蘇北灘涂鹽堿地3種典型鹽生植物根際土壤細菌多樣性及群落結構分析
2021-05-26 19:52:40董靜邢錦城溫祝桂洪立洲劉沖朱小梅趙寶泉賀亭亭趙小慧
江蘇農業科學 2021年8期
關鍵詞:群落結構
董靜 邢錦城 溫祝桂 洪立洲 劉沖 朱小梅 趙寶泉 賀亭亭 趙小慧
摘要:以蘇北灘涂鹽堿地3種典型鹽生植物的非根際與根際土壤為研究對象,利用高通量測序技術對土壤細菌多樣性特征及群落結構進行分析,并對土壤理化性質進行測定。結果表明:鹽地堿蓬、鹽角草和二色補血草根際土壤細菌群落分別由15、16、17門組成,其中裝甲軍門只出現在二色補血草根際土壤中;非根際土壤主要由18門組成,其中廣古菌門僅出現在鹽地堿蓬非根際土壤中,衣原體門僅出現在二色補血草非根際土壤中。非根際土壤細菌酸桿菌門的相對豐度低于根際土壤,而非根際土壤中的放線菌門豐度有所提高。3種植物根際土壤細菌群落的多樣性與速效氮含量正相關,與土壤pH值、電導率、有機質含量、速效磷含量、有機碳含量及速效鉀含量負相關;土壤電導率、速效氮含量和速效磷含量是影響其根際與非根際土壤細菌群落變化的主要因素。
關鍵詞:鹽生植物;根際微生物;土壤細菌;群落結構;土壤理化性質;蘇北;灘涂鹽堿地
中圖分類號: S182文獻標志碼: A
文章編號:1002-1302(2021)08-0212-07
收稿日期:2020-07-16
基金項目:江蘇省農業科技自主創新資金[編號:CX(19)3096];國家自然科學基金 (編號:31800525);江蘇現代農業產業技術體系項目[編號:JATS(2019)228];蘇北科技專項(編號:SZ-YC2018059、SZ-YC2018060)。
作者簡介:董 靜(1988—),女,江蘇鹽城人,碩士,助理研究員,主要從事植物逆境生理相關研究。E-mail:dongjingyc@163.com。
通信作者:邢錦城,碩士,副研究員,主要從事土壤肥料與鹽土農業工程相關研究。E-mail:sdauxxx@163.com。
鹽生植物在適應鹽堿地環境過程中形成了一系列獨特的耐鹽機制用以應對逆境的影響,除此以外,根際微生物對其耐鹽性的提高也會起到一定的作用[1]。在自然條件下,植物與土壤根際微生物之間聯系緊密,根際微生物種類繁多,作為地球上最復雜的生態系統之一,它們對植物的生長和健康有重要影響[2]。植物可以通過根系代謝活動來改變根際微生物群落的組成,同時根際土壤也會受到其活動的影響,最終導致根際與非根際土壤理化性質及微生物群落結構存在顯著差異[3-5]。因此,研究不同鹽生植物對土壤細菌群落結構及多樣性的影響,揭示植物耐鹽堿性與土壤微生物之間的內在聯系,具有重大意義。
