葡萄果實酒石酸生物合成研究進展*
2021-05-23 09:01:48曹慧玲舒河霖邵建輝張海明馬春花
中國果樹 2021年4期
曹慧玲,舒河霖,邵建輝,張海明,馬春花
(1 云南農業大學園林園藝學院,昆明 650201)(2 云南農業大學云南省高校葡萄與葡萄酒工程技術研究中心)
葡萄是世界上栽植最廣泛的果樹之一,有機酸是葡萄果實內在品質的重要組分之一,直接影響葡萄果實口感,并賦予葡萄酒理想的感官特性和抗氧化能力[1]。葡萄果實中有機酸主要包括酒石酸、蘋果酸,以及少量的乳酸、乙酸、檸檬酸和琥珀酸等,它們的含量與組分是決定葡萄果實風味和品質的重要因素。其中酒石酸占有機酸含量的42.8%~77.0%,是葡萄漿果中的主要有機酸,決定了葡萄、葡萄酒的pH值和酸性[2]。此外,酒石酸在葡萄果實轉熟過程中具有相對較好的代謝穩定性,不像蘋果酸含量極易受環境高溫、呼吸和糖異生作用影響而迅速下降[3]。由于酒石酸在葡萄果實和葡萄酒中具有重要意義,國內外學者對其進行了大量研究,并發現了一些與酒石酸生物合成相關的酶及基因,但葡萄果實酒石酸完整的生物合成途徑尚不明確。因此,筆者綜述了目前國內外葡萄果實酒石酸的相關研究,為今后進一步解析葡萄果實酒石酸的完整生物合成途徑和調控機制提供參考。
1 酒石酸
1.1 酒石酸的來源及結構
酒石酸(Tartaric acid,TA),分子式C4H6O6,學名2,3-二羥基丁二酸,白色單體,有強酸味,易溶于水和乙醇,是一種二元羧酸[4],最早在1769年由瑞典化學家卡爾·威廉·舍勒發現。起初因在葡萄酒釀造過程中的發酵罐壁上發現一種晶體,其形狀像石頭而被命名為酒石,后經研究發現其主要成分是酒石酸氫鉀,在葡萄酒釀造過程中易沉淀,可以從酒石酸氫鉀分離出酒石酸,因此,傳統上的酒石酸是葡萄酒發酵過程中產生的固體副產品[5-6]。
酒石酸化學手性最先由Jean-Baptiste Biot[7]于1832年發現,根據旋光面性質的不同,分為2種對映異構體——L(+)-TA和D(?)-TA(圖1)。自然界存在的酒石酸為L(+)-TA[8-9],是葡萄屬植物和天竺屬植物中特有的酸,而在模式植物如擬南芥、番茄以及其他水果和經濟作物中未見積累。D(-)-TA在天然來源中很少存在,通常被認為只由某些微生物種形成。

圖1 L(+)-TA和D(?)-TA的費歇爾投影式
1.2 酒石酸在葡萄中的作用
酒石酸是葡萄果實和葡萄酒所特有的有機酸,因此又名葡萄酒酸。葡萄漿果中的酒石酸酸性較強,是葡萄、葡萄酒酸度的主要貢獻者,在一定程度上決定了葡萄果實的釀酒品質;有機酸有助于抑制微生物繁殖,葡萄汁的pH值主要取決于漿果中的酒石酸含量,漿果中的酒石酸可以將葡萄酒的pH值維持在3.0~3.5,使葡萄酒顏色及其微生物保持穩定。此外,酒石酸含量影響葡萄果實風味的平衡,過高會影響果實的適口性,過低則果實平淡無味。釀酒師可以在葡萄酒釀造過程中,通過添加酒石酸,最大限度地減少氧化和微生物腐敗,從而提高葡萄酒的感官特性和老化潛力;若酒石酸被某些細菌代謝,會使葡萄酒風味缺乏圓潤感,導致泛渾病。
