長湖浮游動物群落結構特征及其季節變化
2021-05-24 01:52:34龔森森吳嘉偉羅靜波譚鳳霞楊德國何勇鳳
水產科學 2021年3期
龔森森,吳嘉偉,柴 毅,羅靜波,譚鳳霞,楊德國,何勇鳳
( 1.長江大學 動物科學學院,湖北 荊州 434025; 2.長江大學, 濕地生態與農業利用教育部工程研究中心,湖北 荊州 434025; 3.中國水產科學研究院 長江水產研究所, 農業農村部淡水生物多樣性保護重點實驗室,湖北 武漢 430223 )
浮游動物是水域生態系統中的初級消費者,也是高營養級水生動物幼體的主要開口餌料以及動物性濾食性魚類的主要食物,能有效調控浮游植物數量,降低水體初級生產力,在水域生態系統的結構功能和生物生產力的研究中具有重要意義[1-3]。浮游動物通常被認為是水質評價的重要指示生物,其種類組成、優勢種、豐度及生物量等群落結構特征可以反映出水環境變化狀況,預測其變化趨勢[4]。目前,浮游動物已被廣泛應用于湖泊、水庫等內陸水域次級生產力的評估以及水環境的監測和評價,對水域生態系統的可持續發展利用具有重要意義[5-6]。
長湖是湖北省第三大天然淡水湖泊,是長江中游地區重要濕地生態系統,2010年被批準建立國家級鲌類種質資源自然保護區,具有蓄洪、灌溉、養殖和航運等綜合功能[7-8]。近年來,引江入漢、蒙華鐵路和318國道長湖特大橋等涉水工程的陸續建設對該水域產生不同程度的影響。多種評價方法綜合分析表明,自2007年以來,長湖水質狀況一直處于中度—重度富營養化狀態[9-10]。目前圍繞長湖的研究主要涉及浮游植物、水質監測、高等水生植物以及魚類資源調查等方面[11-13],其中關于浮游動物的研究僅見于2012年[14]。筆者闡明浮游動物季節演替規律并探討與環境因子的關系,為長湖水生生物資源的可持續發展與利用提供重要參考,并為下一步生態系統的保護、恢復與修復提供科學參考依據。
1 研究方法
1.1 區域概況及采樣點的設置
1.1.1 區域概況
長湖位于長江和漢江之間,地跨荊州、荊門和潛江,屬于北亞熱帶濕潤季風氣候,年降雨量約1200 mm,平均水深2.1 m[15]。長湖自西向東可分為廟湖區、海子湖區、馬洪臺湖區和圓心湖區4個區域(圖1)。“引江入漢”工程使廟湖區被2個堤壩(圖1實線隔斷)攔截成靜水區域,故未在此區域設置采樣點。海子湖區狹長,分布有多個進出水口,馬洪臺區和圓心湖區均為開闊水域,其中圓心湖區有較多的湖汊和湖灣[16]。