土壤微生物對保持生物多樣性以及維護生態系統功能均有著重要的作用[6]。近年來,鹽堿區域土壤、植物及其根際微生物相互作用、鹽堿地生態環境可持續發展等方面引起了國內外學者的重視[7-10]。我國蘇北沿海灘涂面積廣、鹽分重,嚴重限制了植物的生存與分布,鹽地堿蓬(Suaeda salsa)、鹽角草(Salicornia europaea)和二色補血草(Limonium bicolor)是3種廣泛分布于蘇北沿海灘涂鹽堿地的典型鹽生植物,均具有極強的耐鹽堿性,是適合鹽漬地修復的類群。為研究鹽生植物抗逆性及環境適應機制,本研究通過高通量測序對幾種灘涂典型鹽生植物的非根際與根際土壤細菌群落結構進行探討,分析土樣中細菌類群多樣性和優勢種群及其與環境因子的相關性,以期為揭示根際微生物對鹽生植物耐鹽堿性的作用提供理論依據。
1 材料與方法
1.1 材料
土壤樣品于2019年5月21日采自江蘇黃海濕地大豐麋鹿國家級自然保護區的鹽堿地,該地屬于亞熱帶季風氣候,降雨主要集中在6—8月。分別在鹽地堿蓬(JP)、鹽角草(HP)和二色補血草(BX)居群中選擇長勢相同的植株作為樣本,每種植物采集15株作為重復。完整挖出植株根系,采集根部附著的與根系結合較為緊密的土壤(1 mm),混合均勻,作為根際土壤;同時撇去表層土,采集距離根系0~30 cm的土壤,作為非根際土壤,所取土樣均裝入無菌離心管中,置于冰盒中帶回實驗室,4 ℃暫存待測。
1.2 方法
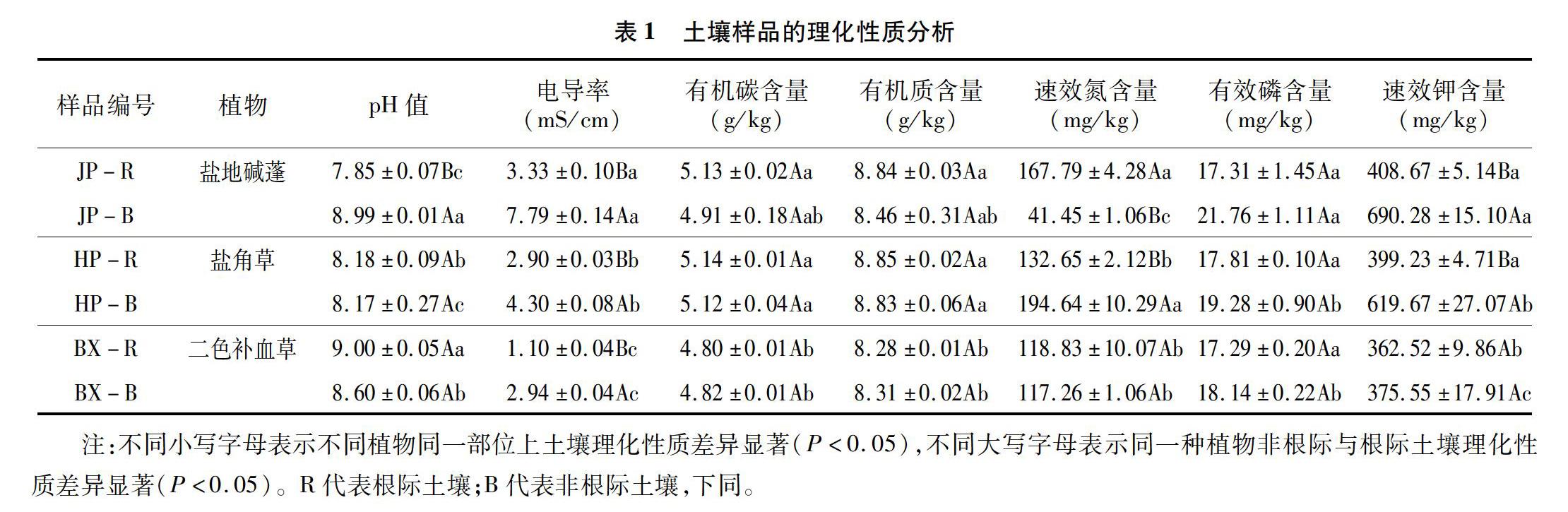
土壤樣品的理化性質測定指標包括土壤pH值、有機碳(TOC)含量、電導率(EC)、有機質(SOM)含量、速效氮(AN)含量、有效磷(AP)含量、速效鉀(AK)含量[11-12]。
按照DNA提取試劑盒(QIAGEN)操作步驟提取土壤樣品DNA。參照劉國麗等的方法[12],由微分(上海)基因科技有限公司進行DNA文庫構建及測序。
通過QIIME軟件計算α多樣性指數。利用UniFrac計算樣本距離并繪制聚類樹。通過典范對應分析(CCA)篩選出對細菌群落結構產生影響的主要環境因子。
2 結果與分析
2.1 不同土壤理化參數
由表1可知,3種鹽生植物之間的pH值、EC值以及TOC、SOM、AN、AP、AK含量在不同植物的非根際及根際土壤中均有差異。除鹽地堿蓬根際土壤pH值顯著小于非根際土壤外(P<0.05),其余2種植物中均無顯著差異。3種植物根際土壤的EC值均顯著小于非根際土壤(P<0.05)。TOC及SOM在根際與非根際土壤之間均無顯著差異,其中二色補血草根際土壤中TOC與SOM含量均顯著高于其余2種植物。鹽地堿蓬非根際AN含量顯著低于根際土壤,而其根際土壤AN含量顯著高于鹽角草、二色補血草根際土壤。非根際土壤AP與AK含量均高于根際土壤,其中鹽地堿蓬和鹽角草非根際土壤AK含量均顯著高于根際土壤。
2.2 土壤細菌多樣性及群落結構分析
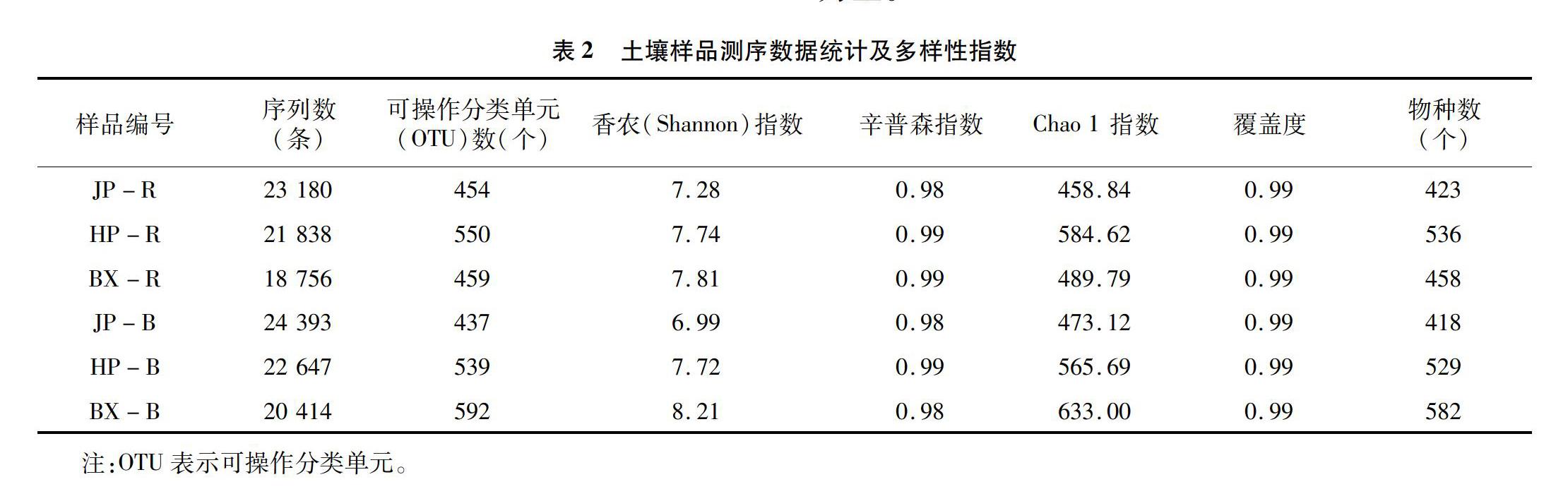
2.2.1 測序數據分析
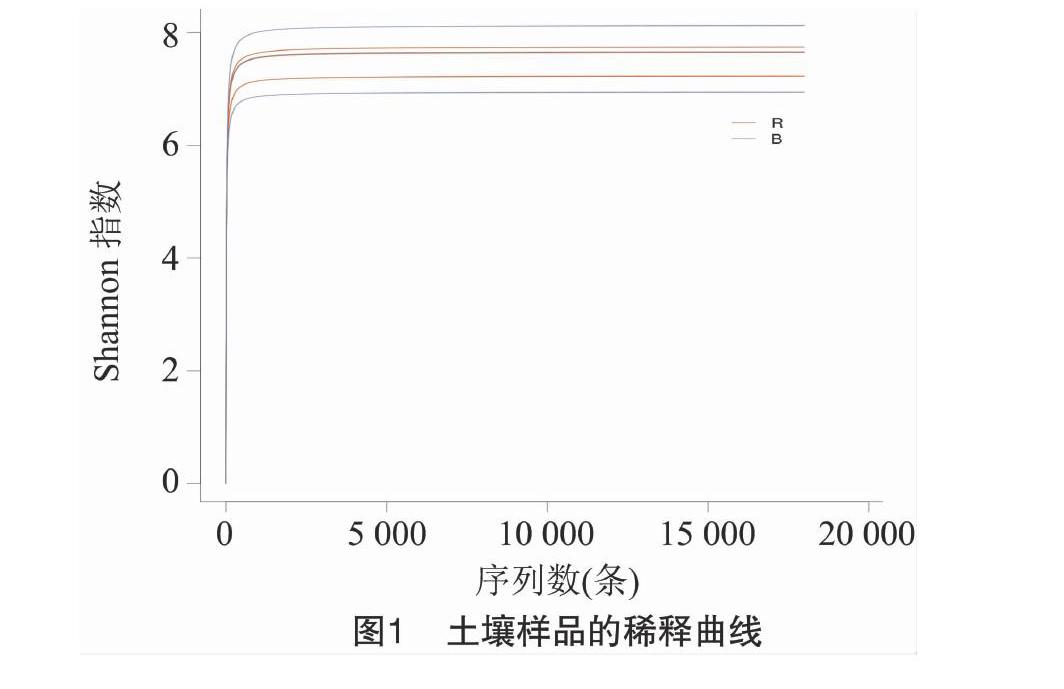
所得有效序列平均長度在460 bp以上。圖1表明,隨著測序深度的增加,樣品稀釋曲線基本趨于平緩,并且測序覆蓋度均為0.99(表2),綜上表明本次測序結果較為合理。
2.2.2 細菌多樣性