2 葡萄果實酒石酸的生物合成
早期的研究表明,酒石酸存在于葡萄葉片中,而其在葡萄漿果中的積累是由葉片合成運轉而來,但Hale和Hardy在20世紀60年代初證明葡萄漿果本身就是合成酒石酸的場所[10]。Debolt等[11]使用抗壞血酸進行放射標記,也證明了酒石酸的合成主要發生在葡萄漿果中。Debolt等[11]發現酒石酸在葡萄花后1個月內大量積累,而Deluc等[12]研究表明酒石酸生物合成積累通常發生在漿果轉色期前,之后幾乎沒有新的酒石酸合成。通常酒石酸在果實生長發育過程中逐漸積累,在轉色之后濃度開始下降,成熟期酒石酸濃度下降主要是由于漿果體積增大導致酒石酸被稀釋的結果[12]。在葡萄漿果成熟和隨后的釀酒過程中,酒石酸不會通過呼吸作用被葡萄漿果細胞代謝,保持代謝穩定[13]。放射性同位素標記和示蹤技術,揭示了葡萄果實中酒石酸的合成前體是抗壞血酸,在C4/C5位點裂解后逐步催化生成酒石酸[11,14-15]。目前,人們對這一途徑知之甚少,也沒有研究發現酒石酸參與各種初級代謝途徑[16],因此要探明酒石酸生物合成途徑還需要進一步研究。
2.1 葡萄果實酒石酸生物合成途徑
研究表明,葡萄酒石酸生物合成的主要前體物質是抗壞血酸(AsA)[14,17],通常認為酒石酸合成途徑中存在2個不同的階段,即上游階段和下游階段(圖2)。
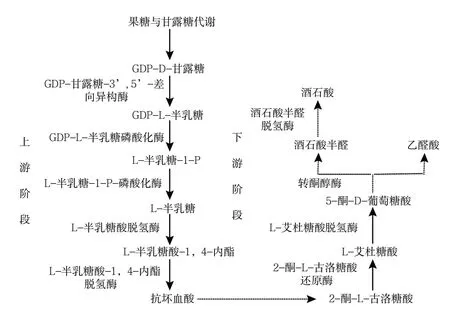
圖2 葡萄果實可能的酒石酸合成途徑[18-19]
(1)上游階段:合成酒石酸前體物質抗壞血酸。在植物中至少已經報道了4種抗壞血酸生物合成途徑。現已知抗壞血酸生物合成的主要途徑是基于光合作用的碳通量通過一系列酶催化反應的結果,由Wheeler等[20]在無碳鏈倒位和L-半乳糖酸-1,4-內酯是抗壞血酸合成的直接前體下提出,稱為Smirnoff-Wheeler途徑,即L-半乳糖途徑。這條途徑在葡萄漿果的生長階段占主導地位[21],其關鍵中間體是L-半乳糖(L-Gal),此途徑涉及D-葡萄糖轉化為GDP-D-甘露糖(GDP-D-Man)、GDP-L-半乳糖(GDP-L-Gal)、L-半乳糖、L-半乳糖酸-1,4-內酯(L-GalL)和抗壞血酸[22]。在此途徑中,GDP-D-甘露糖通過GDP-甘露糖-3′,5′-差向異構酶(GME)催化生成GDP-L-半乳糖,GDP-L-半乳糖經GDP-L-半乳糖磷酸化酶(VTC2)催化生成L-半乳糖-1-P,再經L-半乳糖-1-P-磷酸化酶(VTC4)催化生成L-半乳糖[23],之后L-半乳糖經過L-半乳糖酸脫氫酶(L-GalDH)脫氫生成抗壞血酸的直接前體L-半乳糖酸-1,4-內酯,接著被L-半乳糖酸-1,4-內酯脫氫酶(L-GLDH)催化生成抗壞血酸。該途徑涉及的酶多數已被檢測、純化或克隆。