1.1.2 采樣點設置
依據《水域生態系統觀測規范》[17],共設置14個采樣點:海子湖區4個(H1~H4),馬洪臺區5個(M1~M5),圓心湖區5個(Y1~Y5),采樣點坐標見圖1。2015年4月(春)、7月(夏)、11月(秋)及2016年1月(冬)各采樣1次。
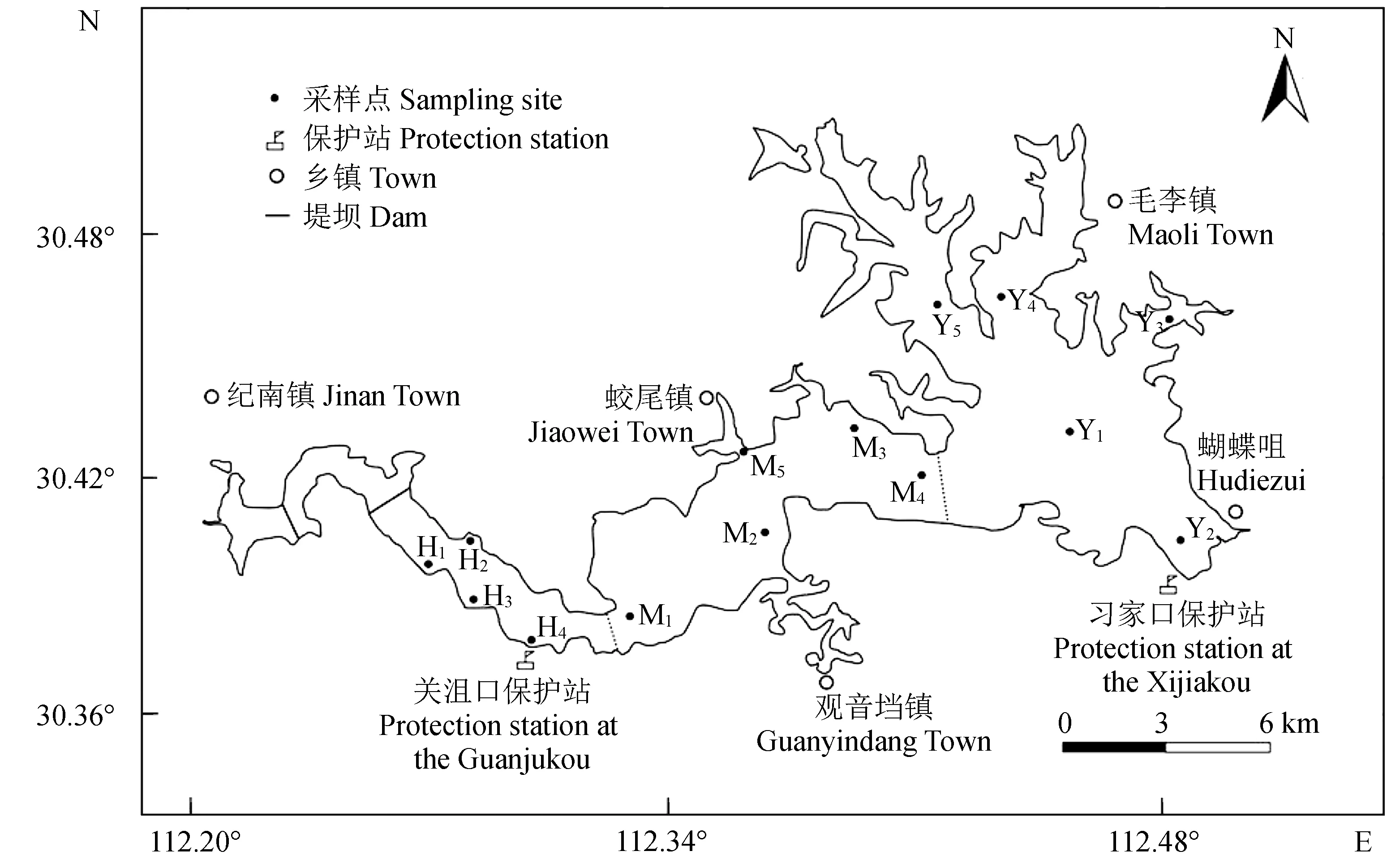
圖1 長湖采樣點示意Fig.1 Sketch map of sampling sites in Changhu Lake
1.2 樣品的采集、處理及分析
1.2.1 樣品采集與處理
參考《水域生態系統觀測規范》和《水生生物學》[18]中樣品采集方法采集浮游生物樣本。參照《中國淡水輪蟲志》[19]、《中國常見淡水枝角類檢索》[20]、《中國動物志》[21]、《淡水微型生物圖譜》[22]鑒定種類;定量計數采用多片計數取平均值的方法,生物量為固定樣本運用體積法[18]得出的濕質量(mg/L)。
1.2.2 水質理化因子
按照國家水質標準方法GB 3838—2002,利用美國HQ40D(Hach Loveland)多功能水質分析儀測定并記錄各采樣點溶解氧含量、pH和水溫,用黑白盤測定透明度,水樣帶回實驗室測定總磷、總氮、氨氮、硝態氮、亞硝態氮、磷酸鹽和總懸浮物含量等水質參數。
1.3 數據處理
1.3.1 優勢度
用優勢度指數(Y)確定優勢種,Y>0.02即為優勢種。
Y=(ni/N)×fi
(1)
式中,ni為浮游動物群落中第i種物種的豐度,N是群落中物種的總豐度,fi為第i種浮游動物在各站位的出現頻率[23]。
1.3.2 物種更替率
物種更替率(R)按下式計算:
(2)
式中,a和b分別表示相鄰兩個季節的物種的種類數,c為兩個季節共有物種的種類數[24]。
1.3.3 排序及分析
排序及分析運用R 3.5.3軟件的Rstudio窗口完成。對浮游動物優勢種與環境因子進行冗余分析,對各季節不同站位的生物量、豐度及物種數等生物指標進行非度量多維尺度分析,對優勢種的各項生物指標進行聚類分析。參考《數量生態學—R語言的應用》[25]使用R語言軟件。