從表2可以看出,JP和HP根際土壤的細菌群落的香農指數均高于非根際土壤。BX根際土壤細菌群落的多樣性指數明顯高于JP,其Chao 1指數與物種數均明顯低于HP,這表明HP根際土壤的細菌群落豐度高于BX;BX的非根際土壤的細菌群落多樣性(辛普森指數除外)和豐度均明顯高于其他2種鹽生植物的非根際土壤。總體來看,與JP和HP相比,BX土壤的細菌群落豐富度最高,其微生物種群生態位也最寬。
2.2.3 不同土壤樣品細菌群落結構
從樣品檢測出細菌19門、36綱、49目、73科、189屬。由圖2可知,所有土樣中的細菌主要屬于變形菌門(Proteobacteria)、擬桿菌門(Bacteroidetes)和酸桿菌門(Acidobacteria)3個菌門;這3個門的細菌又分別以丙型變形菌綱(Gammaproteobacteria)、鞘脂桿菌綱(Sphingobacteriia)和α-變形菌綱、δ-變形菌綱為主。
在所有土樣中,擬桿菌門和變形菌門均為優勢門。共有10個優勢門細菌在根際土壤中被檢出,其中鹽地堿蓬根際土壤中檢測出8個,其豐度從大到小的分別為變形菌門(Proteobacteria,62.09%)、擬桿菌門(Bacteroidetes, 23.32%)、酸桿菌門(Acidobacteria,3.85%)、放線菌門(Actinobacteria ,2.27%)、厚壁菌門(Firmicutes,1.94%)、綠彎菌門(Chloroflexi, 1.65%)、浮霉菌門(Planctomycetes, 1.52%)、異常球菌-棲熱菌門(Deinococcus-Thermus,1.21%),占根際土壤微生物總量的88.76%,其中食氫菌門(Hydrogenedentes)等3門的豐度低于0.1%;鹽角草根際土壤中優勢門有Proteobacteria (62.26%)、Bacteroidetes (24.23%)、Acidobacteria (4.67%)、Planctomycetes (2.65%)、Chloroflexi(1.49%)、Actinobacteria (1.42%)和Ignavibacteriae (1.21%)7個門;二色補血草根際土壤優勢細菌群為Proteobacteria(72.68%)、Bacteroidetes (10.90%)、Acidobacteria (5.69%)、Actinobacteria (2.56%)、Planctomycetes (2.51%)、Chloroflexi(2.00%)、疣微菌門(Verrucomicrobia,1.49%),其中Proteobacteria豐度明顯高于其余植物。在非根際土壤中也有10個優勢門被檢出,其中Proteobacteria和Bacteroidetes是最占優勢的2個門,共占非根際土壤細菌豐度的69%以上;在鹽地堿蓬的非根際土壤中的豐度分別為55.66%和29.38%,在鹽角草非根際土壤中豐度分別為65.08%和20.21%,而在二色補血草非根際土壤中分別為56.79%和25.46%。