(2)下游階段:合成酒石酸。結合研究者的發現和各產物的結構式,推斷酒石酸生物合成是抗壞血酸經過水解和氧化轉化為2-酮-L-古洛糖酸(2-KGA),接著可能被2-酮-L-古洛糖酸還原酶(2-KGR)還原為L-艾杜糖酸,然后L-艾杜糖酸被L-艾杜糖酸脫氫酶(L-IdnDH)氧化成5-酮-D-葡萄糖酸(5-KGA)。5-酮-D-葡萄糖酸在C4和C5之間裂解產生4碳的L-threo-tetruronate,接著被氧化形成酒石酸(圖2、圖3)。但植物體內酒石酸生物合成的途徑及生物合成過程中的相關酶還需要進一步驗證和證實。
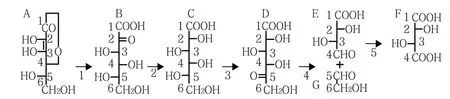
圖3 葡萄酒石酸生物合成途徑中間產物結構式[24]
2.2 葡萄果實酒石酸生物合成途徑相關酶及基因
目前,國內外對植物中抗壞血酸生物合成和代謝已有較多的研究[13,25-26],關于L-半乳糖途徑中涉及的大多數酶已被檢測、純化或克隆(表1)。其中GME、VTC2、L-GalDH等被認為是抗壞血酸生物合成階段的關鍵酶,其酶活性和酶基因表達量對抗壞血酸含量的調控極其重要[15,27-29]。Tabata等[30]和安華明等[31]分別在煙草和刺梨中驗證了L-GLDH的功能,最近,劉慶帥[32]的研究也證實了L-GLDH基因與黑穗醋栗果實抗壞血酸的積累顯著相關,且品種間抗壞血酸含量的差異可能主要受L-GLDH調控。但目前葡萄果實從抗壞血酸到酒石酸生物合成途徑中相關酶及基因的研究還較少[15],在下游階段,抗壞血酸經過氧化、還原反應最終合成酒石酸,可能涉及到一系列的水解酶、氧化酶、還原酶和裂解酶[33],其相關研究取得了一定的進展。
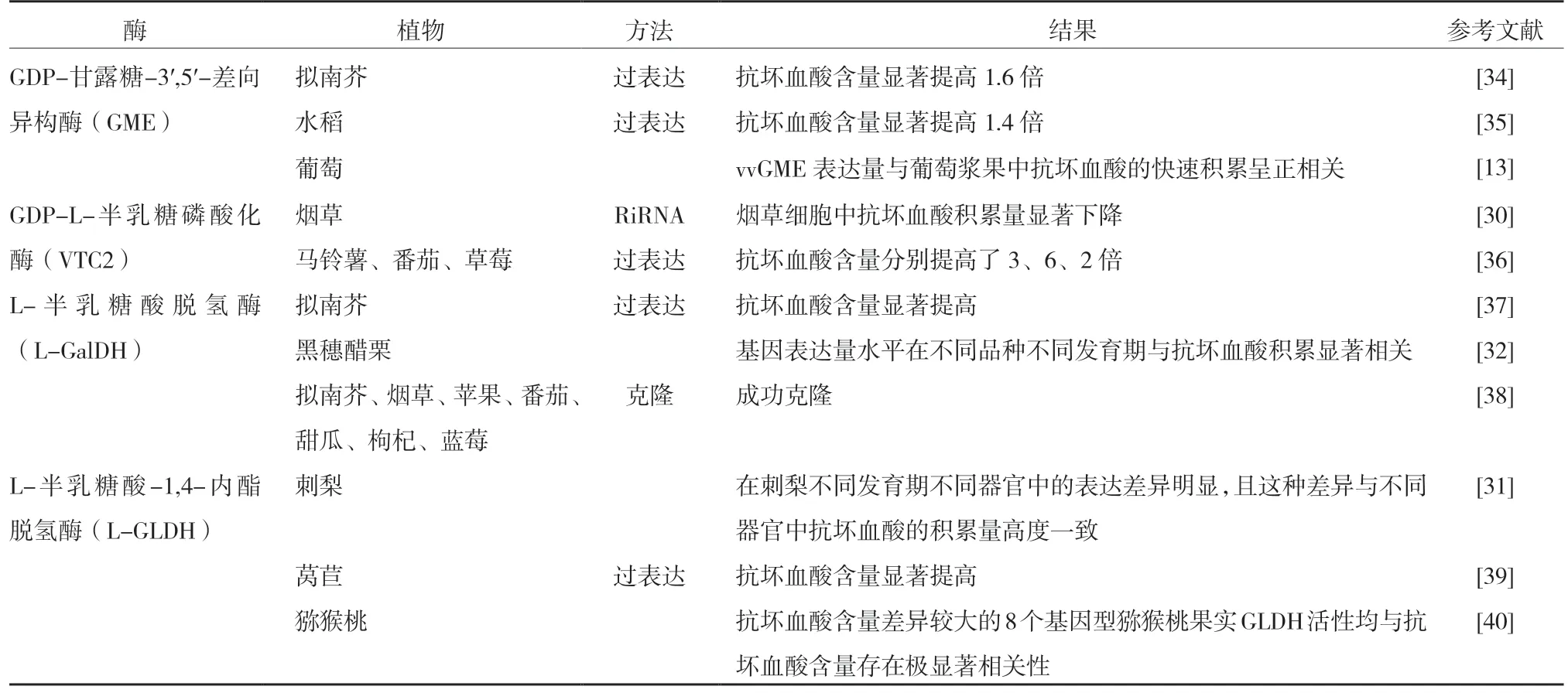
表1 L-半乳糖途徑關鍵酶基因的相關研究
2.2.1 L-艾杜糖酸脫氫酶(L-IdnDH)
L-艾杜糖酸脫氫酶催化L-艾杜糖酸氧化成5-酮-D-葡萄糖酸,迄今為止,它是在從抗壞血酸到酒石酸合成過程中被確定的酶之一,是在酒石酸合成途徑中鑒定出的唯一基因,同時也是限速酶[14]。Debolt的酶學實驗以及早期的14C同位素飼喂實驗證實了艾杜糖酸脫氫酶所催化的反應是葡萄屬植物中酒石酸生物合成的限速步驟[24];Debolt等[14]通過對葡萄屬25個品種的果實有機酸譜庫分析,發現烏頭葉蛇葡萄(Ampelopsis aconitifolia)不含酒石酸,且沒有L-艾杜糖酸脫氫酶基因轉錄物;而且Wen等[41]也發現葡萄果實中VvIdnDH的轉錄水平和翻譯水平與酒石酸含量的變化趨勢相同,并且在成熟漿果中幾乎沒有檢測到VvIdnDH轉錄,進一步推斷葡萄成熟期可能不合成酒石酸;Ren等[42]通過CRISPR/Cas9系統,靶向編輯了L-艾杜糖酸脫氫酶基因,對‘霞多麗’葡萄懸浮細胞進行了遺傳轉化,轉基因細胞團中酒石酸含量表現為降低的趨勢,這表明L-IdnDH調控酒石酸的生物合成,證明L-IdnDH是葡萄酒石酸生物合成的關鍵酶。