2 結 果
2.1 浮游動物種類組成
2.1.1 種類組成
本次調查共鑒定出浮游動物3門34屬61種(表1)。其中輪蟲13屬29種(占總數的47.54%),原生動物9屬15種(占比24.59%),枝角類7屬11種(占比18.03%),橈足類6屬6種(占比9.84%)。夏季種類數最多,為43種,其次是春季,為37種,秋、冬季節分別為20種和19種。

表1 長湖浮游動物種類組成Tab.1 The species composition of zooplankton in Changhu Lake
2.1.2 豐度和生物量
各類群浮游動物在各區域的豐度變化見圖2,其變化特征為夏(6781 個/L)>秋(2912 個/L)>春(1944 個/L)>冬(1359 個/L),季節性差異顯著(單因素方差分析,F=17.910,P<0.001)。其中輪蟲占絕對優勢,最高達89.12%(夏季)。從各區域豐度分布來看,海子湖區浮游動物豐度在各季節均最高。
各季節平均生物量整體表現為夏(17.78 mg/L)>秋(15.45 mg/L)>春(5.07 mg/L)>冬(2.34 mg/L),季節性差異顯著(單因素方差分析,F=8.443,P<0.001)(圖3)。其中輪蟲生物量在春、夏、秋3個季節當中均為最高,最高達13.99 mg/L(秋季)。從各區域分布來看,海子湖區平均生物量在春、夏、冬3個季節中均為最高。
2.2 優勢種
利用優勢度指數共篩選出12種浮游動物優勢種(表2),包括8種輪蟲和4種原生動物,其中萼花臂尾輪蟲和針簇多枝輪蟲為4季共有優勢種,二者在各采樣點的出現頻率均為100%;由優勢度指數可知,針簇多枝輪蟲在4個季節的優勢度均為最高,其中冬季達0.657。

圖2 各采樣區域浮游動物豐度的季節變化Fig.2 Seasonal changes in zooplankton abundance in sampling sitesH.海子湖區域; M.馬洪臺區域; Y.圓心湖區域; 下同. H.Haizi Lake area; M.Mahongtai area; Y.Yuanxin Lake area;et sequentia.

