不同生長年限黃芪田間雜草群落結構特征
2021-05-21 05:22:00任艷劉莎劉付松王道清吳發明
草業學報 2021年5期
任艷 ,劉莎 ,劉付松 ,王道清 ,吳發明 *
(1. 西南民族大學藥學院,四川 成都 611137;2. 遵義醫科大學,貴州 遵義 563000;3. 勁牌持正堂藥業有限公司,湖北黃石435000)
黃芪為豆科植物蒙古黃芪(Astragalus membranaceus)和膜莢黃芪(Astragalus mongholicus)的干燥根,是我國傳統常用中藥之一,具有補氣升陽,固表止汗,利水消腫,生精養血等功效,常用于各種虛證[1]。現代藥理研究表明黃芪具有免疫調節,耐缺氧、抗疲勞等藥理作用[2]。黃芪除藥用外,常作保健食品、飲料、化妝品等[3]。蒙古黃芪和膜莢黃芪對干旱環境具有極強的適應能力,是我國西北干旱半干旱地區無灌溉山區重要的經濟作物[4]。除草是中藥農業生產過程中田間管理的最主要環節[5],雜草不僅和藥用植物競爭水分和養料[6],一些優勢雜草還會擠占空間搶占光照資源,影響藥用植物光合作用[7]。由于中藥材特殊的商品屬性,在除草過程中對農藥使用的管控極為嚴格,傳統人工除草則需要投入大量人力、物力等生產成本。黃芪生產周期長,三年生以上黃芪才具有藥用價值,在生長過程中若不做好雜草的防控則會嚴重影響黃芪藥材的產量和品質。對于多年生的中藥材而言,雜草群落結構會隨著生長年限的推移發生變化,在優勢雜草發生變化時,提前選擇合適的時間和防除策略是提高田間雜草的防除效率和降低防除成本的重要舉措[8]。因此,掌握黃芪田間雜草群落的結構和演替趨勢是雜草防除決策前的基礎工作。
在國內,對大作物大豆(Glycine max)[9]、棉花(Gossypium arboreum)[10]、小麥(Triticum aestivum)[11]、玉米(Zea mays)[12]等田間雜草群落結構及其演變規律進行了較為系統的調查研究,但針對中草藥田間雜草群落結構及其演替過程的研究尚在起步階段。本研究對黃芪生長過程中田間雜草群落及其演替過程進行了調查和分析,旨在揭示黃芪田間雜草群落特征和變化規律,為其田間雜草防控時間和防控措施的抉擇提供參考依據。
1 材料與方法
1.1 研究區概況
黃芪種植區地處靖遠縣北半部哈思山自然保護區輻射農業帶的永新鄉,屬溫帶干旱半干旱氣候,年均氣溫8.9 ℃左右,極端最高氣溫35.1 ℃,年均降水量240 mm,蒸發量1634 mm,年平均日照時數2696 h,無霜期165 d[13]。該地區主要栽培藥用植物品種有枸杞(Lycium chinense)、黃芪、甘草(Glycyrrhiza uralensis)、黨參(Co?donopsis pilosula)等。黃芪的種植歷史有十余年,種植品種主要為蒙古黃芪,夾雜少量膜莢黃芪和其他品種。該地區種植黃芪方式為種子直播和育苗移栽,不同生長年限黃芪基本處于半野生狀態,為黃芪田間雜草群落結構及演替規律的調查研究提供了便利。