衣原體門僅出現在二色補血草非根際土壤中,該菌門包括11個屬,其中只有Chlamydia被確認與人或動物的衣原體病相關[13];廣古菌門只在鹽地堿蓬的非根際土壤中檢出,該門包括可在極高鹽濃度下生活的鹽桿菌及海洋類群[14];裝甲軍門只出現在二色補血草根際土壤中,該菌門近年來才被人們所熟知,而其內部系統發育結構的研究尚處于初級階段,目前缺乏一致的研究結論。
從屬水平上看,根際土壤中的優勢屬與非根際土壤相比,數量較多。在鹽地堿蓬、鹽角草和二色補血草根際土壤的細菌優勢屬分別有24、21、27個,而在非根際土中優勢屬有17、22、19個。每種植物根際土中豐度大于10%的菌屬存在明顯差異,其中鹽地堿蓬中為Thiobacillus(12.55%),鹽角草中為Desulfuromonas(22.87%),二色補血草中為Desulfuromonas (10.80%)。在非根際土壤中大于10%的菌屬共有4個,分別為Aliifodinibius、Desulfuromonas、Gracilimonas、Halomonas;同種植被下與非根際土壤相比,Aliifodinibius在根際土壤細菌中的豐度顯著降低(0.06%~2.10%),而一些低豐度屬如Gp10則在根際土壤中的豐度更高,成為優勢屬,表明鹽地堿蓬、鹽角草和二色補血草這3種植物的根際土壤細菌的群落組成和豐度與非根際土壤存在較大的差異。
2.2.4 差異性分析
在本研究中,同一植被的根際和非根際土壤在細菌群落組成及豐富度方面存在明顯差異,不同植物的根際土壤細菌群落亦如此。在門水平上,與非根際土壤相比,Proteobacteri、Acidobacteria、Planctomycetes在根際土壤中的豐度明顯更高;而Bacteroidetes、Actinobacteria的豐度與非根際土壤相比,相對較低(圖3)。Chloroflexi、Planctomycetes豐度在3種植物的根際土壤中存在明顯差異(圖2-A)。
通過比較發現,Desulfuromonas、Fulvivirga、Ignavibacterium、Thioprofundum、Geminicoccus、Erythrobacter、Algoriphagus、Phycisphaera和Gp10為這3種植物根際土壤的共有優勢屬(圖2-C),其中Desulfuromonas在鹽角草根際土壤的豐度(22.87%)明顯高于鹽地堿蓬(1.30%)和二色補血草(10.80%),Thioprofundum、Erythrobacter和Algoriphagus在3種植物根際土壤中的豐度亦如此。Desulfuromonas、Gracilimonas、Marinobacter及Gp21為這3種植物非根際土壤中細菌群落總的共有優勢菌屬。其豐度在鹽地堿蓬非根際土壤中為1.53%~11.94%,在鹽角草非根際土壤中為1.40%~23.87%,而在二色補血草非根際土壤中僅為1.11%~2.75%。綜上可以看出,鹽地堿蓬、鹽角草及二色補血草這3種植物即使在相同的鹽堿環境中,其根際土壤的細菌群落在結構及組成上存在較大差異。