但也有研究者提出了不同的看法,Cholet等[15]認為L-IdnDH可能不是酒石酸生物合成途徑中唯一的關鍵酶,該酶可能或多或少地與其他尚未鑒定的酶協同起作用,并且可能受氣候條件影響較大。另外,天竺屬植物酒石酸生物合成方式與葡萄不同,由抗壞血酸C2/C3的裂解形成L-蘇糖酸后進一步形成酒石酸,Narnoliya等[19]對玫瑰香天竺葵酒石酸的研究中卻檢測到了葡萄L-IdnDH基因的表達,說明L-IdnDH基因可能在玫瑰香天竺葵酒石酸生物合成中起作用,天竺屬植物酒石酸生物合成途徑可能存在抗壞血酸C4/C5的裂解。
2.2.2 2-酮-L-古洛糖酸還原酶(2-KGR)
2-酮-L-古洛糖酸還原酶催化2-酮-L-古洛糖酸(2-KGA)還原為L-艾杜糖酸。許多微生物中已經報告了具有催化此化學反應能力的酶,但是其在植物中少見報道。大腸桿菌中2-KGR的酶學表征驗證了其在大腸桿菌中可以將2-KGA還原為L-艾杜糖酸[33,43],但Ford[44]認為該酶可能是乙醛酸還原酶(GR),其在足夠的底物存在下可以充當羥基丙酮酸還原酶或2-KGR。Jia等[45]從葡萄中初步鑒定了一個具有2-酮-L-古洛糖酸還原酶活性的醛酮還原酶Vv2KGR,使用NADPH作為首選的輔酶,Vv2KGR可以有效地將2-酮-L-古洛糖酸(2-KGA或2KLG)還原為L-艾杜糖酸,這說明了Vv2KGR參與從抗壞血酸到酒石酸生物合成的過程。
2.2.3 Transketolase(TK)和Tartaric semialdehyde dehydrogenase(TSAD)酶
到目前為止,對推測催化5-KGA生成酒石酸半醛的TK以及催化酒石酸半醛生成酒石酸TSAD的研究甚少。Debolt[46]利用葡萄基因組數據初步篩選到TK和TSAD的候選基因。Debolt[46]從葡萄中分離出的重組TK能夠使用5-keto gluconic acid和4-磷酸赤蘚糖作為底物形成酒石酸半醛和6-磷酸果糖,但活性非常低;而且Ford[44]分離出的2個葡萄TK基因(vvTKI和vvTKII)在pET 14b表達中不具有轉酮酶活性;Debolt提供的TSAD的候選基因編碼了植物中琥珀酸半醛脫氫酶的同源基因,有可能將酒石酸半醛氧化為酒石酸,但迄今為止未有相關報道。目前TK和TSAD基因的表征和功能尚不明確,還需要進一步研究和證實。
2.3 葡萄果實酒石酸生物合成的調控
目前關于葡萄果實酒石酸生物合成積累及調控研究也取得了一些進展。葡萄不同屬間果實酒石酸含量存在差異,比如葡萄屬蛇葡萄科中烏頭葉蛇葡萄天然缺乏酒石酸和L-艾杜糖酸脫氫酶基因轉錄物,且表現出極高水平的抗壞血酸水平[46]。同時,酒石酸在葡萄果實不同部位中含量不同,并且隨著果實的發育和成熟,果皮和果肉外部中的酒石酸濃度降低,而果肉內部和種子附近區域的酒石酸濃度保持相對不變[47];此外,Lamikanra等[48]在比較葡萄不同組織中有機酸含量研究發現,成熟期葡萄果實將近70%的有機酸分布在果皮,而種子含酸量卻很少;Falchi等[49]應用高效液相色譜法對葡萄果肉、果皮和種子的成分進行分析,發現不同品種葡萄果實不同部位酒石酸的含量存在差異;趙悅等[50]研究發現,不同產地的‘赤霞珠’葡萄成熟果實果皮、果肉的酒石酸含量存在差異,果皮中酒石酸含量均高于果肉,這說明酒石酸的積累具有組織特異性。