1.2 調查方法和樣點分布
于2019 年7?8 月對黃芪種植區不同生長年限的黃芪田中雜草種類進行田間調查,不同生長年限黃芪選擇3塊農田,陰坡、陽坡和山梁各選擇一塊作為樣地,部分年限種植田塊較少但同一塊田中包含了陰坡、陽坡和山梁,則按地形(即陰坡、陽坡和山梁)將同一塊黃芪田分為3 個樣地,每塊樣地采用倒置“W”9 點取樣法選取樣點,樣方面積為1 m×1 m=1 m2,調查時記錄不同田塊雜草種類和數量(為了便于統計雜草數以雜草主莖、稈數記)[14]。植物鑒定參考《中國植物志》全文電子版[15]、《中國常見植物野外識別手冊·荒漠冊》[16]、《中國常見植物野外識別手冊·祁連山冊》[17]。
1.3 數據統計與分析
根據調查數據計算不同生長年限黃芪田中雜草的物種多樣性、相對優勢度及群落的相似性,采用Excel 2016進行數據處理和統計。物種豐富度(Species)即樣方中包含的所有雜草種類數。Shannon?Wiener 多樣性指數(H′):H′= ?∑Pi·lnPi,式中Pi為樣方中第i物種第 1 次被抽到的概率,Pi=Ni/N,Ni為樣方中第i種雜草的密度,N為該樣方中所有雜草密度之和。Pielou 均勻度指數 (J):J=H′/lnS,S是調查區域內物種數[18?19]。Simpson 優勢度指數相對優勢度 (relative advantage,RA:RA) =Di/∑Di× 100%[20]。Sorenson 指數 (Cs):Cs=2j/(a+b),測定群落相似度,式中j為群落A 與B 所共有的物種數,a為群落A 含有的全部物種數,b為群落B 含有的全部物種數[21]。
2 結果與分析
2.1 黃芪田雜草區系
黃芪田中共調查到的雜草有66 種,分屬24 科59 屬。雜草所屬科屬中單種科有10 個科,占總科數的41.67%,單種屬53 個,占總屬數的89.83%;雙種科有7 個,占總科數的29.17%,雙種屬有5 個,占總屬數的8.47%;僅蒿屬雜草有3 個種。以菊科和豆科雜草最多,分別有10 種,兩個科雜草種類占總數的30.30%;禾本科次之,雜草種類有6 個;其次為唇形科和莧科,分別有5 種。菊科、豆科、禾本科、唇形科和莧科5 個科共有雜草種數36 個,占總種數的54.55%。雜草種類較多的在種間分布為聚集型,在屬間分布為分散型。
2.2 黃芪田中雜草種類
根據雜草的綜合優勢度、田間出現頻度、在不同年限黃芪田的具體發生情況和實際危害,可以將黃芪田中雜草劃分為4 種類型,即優勢雜草、區域性優勢雜草、常見雜草和一般雜草[22]。