圖3 各采樣區域浮游動物生物量的季節變化Fig.3 Seasonal changes in zooplankton biomass in sampling sites in various seasons
2.3 物種更替率
統計相鄰2個季節的總物種和共有物種,利用物種更替率公式計算出各類群物種更替率(表3)。總體來看,平均更替率呈現橈足類(80.01%)>輪蟲(75.18%)>原生動物(74.66%)>枝角類(46.88%)。其中原生動物秋—冬季更替率最高(90.00%),枝角類冬—春季更替率最低(25.00%)。

表2 長湖浮游動物優勢種Tab.1 Dominant species of zooplankton in Changhu Lake
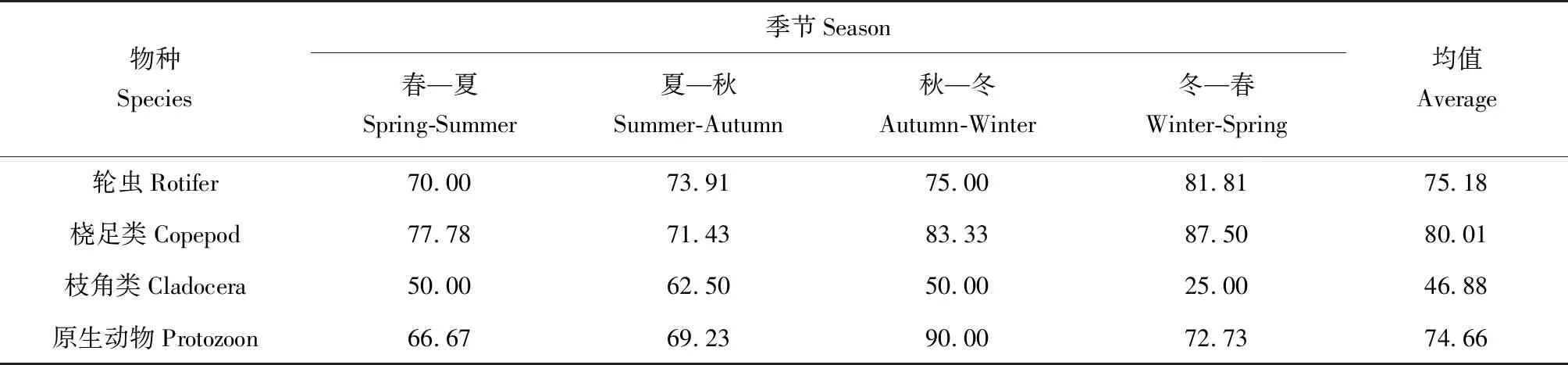
表3 長湖各類群物種更替率季節性變化 %Tab.2 The seasonal variation in species succession rate of various zooplanktons in Changhu Lake
2.4 相似性分析
以4個季節采樣點為分類單元,以各樣點在各季節的浮游動物豐度及生物量為依據進行非度量多維尺度分析,排序分析的平均脅強系數(Stress)為0.092∈(0.05,0.10),表明其結果可信[26]。各季節不同采樣點的浮游動物豐度及生物量聚類分布差異明顯,較好地表現出其季節性差異;其中各分類單元在冬季很好地聚為一類,與其他3個季節有明顯的差異,而春季分類單元的分布較零散,且與夏、秋季個別分類單元表現出一定的相似性(圖4)。
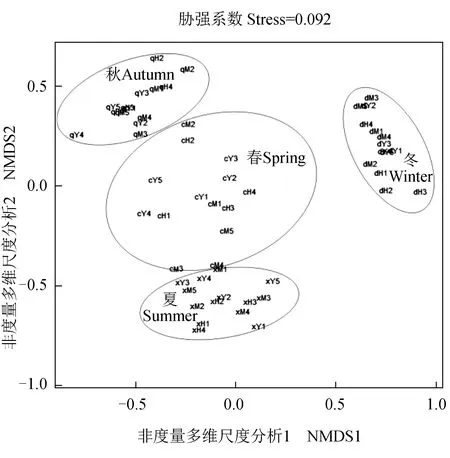
圖4 長湖各樣點非度量多維尺度分析Fig.4 Non-metric multi-dimensional scale analysis of various sampling site in Changhu Lake
以所有優勢種為分類單元進行層次聚類分析。先計算Bray-curtis相異系數(取值0~1,越接近于1表明樣本間浮游動物群落組成差異越大),再計算同表型距離及Gower距離對聚類方法進行篩選。結果顯示,算術平均的非權重成對組聚類法最優;利用輪廓寬度圖對最佳聚類簇數進行篩選,最終將優勢種聚類樹劃分為4個簇,劃分簇的Height值約為0.81(圖5)。聚類結果表明,各分類簇中卜氏晶囊輪蟲和緣板龜甲輪蟲的時空分布特征最為相似,萼花臂尾輪蟲和針簇多肢輪蟲次之。
2.5 相關性分析
選取各季節水環境因子與對應季節優勢種進行相關性排序分析。先將各季節優勢種的原始統計數進行去趨勢對應分析,結果顯示,4個季節排序前4個軸中最大值依次是1.07、0.64、1.43和0.85,均小于3.0,選做冗余分析。再將各季節水質因子與優勢種統計數經lg (X+1)轉換(pH除外)后進行冗余分析,獲得4個季節環境因子與優勢種的相關雙序圖(圖6)。圖中箭頭表示環境因子,“Δ”表示優勢種,箭頭連線的長短以及物種到原點連線與環境因子夾角的余弦值表示物種分布與環境因子相關性的大小。

圖5 長湖浮游動物優勢種聚類分析Fig.5 Cluster analysis of dominant species of zooplankton in Changhu Lake X1~X12.優勢種代碼(表2). X1—X12.dominant species code(Tab.2).