如圖4所示,6個土壤樣品中細菌群落組成總體分為兩大族群,二色補血草非根際土壤為一個族群,其余5個土壤樣品中細菌聚為另一個族群,說明二色補血草非根際土壤與其余土壤細菌群落種群的構成變化較大。鹽地堿蓬、鹽角草以及二色補血草這3種鹽生植物根際土壤的細菌群落間差異相對較小,其中,當取值水平在0.85以上時,這3種鹽生植物根際土壤的細菌群落可聚為一類。
2.2.5 土壤細菌的群落結構與多樣性及其與土壤理化性質相關性
通過分析發現,這6種土壤樣品細菌群落的Shannon指數和Chao1指數與AN含量呈正相關關系,與pH值、EC值、TOC含量、SOM含量 、AP含量及AK含量呈負相關關系;Simpson指數與TOC含量、SOM含量及AN含量呈正相關關系,與pH值、EC值、AP含量及AK含量呈負相關關系,但相關性均未到達顯著水平(表3)。
為進一步探明3種鹽生植物根際土壤理化因子對土壤微生物群落結構的影響,將測序所得的相對豐度位于前50的屬物種數據與土壤理化參數做典范對應分析(CCA),第一、第二排序軸上土壤細菌群落的的解釋量分別為31.4%、41.2%,總排序軸解釋量為72.6%,損失的信息僅有27.4%(圖5),這說明土壤pH值、EC值、TOC、AN和AP含量在研究鹽生植物根際土壤中的重要性。
圖5中第一排序軸與pH值、EC值和AP含量呈負相關,相關系數分別為-0.469、-0.911和 -0.867,這表明從左到右,在橫坐標上pH值、EC值和AP值是逐漸降低的;第一排序軸與土壤TOC和AN含量呈正相關,相關系數分別0.193和0.813,說明土壤TOC含量和AN含量從左到右逐漸升高。通過以上分析發現,EC值、AN含量和AP含量是影響3種鹽生植物根際和非根際土壤的細菌群落組成的主要影響因子。
3 討論與結論
本研究以蘇北灘涂鹽堿地3種典型鹽生植物鹽地堿蓬、鹽角草及二色補血草根際與非根際土壤為研究對象,通過對土壤細菌群落多樣性進行調查,發現在3種植物之間的土壤細菌Shannon指數存在差異。其中二色補血草與其他2種植物相比,其根際土壤細菌群落具有更高的多樣性及豐度,這也許是因為二色補血草與其余2種植物相比,其主根直徑較寬較長,并且側根也很發達。根系生長越旺盛的植物,其根際土壤中便會有更多的根系脫落物及分泌物[15],而不同的植被類型,也可能是產生差異化的原因。
植物可從營養代謝角度出發,利用自己的根系活動主動使根際環境發生變化,選擇性地提高或者降低土壤細菌類群的豐度及多樣性,最終形成有益的根際微生物群落結構[16-17]。如芽胞桿菌屬、鞘氨醇單胞菌屬可能促進西洋參的生長[18];大豆苗期土壤中的細菌豐富度和多樣性指數均明顯低于成熟期[19];Acidobacteria、Chloroflexi和Firmicutes等是香榧根際土壤的優勢類群[20]。本試驗中3種鹽生植物根際及非根際土壤的細菌群落組成及多樣性差異顯著,α多樣性分析表明,來自藜科的鹽地堿蓬及鹽角草非根際土壤細菌的多樣性低于根際土壤,這與李巖等的相關研究結果[21-22]一致。而白丹花科的二色補血草正好相反,這與安韶山等對冰草的研究結果[23]相同。在本研究中,除二色補血草非根際土壤微生物Shannon指數明顯大于根際土壤外,其余2種鹽生植物根際Shannon指數均高于非根際土壤,這可能是根系分泌物所引起的根際負效應[24]。