優勢雜草在不同生長年限黃芪田全部樣方地中均有分布,發生優勢度高,對黃芪生產危害大,防除困難,主要有:冰草(Agropyron cristatum)、蟲實(Corispermum mongolicum)和狗尾草(Setaria viridis)。
區域性優勢雜草為發生優勢度高,在不同生長年限均有分布,對黃芪生長影響大,防除困難,但在部分樣方地中缺失的雜草品種,主要有:田旋花(Convolvulus arvensis)、苦荬菜(Ixeris polycephala)、小薊(Cirsium setosum)、乳苣(Lactuca tatarica)。
常見雜草為優勢度較高,防除難度一般,僅在部分年限黃芪田中出現的雜草,主要有:二裂委陵菜(Potentilla bifurca)、披針葉決明(Thermopsis fabacea)、獨行菜(Lepidium apetalum)、紫花苜蓿(Medicago sativa)、甘草(Glyc?yrrhiza uralensis)、密花香薷(Elsholtzia densa)、糙葉黃耆(Astragalus scaberrimus)、刺葉柄棘豆(Oxytropis aci?phylla)、醉馬草(Achnatherum inebrians)、擬糙葉黃耆(Astragalus pseudoscaberrimus)、菘藍(Isatis tinctoria)、冷蒿(Artemisia frigida)、藜(Chenopodium album)、車前(Plantago asiatica)、蒲公英(Taraxacum mongolicum)、豬毛菜(Salsola collina)、草木樨(Melilotus officinalis)、微孔草(Microula sikkimensis)、雜配藜(Chenopodium hybridum)。
一般雜草為優勢度低、發生頻率不高的雜草品種,主要有:野胡蘿卜(Daucus carota)、豬殃殃(Galium spuri?um)、鶴虱(Lappula myosotis)、駱駝蓬(Peganum harmala)、白花枝子花(Dracocephalum heterop)、狗娃花(Aster hispidus)、柴胡(Bupleurum chinense)、黃花蒿(Artemisia annua)、地梢瓜(Cynanchum thesioides)、苦苣菜(Sonchus oleraceus)、澀薺(Malcolmia africana)、鹽生草(Halogeton glomeratus)、細柄野蕎麥(Fagopyrum gracilipes)、胡枝子(Lespedeza