、游離脯氨酸含量、過氧化物酶和超氧化物歧化酶活性與對照相比有所上升,其中‘肇東’各個指標的上升幅度均最大,表明4 種苜蓿中‘肇東’對低溫的抗逆性更強。4種苜蓿的抗寒性為‘肇東’ > ‘龍牧801’ > ‘東農1 號’ >‘敖漢’。對照組中4 種苜蓿的葉片厚度為‘敖漢’ >‘東農1 號’ > ‘龍牧801’ > ‘肇東’。抗寒性弱的苜蓿品種葉片厚度較大,抗寒性強的品種葉片厚度較小。抗寒性強的品種柵欄組織厚度、海綿組織厚度均小于抗寒性弱的品種,其細胞結構緊密度和柵海比均大于抗寒性較弱的品種。