在本研究中,3種鹽生植物根際土壤優勢菌門與非根際土壤組成相似,但比例發生了明顯的變化,多樣性更加豐富。Proteobacteria和Bacteroidetes在所有土壤樣品中均為優勢門,但Proteobacteria的豐度比Bacteroidetes更高,這2個菌門的細菌在我國許多鹽堿土壤中均為優勢菌群[25-27],且在多種植物根際土壤中被列為優勢菌群[28-29]。Proteobacteria被認為是細菌中最大的一菌門,包括很多病原菌和固氮菌,與土壤碳利用有關。Bacteroidetes包括Bacteroidia、Flavobacteria和Sphingobacteriia這3個綱,而Sphingobacteriia在本研究結果中豐度最高,其重要類群Cytophaga在海洋細菌中占據了相當大比例,具有降解纖維素功能[30],這可能與采樣點距離我國江蘇省黃海海岸僅4 km有關。
由聚類分析結果可知,3種植物根際土壤的細菌群落之間的相似性較高,而非根際土壤細菌之間也具有較高的相似性,且根際與非根際之間的相似性低于根際間。這表明在同一種環境中,雖然不同種鹽生植物根際微生物群落有特異性,但是在長期鹽堿脅迫下呈現出了趨同性,即根際細菌群落結構在不同的鹽生植物之間相似,這也許是鹽生植物的根際效應在起作用。相關性分析發現,土壤EC值及AN和AP含量是影響根際土壤微生物群落結構的主要因子,其中AN含量與細菌群落多樣性指數均呈正相關關系,這可能與本研究所取植物樣品均位于同一鹽漬環境中有關。土壤EC值與鹽分含量有關,EC值越大,表示可溶性鹽離子濃度越高,說明鹽分含量是影響土壤微生物群落結構的重要因素。有研究表明,土壤有機碳可以直接驅動微生物的組成及多樣性[31],李巖等也認為TOC含量是影響2種鹽生植物根際土壤細菌群落組成的主要影響因子[21],而在本研究中TOC含量與Shannon指數及Chao 1指數呈負相關關系,這可能是因為土壤有機碳含量并不是影響鹽堿地環境中細菌群落多樣性形成的限制因素。
參考文獻:
[1]Qin Y,Druzhinina I S,Pan X Y,et al. Microbially mediated plant salt tolerance and microbiome-based solutions for saline agriculture [J]. Biotechnology Advances,2016,34(7):1245-1259.
[2]Raaijmakers J M,Paulitz T C,Steinberg C,et al. The rhizosphere:a playground and battlefield for soilborne pathogens and beneficial microorganisms[J]. Plant and Soil,2009,321:341-361.