bicolor)、蒙古芯芭(Cymbaria mongolica)、芨芨草(Achnatherum splendens)、夏至草(Lagopsis supi?na)、野燕麥(Avena fatua)、黃花角蒿(Incarvillea sinensis var.przewalskii)、尖齒糙蘇(Phlomis dentosa)、阿拉善馬莧蒿(Pedicularis alaschanica)、蓼子樸(Inula salsoloides)、銀灰旋花(Convolvulus ammannii)、針茅(Stipa capilla?ta)、大籽蒿(Artemisia sieversiana)、宿根亞麻(Linum perenne)、乳漿大戟(Euphorbia esula)、牻牛兒苗(Erodium stephanianum)、小車前(Plantago minuta)、沙棘(Hippophae rhamnoides)、檸條(Caragana korshinskii)、紅花巖黃耆(Corethrodendron multijugum)、榆樹(Ulmus pumila)、萹蓄(Polygonum aviculare)、黃芩(Scutellaria baicalen?sis)、小葉鐵線蓮(Clematis nannophylla)、黃花鐵線蓮(Clematis intricata)、野西瓜苗(Hibiscus trionum)、甘青鐵線蓮(Clematis tangutica)、杏樹(Armeniaca vulgaris)。
此外,黃芪田中部分雜草為外來種,主要為近年來引種栽培的牧草、中藥材種子逃逸或根莖走竄入侵農田形成的雜草,有紫花苜蓿、沙棘、黃芩、檸條、菘藍。
2.3 不同生長年限黃芪田雜草群落的物種多樣性
黃芪田中雜草的多樣性整體表現為栽種年限越長雜草種類越多,以六年生黃芪田中雜草的物種豐富度最高,達到48 種,占統計總數的72.73%,其次為八年生和十年生,各有47 種。一年生黃芪田間雜草的Shannon?Wiener 指數和 Pielou 指數均最低,而 Simp?son 指數最高,說明一年生黃芪田間雜草的離散度最小,集中性高,其群落結構中優勢種相對其他生長年限更為明顯,群落結構的穩定性相對較差。十年生黃芪田間雜草的Shannon?Wiener 指數和Pielou 指數均最高,分別為一年生黃芪田1.84 和1.67 倍,而Simp?son 指數最低,僅是一年生黃芪田的18.28%。而九年生黃芪田間雜草群落的Shannon?Wiener 指數和Simpson 指數和十年生黃芪田趨于接近,Pielou 指數基本相同。這說明九至十年生黃芪田中雜草的分散度較高,雜草的種類和數量趨于平衡,群落結構相對穩定(表1)。
