玉米干旱響應轉錄因子的研究進展
2021-05-13 10:52:50周洋洋劉書伯于豐碩劉桐羽劉夢彤關淑艷馬義勇
山東農業科學 2021年4期
關鍵詞:植物
周洋洋,劉書伯,于豐碩,劉桐羽,劉夢彤,關淑艷,馬義勇
(1.吉林農業大學生命科學學院,吉林 長春 130118;2.吉林農業大學農學院,吉林 長春 130118)
玉米(Zea mays L.)不僅是被廣泛種植的糧食作物,也是重要的飼料作物和工業原料作物。干旱會導致葉片萎縮卷曲并造成細胞損傷等,進而對產量造成嚴重影響,是限制我國玉米生產發展的主要因素[1]。轉錄因子(TF)是植物響應脅迫的調控因子,目的基因啟動子通過與轉錄因子相互作用對基因的表達進行調控;轉錄因子通過調控一系列的胞內信號轉導過程,如促進或抑制目的基因功能,使植物對非生物脅迫產生響應[2]。近幾年在作物中已鑒定出多種干旱響應轉錄因子家族,其中較為常見的是MYB、WRKY、bZIP、bHLH、ERF等[3],然而對于玉米干旱響應轉錄因子家族的系統分析還未見報道。本文主要闡述玉米干旱響應轉錄因子家族的種類、結構、生物學功能及作用機制,并從轉錄因子順式作用元件、轉錄因子基因表達以及轉錄因子蛋白間的相互作用方面論述轉錄因子在干旱調控中的作用機制,以期為進一步培育高產、耐旱玉米新品種提供理論基礎及科學啟示。
1 玉米干旱響應轉錄因子家族的種類與結構
利用plantTFDB(http://planttfdb.cbi.pku.edu.cn/)在線檢測到玉米中共有56個轉錄因子家族,經過比對分析玉米的轉錄組數據得到轉錄因子基因2 270個[4]。通過對轉錄因子基因進行篩選、qRT-PCR檢測并結合SNP及農藝性狀鑒定,最終發現與氣孔關閉、根系發育及活性氧清除等干旱響應過程有關的轉錄因子基因主要分布在15個轉錄因子家族[5-7]。通過生物信息學分析并對玉米轉錄組數據進行篩選統計,進一步明確了這15個玉米干旱響應轉錄因子家族分別為AP2、C2H2、MYB、NAC、WRKY、ERF、GRAS、HSF、ARF、HD-ZIP、NF-YB、bZIP、GATA、NF-YA和bHLH[8]。
在干旱脅迫下,轉錄因子的結構域對其執行特定功能起著決定性的作用,并且不同的結構域具有不同的結合方式。例如,Basic-leucine zipper(bZIP)是一類植物中廣泛存在的轉錄因子,其N末端通常與DNA結合,C末端通常與富含亮氨酸的基序結合形成二聚體;bZIP家族通過與啟動子區域的ABRE(ABA響應元件)結合,通過ABA途徑發揮作用[9,10]。WRKY家族有一個保守的WRKY結構域,該結構域與W-box(60個氨基酸的保守片段)結合以調控基因的表達[11]。表1為玉米干旱響應轉錄因子家族的種類及其結構。

表1 玉米干旱響應轉錄因子種類與結構
2 玉米干旱響應轉錄因子家族的生物學功能
Shikha等[22]研究發現玉米15個干旱響應轉錄因子家族中63.30%的基因與生物學功能(BP)相關,40.53%的基因與細胞定位(CC)相關,88.51%的基因與分子功能(MF)相關;值得注意的是,其中33%的基因與干旱響應過程密切相關,這些基因參與了ABA 信號轉導、活性氧(ROS)清除、光合作用、氣孔發育以及蔗糖代謝過程。轉ZmbZIP72基因玉米植株在干旱脅迫下該基因表達呈上調趨勢,使得植株中ABA含量有所增加,進而提高植株抗氧化能力、水分利用效率,增強植株對干旱脅迫的耐受性[23]。玉米植株中ZmNF-YA13基因通過與ZmSOD基因相互作用,進一步激活了氧自由基代謝過程,使植株的抗旱性有所提高[12]。干旱脅迫下玉米組織中AhNAC2基因和AhNAC3基因均上調表達,但是兩者表達方式存在一定差異[24]。