[3]Bakker M G,Chaparro J M,Manter D K,et al. Impactsofbulk soil microbial community structure on rhizosphere microbiomes of Zea mays[J]. Plant and Soil,2015,392:115-126.
[4]Edwards J,Johnson C,Santos-Medellin C,et al. Structure,variation,and assembly oftheroot-associated microbiomes of rice[J]. Proceedings of the National Academy of Sciences of the United States of America,2015,112(8):E911-E920.
[5]李森森,馬大龍,臧淑英,等. 不同干擾方式下松江濕地土壤微生物群落結構和功能特征[J]. 生態學報,2018,38(22):7979-7989.
[6]畢江濤,賀達漢.植物對土壤微生物多樣性的影響研究進展[J]. 中國農學通報,2009,25(9):244-250.
[7]李巖,何學敏,楊曉東,等. 不同生境黑果枸杞根際與非根際土壤微生物群落多樣性[J]. 生態學報,2018,38(17):5983-5995.
[8]Yang J,Ma L A,Jiang H C,et al. Salinity shapes microbial diversity and community structure in surface sediments of the Qinghai-Tibetan Lakes[J]. Scientific Reports,2016,6:25078.
[9]Borruso L,Bacci G,Mengoni A,et al. Rhizosphere effect and salinity competing to shape microbial communities in Phragmites australis (Cav.) Trin. ex Steud[J]. FEMS Microbiology Letters,2014,359(2):193-200.
[10]李 新,焦 燕,代 鋼,等. 內蒙古河套灌區不同鹽堿程度的土壤細菌群落多樣性[J]. 中國環境科學,2016,36(1):249-260.
[11]鮑士旦. 土壤農化分析[M]. 3版. 北京:中國農業出版社,2000:14-20.
[12]劉國麗,牛世偉,徐嘉翼,等. 基于高通量測序分析優化施氮對養蟹稻田土壤細菌多樣性的影響[J]. 吉林農業大學學報,2019,42(6):686-694.
[13]李 鵬,端 青,宋立華. 衣原體最新分類體系與分類鑒定方法研究進展[J]. 中國人獸共患病學報,2014,30(12):1262-1266.