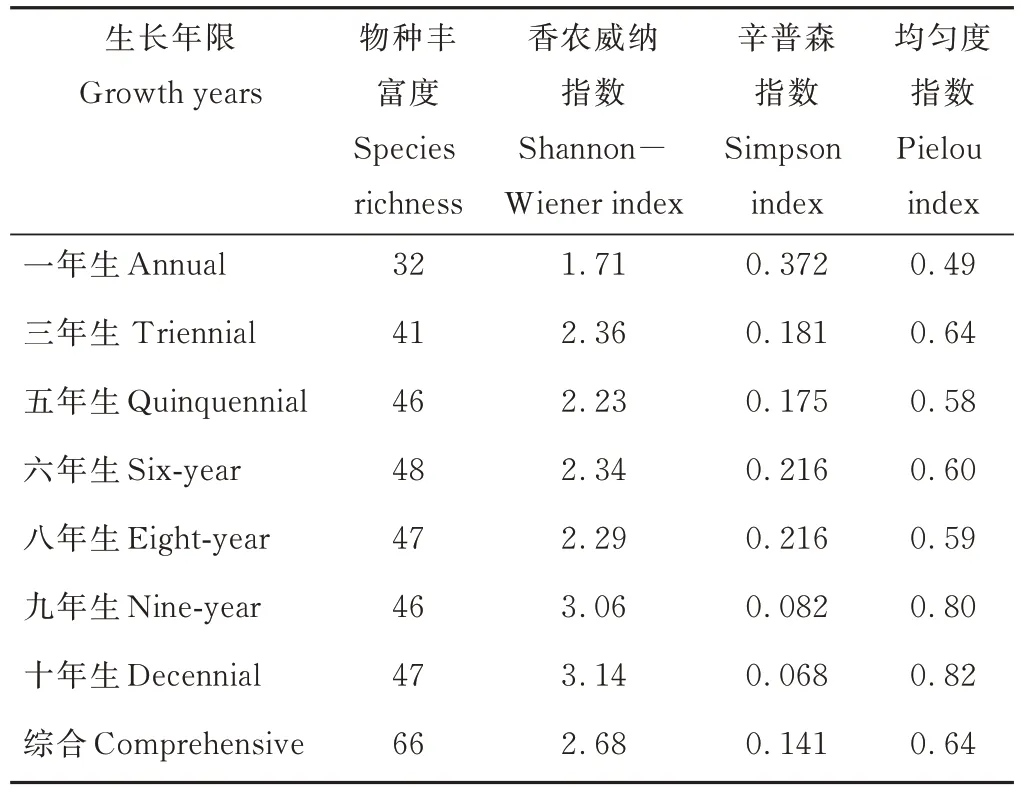
表1 黃芪田雜草群落的物種多樣性Table 1 Species diversity of weed community in field of A.membranaceus
2.4 不同生長年限黃芪田雜草發生的特點及群落結構
黃芪田中雜草群落結構伴隨黃芪生長年限的增加而變化。從雜草發生的優勢度來看(表2),冰草為黃芪田中主要優勢雜草,綜合優勢度達到72.29,其次為苦荬菜,綜合優勢度達到10.71,綜合優勢度大于1 的雜草還有狗尾草、蟲實、田旋花和乳苣。一年生黃芪田距離耕作時間短,在耕作過程中對田間宿根性雜草(多年生,主要以根莖走竄傳播)進行有效防除,以根莖走竄傳播為主的雜草傳播速度相對較慢,而疏松的耕作層為一年生種子傳播雜草的萌發和生長提供了良好的條件,因此一年生黃芪田間雜草以一年生種子雜草優勢度高,進而構成了“蟲實+狗尾草”為優勢種的群落結構。三年、五年、六年和八年生黃芪田間一年生種子雜草的數量逐漸減少,在雜草種間的生存空間競爭優勢逐漸喪失,而主要以走竄傳播的多年生雜草逐漸占據優勢地位,形成了以“冰草+苦荬菜+乳苣+田旋花”為優勢種的群落結構。九年生和十年生黃芪田間雜草逐漸演變成為“冰草+n”多優勢種的結構特征,苦荬菜、乳苣、田旋花依舊為優勢種,但已經喪失了絕對主導地位。大量多年生雜草逐漸形成較強的競爭力,野決明(Thermopsis lupinoides)、小薊、糙葉黃耆等多年生植物在部分樣方開始占據主導地位,尤其以醉馬草、冷蒿為代表的大草本和類灌木植物在黃芪田間的生存競爭力凸顯出來,逐漸占據主導地位(從附近退耕還林地帶可以觀察到超過十五年以上的荒地冷蒿和醉馬草為絕對優勢種)。
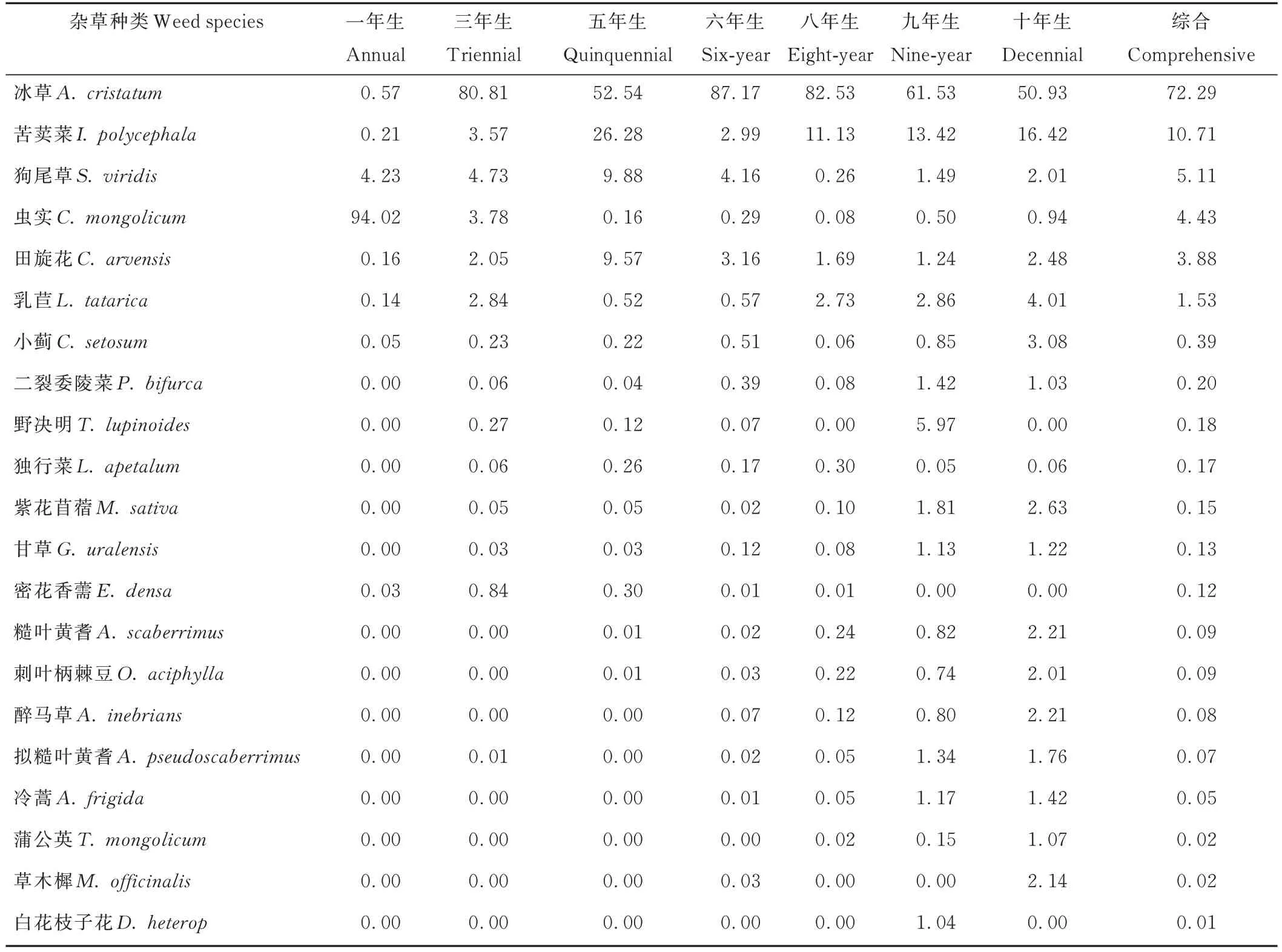
表2 黃芪田主要雜草的相對優勢度Table 2 Relative dominance of main weeds in field of A.membranaceus
2.5 不同生長年限黃芪田雜草群落的相似性
從雜草群落物種組成的相似性來看(表3),生長年限相近的黃芪田間雜草群落結構相似,以三年生和五年生黃芪田最為相似,Sorenson 指數達到0.828,這說明三至五年生黃芪田間雜草群落結構最為相近。生長年限相差越大則田間雜草群落結構的相似度越低,一年生和九年生黃芪田間雜草群落組成差異最大,Sorenson 指數僅為0.513。十年生黃芪田間雜草群落結構除和一年生田間雜草群落結構的相似度最低之外,和其余生長年限黃芪田間雜草群落結構之間的相似度接近,Sorenson 指數均在0.75 左右。分析結果表明:在黃芪田間雜草群落演替的時間軸上,排除人為干擾,距離相近的兩個時間點雜草群落結構趨同,時間點越遠雜草群落結構之間相似性降低。