各類群浮游動物優勢種與環境因子表現出不同程度的相關性。夏季原生動物中湖生累枝蟲、尖頂砂殼蟲和似鐘蟲均與總氮含量呈較明顯的正相關性。春季萼花臂尾輪蟲與硝態氮含量和透明度呈正相關關系,針簇多肢輪蟲與磷酸鹽含量也表現出正相關,夏季萼花臂尾輪蟲與總氮以及針簇多肢輪蟲與總磷均有較明顯的正相關,秋季蓋氏晶囊輪蟲與pH和水溫、萼花臂尾輪蟲和針簇多肢輪蟲與總磷含量也呈正相關。冬季萼花臂尾輪蟲、螺形龜甲輪蟲、針簇多肢輪蟲與溶解氧含量呈顯著正相關。
3 討 論
3.1 浮游動物群落結構特征
作為水域生態系統食物鏈中的初級消費者,浮游動物同時受到生產者(主要是浮游植物)和次級消費者[主要是水生動物幼體和鳙魚(Aristichthysnobilis)等動物性濾食性魚類]的影響,其中浮游植物的種類組成、豐度和生物量等特征可直接影響浮游動物的組成和分布等群落特征。在本研究中,浮游動物的種類組成為輪蟲占比最高,總種類數為夏季最多。與同期浮游植物調查結果[27]比較發現,長湖屬于典型的以綠藻門(占比50.8%)為主的水域類型,且種類數量也是夏季最多,這表明長湖浮游植物和浮游動物具有相似的季節分布和變化規律。還有研究表明,輪蟲分布廣,環境適應性強,尤其在富營養化淺水湖泊中,原生動物和輪蟲通常是浮游動物的主要組成部分[28]。本次調查中原生動物和輪蟲占比超過70%,占據了浮游動物的主要組成部分,尤其是輪蟲的豐度和生物量均占絕對優勢,符合輪蟲在富營養化水域環境中的分布規律。

圖6 各季節長湖內浮游動物優勢種與對應環境因子的冗余排序Fig.6 RDA sequencing diagram between dominant species of zooplankton and corresponding environmental factors in Changhu Lake in each season DO.溶解氧; T.水溫; TP.總磷; TN.總氮; NH4+-N.氨氮; NO3--N.硝態氮; NO2--N.亞硝態氮; PO43--P.磷酸鹽; SD.透明度; TSS.總懸浮物; X1~X12為優勢種代碼(表2). DO.dissolved oxygen level; T.water temperature; TP.total phosphorus level; TN.total nitrogen level; NH4+-N.ammonia nitrogen level; NO3--N.nitrate nitrogen level; NO2--N.nitrite nitrogen level; PO43--P.phosphate level; SD.Secchi disk transparency; TSS.total suspended material level; X1—X12.dominant species code(Tab.2).