此外,光照對葡萄酒石酸的生物合成起著重要作用。葡萄植株經高光和弱光處理后,酒石酸水平隨著光照強度的增加而增加,L-艾杜糖酸脫氫酶基因的表達量也發生變化[46]。Debolt等[51]在葡萄花期用黑色塑料盒遮光以排除光線,同時保持環境溫度在一定范圍內,結果顯示出每個漿果的酒石酸含量降低。研究表明一些植物生長調節劑對酒石酸的生物合成代謝具有重要調控作用。Ziliotto等[52]在花后65 d用NAA處理葡萄后立即觀察到酒石酸的顯著增加;葡萄花序與果穗經6-BA處理后,抑制了L-艾杜糖酸脫氫酶基因表達,對酒石酸含量產生了顯著的抑制作用[53];‘極早蜜’葡萄在花后60 d葉面噴施水楊酸可以抑制果實酒石酸的積累[54]。
這些研究結果表明,酒石酸在不同類型葡萄、葡萄果實不同部位的含量有差異,且一些外界條件(如光照、植物生長調節劑等)會影響酒石酸的生物合成,這對我們研究和驗證葡萄果實酒石酸生物合成途徑中未被證實的一些酶(如轉酮醇酶、酒石酸半醛脫氫酶等)基因的功能提供了參考依據。
3 研究展望
全球氣候變暖對葡萄果實品質有顯著影響,葡萄果實中有機酸含量主要受溫度影響。因為蘋果酸在果實成熟過程中會因較高的溫度降解,導致漿果酸度下降較快[55],但漿果中酒石酸濃度對高溫的敏感性遠低于蘋果酸濃度。漿果中酒石酸含量高的品種對氣候變化不那么敏感,在漿果成熟期間,每個漿果中的酒石酸含量是相對穩定的[51],因此酒石酸含量高的品種能更好地適應未來不斷變化的氣候條件。有些溫暖地區的葡萄在釀酒過程中因其自然酸度不滿足要求,釀酒師通常需要人工添加更多的酒石酸,導致釀酒成本過高[56]。所以研究影響葡萄果實酒石酸含量的因素,探明酒石酸在葡萄中的生物合成規律以及如何對葡萄果實的酒石酸積累進行有效調控將成為今后的研究熱點。
因此,圍繞以下2個方面的深入探討可能是未來葡萄酒石酸研究領域的主要方向:①目前葡萄果實酒石酸生物合成途徑中的酶尚未被完全證實,其生物合成主要關鍵酶基因的克隆、功能鑒定、遺傳轉化等還有待研究。尤其是TK基因的表征和功能尚不明確,還需要進一步研究和證實;②葡萄果實酒石酸積累是一個復雜的生物學過程,目前關于葡萄果實酒石酸含量的調控因素研究以光照、植物生長調節劑處理為主,其他脅迫因素如高(低)溫、干旱或水澇等生物脅迫或非生物脅迫對酒石酸合成的影響研究較少,這都需要進一步研究。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
中國衛生(2016年3期)2016-11-12 13:23:26