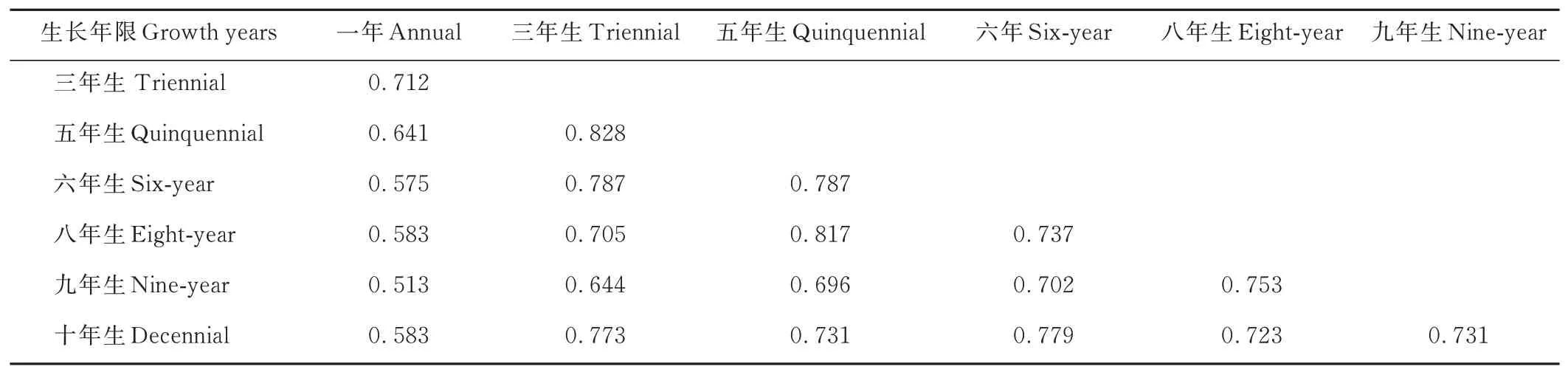
表3 不同生長年限黃芪田雜草群落相似性Table 3 Similarity of weed community in field of A.membranaceus with different growth years(Sorenson index)
將各生長年限黃芪田間全部雜草(共66 種雜草,樣方中沒有某種雜草記為0)構成矩陣進行系統聚類分析,不同生長年限黃芪田間雜草群落物種和數量構成的相似性結果表現和Sorenson 指數基本一致(圖1),7 個不同生長年限黃芪田間雜草群落結構可分為2 個大組別,4 個小組別。一年生單獨為1 個小組,三年生和八年生分為1 個小組,五年生和六年生分為1 個小組,九年生和十年生分為1 個小組。4 個小組別特征表明生長年限相近的黃芪田間雜草群落結構趨于一致,三年生和八年生黃芪田間雜草群落特征被聚為一個小類,一年生和九年生、十年生黃芪田間雜草群落特征被聚為一大類,這兩個聚類特征表明不同年限黃芪田間雜草的特征表現為兩頭小(種類少、數量少)、中間大(種類多、數量多)的特征。
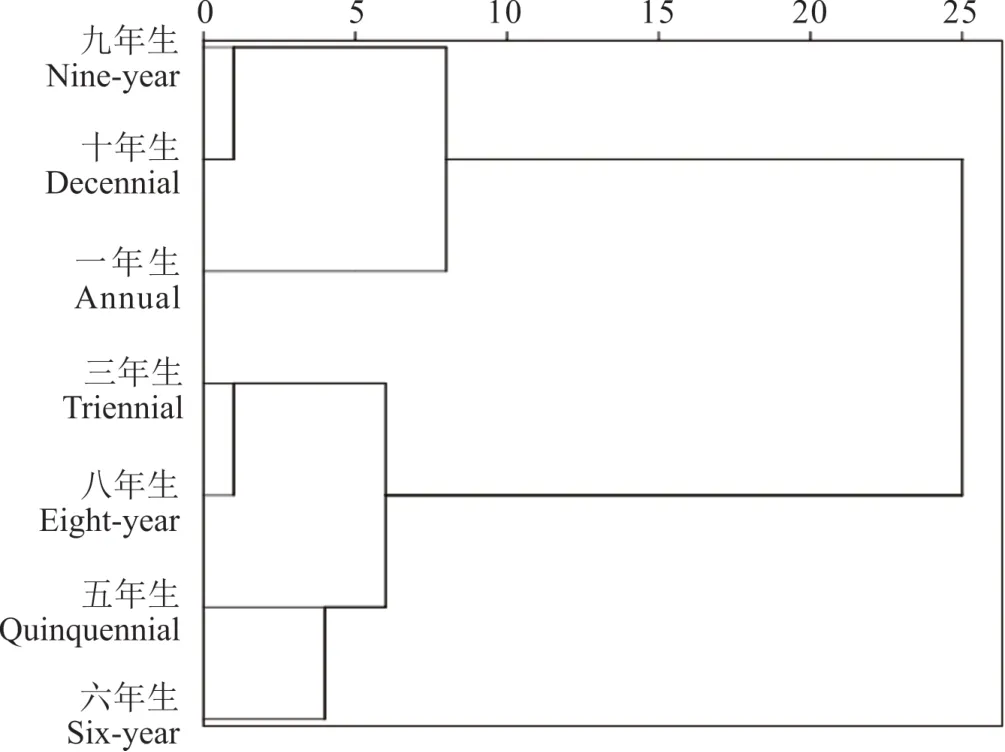
圖1 不同生長年限黃芪田雜草群落的聚類分析Fig. 1 Cluster analysis of weed communities in A. membra?naceus with different growth years
3 討論與結論
雜草既是農田生態系統的重要構建者,也是限制農業生產的主要生物因子[23]。調查結果表明黃芪田間惡性雜草主要有禾本科的冰草,莧科的蟲實,菊科的苦荬菜、乳苣、小薊,旋花科的田旋花等。調查發現黃芪田間雜草的傳播主要有兩種模式,一種以種子傳播的游擊戰模式,另一種以根莖走竄傳播的根據地模式。種子隨風傳播的代表為蟲實、豬毛菜、澀薺、小薊、獨行菜、苜蓿等;蟲實和豬毛菜為干枯植株從地面脫落卷曲成輪狀,整個植株隨風游走將種子傳播開來;小薊、苦荬菜、乳苣、蒲公英等菊科植物則借助果實上的冠毛隨風傳播[24]。冰草根莖末端尖銳鋒利,在疏松的農田中一年內可延伸數米遠,縱向傳播后又會橫向分散傳播,最終占領全部土地空間,其根莖再生能力極強,是以根莖走竄傳播的惡性雜草的典型代表。苦荬菜、乳苣、田旋花等不僅能夠通過種子傳播,其根莖也極具傳播能力。因此針對不同雜草的傳播特征在黃芪生產過程要制定針對性的雜草管控方法才能有效地提高防治效果,以種子傳播的雜草多為一年生植物,這類雜草的防除盡量選擇在其開花結果之前進行,避免其大量結種,同時除掉的雜草如果要回田則需要完全腐熟破壞種子的生活才行,特別是動物肥需要完全腐熟;而以根莖走竄傳播的雜草需要斷絕其傳播途徑,如深除其根、倒茬、干旱季節深耕暴曬等。另外,田埂部位雜草也應及時防除避免其田間傳播。