除浮游植物外,食物鏈中的次級消費者(主要是魚類)對浮游動物牧食壓力也會影響其群落結構特征。王亞龍等[8]在2014年對長湖魚類進行資源調查發現,長湖魚類資源已出現衰退,以湖泊定居型為主,優勢魚類以小型魚類[鯽魚(Carassiusauratus)、(Hemiculterleucisculus)、麥穗魚(Pseudorasboraparva)、棒花魚(Abbottinarivularis)、小黃黝魚(Micropercopsswinhonis)、達氏鲌(Culterdabryi)等]或大型魚類[鰱魚(Hypophthalmichthysmolitrix)、鳙魚、鲇魚(Silurusasotus)、鯉魚(Cyprinuscarpio)等]的幼體為主,而幼魚和濾食性魚類對浮游動物的攝食需求較大,也會影響其群落組成、豐度和生物量等,且具有較明顯的季節性變化。
3.2 浮游動物優勢種
本次調查優勢種共8種輪蟲和4種原生動物。其中,萼花臂尾輪蟲和針簇多肢輪蟲在4個季節的出現頻率均為100%;夏季出現頻率為100%的種類還有前節晶囊輪蟲、湖生累枝蟲和尖頂砂殼蟲;秋季出現頻率為100%還有蓋氏晶囊輪蟲;冬季還有螺形龜甲輪蟲。這些種類均為富營養化水體常見種,一般屬廣溫性種類,分布廣,適應性強。從優勢度指數來看,針簇多肢輪蟲在4個季節的優勢度指數均為最高,不同季節優勢度指數較高的種類還有:夏季萼花臂尾輪蟲(0.184),秋季蓋氏晶囊輪蟲(0.235)、邁氏三肢輪蟲(0.111),冬季螺形龜甲輪蟲(0.114)。這些種類均屬于典型浮游種類,水平分布和垂直分布均較廣。據報道,營浮游生活的典型輪蟲的形態往往有明顯的季節變異現象[29]。本次調查鑒定中也發現,螺形龜甲輪蟲夏季類型的棘突明顯較冬季類型長且明顯,這也是輪蟲適應環境的一種變異。
物種聚類和排序的結果反映了物種分布和數量,非度量多維尺度分析,被認為是研究生態學較好的方法之一。本次對優勢種的聚類分析結果表明,卜氏晶囊輪蟲和緣板龜甲輪蟲聚為一簇且Height值較高,萼花臂尾輪蟲和針簇多肢輪蟲次之,表明了同簇內物種占據相似的生態位或對生境有相同或相似的需求。本次各季節不同采樣點(以對應季節該樣點浮游動物為依據)的非度量多維尺度分析二維點陣排序圖脅強系數小于0.1,較好地體現了不同季節各采樣點物種的關系。不同季節各采樣點均能較好地聚為一類,這充分體現了浮游動物分布的季節差異性。夏季主要以耐高溫、高營養鹽的種類居多,而冬季則以耐低溫種類為主,春、秋季則由于溫度、光照度等因素的相似性而出現部分重疊,也表現出物種對生境有相同或相似的需求,而相同季節各采樣點(豐度和生物量)之間的分化不明顯。
3.3 與環境因子的相關性分析
除受到食物供給、捕食、競爭等生物因素影響外,浮游動物的群落結構特征也與水環境因子等非生物因素緊密相關。冗余分析排序可以分析物種與環境因子的關系,反映出物種對環境的需求程度[30]。從本次浮游動物優勢種與環境因子的冗余分析可以看出,總氮、總磷、硝態氮、磷酸鹽、總懸浮物以及pH等是影響浮游動物群落變化的重要環境因子。如夏季前節晶囊輪蟲、湖生累枝蟲、尖頂砂殼蟲和似鐘蟲均與總氮含量表現出較明顯的正相關性;春季萼花臂尾輪蟲與硝態氮含量以及秋季與總磷含量均呈明顯的正相關關系。姚佳玫等[31]對富春江的調查發現,在一定范圍內淡水水體中輪蟲種類組成和數量、密度與pH呈正相關,一般表現為:pH升高輪蟲數量和密度也會增加。此外總磷還是評價水體富營養化程度的重要指標——尤其淺水湖泊中,富營養化程度隨著總磷水平的升高而加劇,浮游植物的密度和生物量也隨之升高,這也影響輪蟲群落結構。