干旱脅迫下bHLH轉錄因子家族參與調節氣孔發育過程及組織分化過程,OsbHLH148基因參與茉莉酸信號途徑,進而提高了植株的抗旱能力[13]。干旱脅迫下PmNAC基因對植株的生長發育進行適應性調節[25];WRKY轉錄因子能夠促進種子萌發和生長素的合成,在植物對干旱脅迫的響應過程中發揮著關鍵的主導作用[26]。同一轉錄因子基因因調節機制不同而具有不同功能,如參與調節植物發育過程、信號轉導過程等[14]。表2為近幾年玉米干旱響應轉錄因子家族的生物學功能研究情況。

表2 玉米干旱響應轉錄因子家族生物學功能
3 玉米干旱響應轉錄因子家族的作用機制
3.1 玉米干旱響應轉錄因子的順式作用元件
轉錄因子通過與位于啟動子區的順式作用元件(CREs)結合,調控基因的表達。Mishra等[31]利用PlantPAN(http://plantpan.itps.ncku.edu.tw/)數據庫,對玉米干旱響應的轉錄因子、順式作用元件以及上游啟動子進行篩選,共鑒定出42種CREs,其中28種是所有轉錄因子共有,這些CREs通過調控特定的起始時間及表達程度發揮作用,進而調控基因的轉錄。與干旱響應有關的CREs,如DRE1、DRE/CRT和DRECRTCOREAT等,通過與轉錄因子結合來提高植物對干旱脅迫的響應能力[32]。因此了解玉米干旱響應轉錄因子與不同順式作用元件間的作用機制(圖1),對研究玉米抗旱調節機制具有重要意義。
玉米通過轉錄因子與順式元件如DREB、ABRE/ABF、MYB、WRKY/ERF、DRE/CRT、ABRE等的結合,可進一步調控玉米的抗旱機制[33]。例如轉錄因子NF-YA通過調控有絲分裂相關基因,參與不同的抗旱調節過程,如氣孔大小調節過程、活性氧(ROS)消除過程[34,35];通過與ARF作用,對植物的發育過程進行調節并抑制側枝生長[27];NAC轉錄因子通過與干旱應答順式作用元件MBS結合對植物細胞滲透壓進行調節[36];NF-YB轉錄因子與CBF順式元件結合,可有效提高光合速率[28]。ABA非依賴途徑或ABA依賴途徑中存在的轉錄因子與順式元件相互作用,可以對轉錄過程進行調節,從而激活干旱脅迫響應基因,使氣孔關閉,進而阻止蒸騰作用導致的過度失水[37]。玉米HD-ZIP轉錄因子在干旱脅迫下通過與順式作用元件GARE作用應對干旱脅迫[38]。干旱脅迫下植物中的NAC轉錄因子與順式作用元件CBF結合,對根系生長進行調節以提高植物水分的利用率[39]。綜上,通過了解玉米干旱響應轉錄因子與順式作用元件間的作用機制,可為高效、精準培育耐旱玉米新品種提供理論依據。
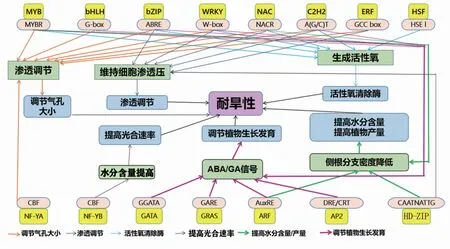
圖1 轉錄因子與順式作用元件的作用機制
3.2 玉米干旱響應轉錄因子基因表達
Akhtar等[40]證實了干旱脅迫下植物轉錄因子基因的作用,如過量表達DREB1/CBF提高了玉米、花生和小麥的耐旱性。轉錄因子基因的表達具有組織特異性,大多數干旱響應轉錄因子基因在根部表達,分泌植物激素并激活ABA途徑,從而使根部產生ABA,然后通過一系列信號轉導過程將信號從根部傳遞到地上部分[41]。當玉米植株處于干旱條件下,位于根部的轉錄因子使目的基因表達呈上調趨勢[42],通過調節MYB、bZIP、WRKY和NAC相關基因的信號轉導過程,并發生一系列的級聯反應,誘導植物產生一系列調節干旱脅迫的生理活動,如氣孔關閉、根部伸長等,以提高其抵御干旱脅迫的能力[43]。Xing等[15]的研究表明,干旱脅迫下,玉米中的ARF轉錄因子基因家族在根部的表達差異最為顯著,其次是葉、芽、穗,在纓中的表達差異最小。干旱脅迫下,玉米HD-Zip轉錄因子基因ATHB7的下調過程受ABA信號轉導過程影響[16];SNAC1基因對下游基因OsPP18的調節基于ABA依賴途徑完成[17];AtMYB60和AtMYB96基因通過ABA依賴途徑進行氣孔的調節過程[18]。干旱脅迫下,玉米植株通過調節WRKY 46轉錄因子編碼基因的表達,促進ABA合成及側根生長,進而提高植株耐旱性[44]。