兩種傳播方式的雜草在不同生長年限黃芪田中表現不同,究其原因是黃芪生長年限較短的農田,地表疏松,有利于種子生根發芽,依靠根莖走竄擴大種群的植物相對擴展速度較慢,因此生長年限較短的黃芪田中一年生種子雜草是構建群落的優勢種,以根莖繁殖的雜草則多分布在田埂邊遠地帶。伴隨著年限增長,黃芪田疏松的土壤結構逐漸變得瓷實,地表逐漸變光滑,導致種子植物繁殖難度逐漸增加,而依靠根莖走竄繁殖的植物則逐漸占領整個農田成為構建雜草群落的優勢種。同時,黃芪植株逐漸長大,有效地擠占了雜草生存空間,這導致小草本在黃芪田間生存環境逐漸變得艱難。因此,一年生種子傳播的小草本伴隨黃芪生長年限的延長數量逐漸減少甚至消亡,黃芪田間雜草群落結構發生改變,能夠和黃芪競爭并能夠獲得相應生存空間的大草本、類灌木植物醉馬草、冷蒿等植物逐漸出現。當灌木、類灌木出現在黃芪田中時,黃芪田中整個生態結構已經發生很大的變化,土壤結構和黃芪本身也已經開始嚴重退化,黃芪根開始腐化、空心,不適合作為藥材使用。
使用除草劑是現代農業最為常用的田間雜草防除方式[25],但針對小眾農作物的中藥材而言專屬除草劑的開發研究與實際應用和傳統大農業品種相比相差甚遠,而且除草劑的濫用是中藥材農藥殘留超標的重要原因[26]。人工除草是除草劑防控雜草之外最常用的田間雜草防除方法,其優點在于綠色環保,但高昂的人工成本往往迫使藥農廣泛選擇農藥防除雜草。因此如何有效降低人工除草成本,提高雜草防控效果就成為中藥農業產業發展過程中的關注點。綜上,掌握田間雜草發生規律,依據中藥材田間雜草群落的結構和發生特點,選擇適當的防控措施是達到更為高效、經濟、綠色的雜草治理目的的重要途徑。
猜你喜歡
科教新報(2022年22期)2022-07-02 12:34:28
小讀者(2021年2期)2021-03-29 05:03:48
趣味(語文)(2020年3期)2020-07-27 01:42:46
華人時刊(2019年13期)2019-11-17 14:59:54
文苑(2018年22期)2018-11-19 02:54:14
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
現代農業(2015年5期)2015-02-28 18:40:49
中國火炬(2014年11期)2014-07-25 10:31:58
雜草學報(2012年1期)2012-11-06 07:08:33