一些原生動物和輪蟲對水體的營養條件有偏好性,如鞭毛蟲類和臂尾輪蟲明顯偏好富營養化水體,有柄的纖毛蟲類如小口鐘蟲(V.microstoma)和褶累枝蟲(E.plicatilis)等通常作為污水處理的指標生物。如本次調查中的優勢種萼花臂尾輪蟲、湖生累枝蟲、似鐘蟲和螺形龜甲輪蟲均是富營養化湖泊和池塘的常見種,而尖頂砂殼蟲和王氏擬鈴殼蟲則為寡污帶常見浮游原生動物,因此以原生動物和輪蟲為環境指示種和水環境狀況評價指標。
大量研究結果表明,水溫也是對水生生物生長發育及群落特征影響較大的環境因子[32-35]。本研究中,長湖浮游動物豐度、生物量及物種豐富度從春季到夏季皆隨溫度的升高而上升,從夏季到秋季再到冬季則相反,冗余分析結果與該直觀變量的變化結果相一致。筆者還發現,輪蟲類群的豐度受水溫、溶解氧及pH的影響較大,營養鹽也對其有一定的影響。原生動物豐度除了受到水溫、溶解氧、pH的限制以外,還受懸浮物、總氮及總磷等指標的影響。長湖的地形地貌及其他生物和非生物因素如水流、水位、降雨量、水草種類與分布等也會一定程度影響浮游動物。
3.4 長湖浮游動物演替特征
從本次調查中季節更替率來看,除枝角類冬—春季節物種更替率較低(25.00%)外,其余各種浮游動物的季節更替率均超過50%,其中超過80%的有秋—冬季原生動物(90.00%)、冬—春季輪蟲(81.81%)、秋—冬季橈足類(83.33%)和冬—春季橈足類(87.50%),表明浮游動物各類群季節變化明顯,這與浮游植物季節性變化規律密切相關。有研究發現[36],夏季浮游植物以喜高溫的藍藻和綠藻門種類為主,尤其以具有假空泡的藍藻門占優勢,早春、晚秋以及冬季均以耐低溫的金藻和硅藻種類為主。因此長湖浮游動物的種群更替率也受同期浮游植物的種類組成、豐度和生物量等因素影響。
本次調查是繼郭坤等[14]之后第二次對長湖浮游動物的系統研究,并增加了對原生動物的種類鑒定。這兩次調查結果比較發現,長湖浮游動物種類多樣性降低,豐度升高(輪蟲種類數42種減少為29種,枝角類16種減少為7種,夏季平均豐度4412 個/L升高為6781 個/L等)。這表明長湖水域生態系統的穩定性繼續減弱,水體富營養化程度進一步加深。另一方面,優勢種仍以浮游輪蟲為主,且大部分屬富營養化水體的指示種。優勢種增加了4種原生動物,其中湖生累枝蟲和似鐘蟲也偏好多污帶或中污帶水體,這些都表明長湖水域仍處于富營養化狀態,這也與本研究中用浮游動物豐度、生物量以及水環境因子對水質的評價結果相一致;此外,該結果還與魏志兵等[37]對同時期長湖浮游植物的研究所得結論相符。本研究除了增加了原生動物種類外,還對優勢種和各季節采樣點進行了聚類分析,較好地體現了浮游動物優勢種以及各采樣點在不同季節的關系,較2012年的調查又有了更深入的分析。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
我愛學·笑話與口才(2025年3期)2025-02-24 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
趣味(語文)(2020年3期)2020-07-27 01:42:46
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
中國衛生(2015年2期)2015-11-12 13:13:54
少兒科學周刊·少年版(2015年1期)2015-07-07 17:15:12