3.3 玉米干旱響應轉錄因子蛋白間的相互作用
在基因進行轉錄調控的過程中,轉錄因子蛋白間的互作起著重要作用,這是因為基因表達模式的多樣性大多來源于不同轉錄因子蛋白間的相互作用[19]。因此為達到了解轉錄因子具體作用機制和調控途徑的目的,首先需要了解干旱響應轉錄因子蛋白間的相互作用,從而進行更深層次的研究。ABA非依賴途徑和ABA依賴途徑間蛋白的互作,可以參與植物在干旱脅迫下的反應,如調控氣孔關閉、防止植物因蒸騰過度而引發的失水過多等[45]。干旱脅迫下,玉米中的ARF5蛋白和ARF9蛋白通過葉片氣孔調節作用提高了植株的抗旱能力[29];水稻中的SNAC1蛋白和DST蛋白通過參與氣孔調節過程提高了植株抗旱能力[46]。ATAF1通過與NCED3基因的啟動子區進行結合,使得NCED3基因直接控制ABA途徑的激素水平,表明了ATAF1轉錄因子在調節ABA依賴途徑中的作用[47]。干旱脅迫下,一些NAC類轉錄因子蛋白通過相互作用調控根系的伸長,進而提高植物對水分的利用率[48]。活性氧(ROS)的轉化過程對玉米抵御干旱脅迫有著重要意義,為了使活性氧的生成與轉化維持動態平衡,需要通過抗氧化劑(如抗壞血酸、谷胱甘肽)、活性氧清除酶(如抗壞血酸過氧化物酶、谷胱甘肽過氧化物酶等)進行調節[49]。當玉米植株受到干旱脅迫時,組織中與NAC、MYB和b-ZIP等轉錄因子相關的蛋白在活性氧(ROS)的轉化過程中發揮著重要作用;C2H2在滲透保護劑的合成過程以及活性氧(ROS)轉化過程中起著重要作用;WRKY轉錄因子蛋白參與了活性氧(ROS)的轉化與清除過程[20,21,50]。在分子水平上,反應機制的復雜性取決于AREB蛋白與DREB蛋白間的相互作用[51]。在玉米中,ZmWRKY17蛋白與其他蛋白相互作用,對氣孔進行調節,進而提高植株的抗旱能力[52]。據報道,玉米中的NF-YB轉錄因子和NF-YC轉錄因子會形成二聚體結構,然后與核內的NF-YA轉錄因子作用,從而對根系的發育過程進行調控[30]。
4 展望
轉錄因子在植物對干旱脅迫的響應過程中起著重要作用。本文對玉米干旱響應轉錄因子家族的種類、結構、生物學功能及作用機制進行了重點闡述,可為篩選干旱響應轉錄因子及其候選基因提供理論基礎,并進一步明確干旱響應轉錄因子家族在玉米育種過程中的應用前景。Foolad等[53]通過對轉錄因子進行篩選,發現MYB和bZIP類轉錄因子家族在植物干旱響應過程中發揮的作用較為顯著,并且目前已應用于作物分子育種。植物的抗旱調節過程由許多基因控制,因此通過轉移一個主基因只能實現較低水平的表型變異,伴隨著分子育種技術的不斷發展,干旱響應的數量性狀育種策略已從少數基因分析(通過標記選擇)轉向全基因組分析(通過基因組選擇),進而獲得更為準確的表型變異數據。植物轉錄因子家族數量較為龐大、結構較為復雜,且隨著研究的深入其數目也在增多,對于新轉錄因子的研究仍需要進行更深層次的挖掘。而隨著分子生物學技術水平的不斷提升,通過先進的技術手段對干旱響應轉錄因子進行更深入的分析研究,不僅有利于高效準確地提高玉米的品質與產量,而且可為玉米種質資源創新提供良好的理論依據。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