[14]賀紀正,沈菊培,張麗梅.土壤中溫泉古菌研究進展[J]. 生態學報,2009,29(9):5047-5055.
[15]曹永昌,楊 瑞,劉 帥,等. 秦嶺典型林分夏秋兩季根際與非根際土壤微生物群落結構[J]. 生態學報,2017,37(5):1667-1676.
[16]張太平,潘偉斌. 根際環境與土壤污染的植物修復研究進展[J]. 生態環境,2003,12(1):76-80.
[17]原紅娟,朱紅惠. 不同植被根際土壤中可培養與未培養黏細菌的多樣性研究[J]. 江蘇農業科學,2020,48(7):264-269.
[18]李 麗,蔣景龍 .基于高通量測序的西洋參根際土壤細菌群落分析[J]. 中藥材,2019,42(1):7-12.
[19]劉 欣,李志英,劉瑞瑞,等. 大豆不同生育期根際土壤細菌群落結構的變化[J]. 廣西植物,2018,38(10):1363-1370.
[20]葉 雯,李永春,喻衛武,等. 不同種植年限香榧根際土壤微生物多樣性[J]. 應用生態學報,2018,29(11):3783-3792.
[21]李 巖,楊曉東,秦 璐,等. 兩種鹽生植物根際土壤細菌的多樣性和群落結構[J]. 生態學報,2018,38(9):1-14.
[22]Cui H Y,Yang X Y,Lu D X,et al. Isolation and characterization of bacteria from the rhizosphere and bulk soil of Stellera chamaejasme L.[J]. Canadian Journal of Microbiology,2015,61(3):171-181.
[23]安韶山,李國輝,陳利頂. 寧南山區典型植物根際與非根際土壤微生物功能多樣性[J]. 生態學報,2011,31(18):5225-5234.
[24]吳林坤,林向民,林文雄. 根系分泌物介導下植物-土壤-微生物互作關系研究進展與展望[J]. 植物生態學報,2014,38(3):298-310.
[25]林耀奔,楊建輝,葉艷妹. 鹽堿地不同土地利用方式下土壤細菌群落結構多樣性差異分析[J]. 環境科學學報,2019,39(4):1266-1273.
[26]李 新,焦 燕,楊銘德,等. 內蒙古河套灌區不同鹽堿程度的土壤細菌群落多樣性[J]. 中國環境科學,2016,36(1):249-260.
[27]Dai J Y,Tang X M,Gao G,et al. Effects of salinity and nutrients on sedimentary bacterial communities in oligosaline Lake Bosten,northwestern China[J]. Aquatic Microbial Ecology,2013,69:123-134.
[28]Yang H,Hu J X,Long X H,et al. Salinity altered root distribution and increased diversity of bacterial communities in the rhizosphere soil of Jerusalem artichoke[J]. Scientific Reports, 2016,6(1):20687.
[29]楊美玲,張 霞,王紹明,等. 基于高通量測序的裕民紅花根際土壤細菌群落特征分析[J]. 微生物學通報,2018,45(11):2429-2438.
[30]Liu J J,Sui Y Y,Yu Z H,et al. High throughput sequencing analysis of biogeographical distribution of bacterial communities in the black soils of northeast China[J]. Soil Biology and Biochemistry,2014,70:113-122.
[31]Delgado-Baquerizo M,Maestre F T,Reich P B,et al. Carbon content and climate variability drive global soil bacterial diversity patterns[J]. Ecological Monographs,2016,86(3):373-390.
猜你喜歡
熱帶農業科學(2016年11期)2017-01-21 14:59:21
熱帶農業科學(2016年11期)2017-01-21 14:57:08
河北漁業(2015年12期)2015-12-21 14:37:31
河北漁業(2015年11期)2015-11-18 16:39:54
河北漁業(2015年11期)2015-11-18 16:37:49
河北漁業(2015年10期)2015-10-15 11:14:57
湖北農業科學(2015年18期)2015-10-13 21:34:01
河北漁業(2015年7期)2015-07-21 11:06:52
河北漁業(2015年6期)2015-06-24 11:04:37
安徽農學通報(2015年5期)2015-03-23 23:51:20
