紫花苜蓿個體性狀對土壤磷素供給能力的響應研究
2021-05-27 03:09:04郭豐輝丁勇馬文靜李西良張勇師尚禮侯向陽
草原與草坪 2021年1期
郭豐輝,丁勇,馬文靜,李西良,張勇,師尚禮,侯向陽
(1.甘肅農業大學 草業學院/草業生態系統教育部重點實驗室/甘肅省草業工程實驗室/中-美草地畜牧業可持續發展研究中心,甘肅 蘭州 730070;2.中國農業科學院草原研究所,內蒙古 呼和浩特 010010;3.內蒙古民族大學,內蒙古 通遼 028000)
紫花苜蓿(Medicagosativa)作為豆科植物,其根系可與固氮菌共生形成根瘤,具有較強的固氮能力。苜蓿每年固氮量為220~670 kg/hm2,相當于470~1 450 kg的尿素[1]。而磷元素作為植物生長所必需的大量元素之一,由于其移動性差,易被固定,有效性低,往往成為制約植物生長的主要因素[2]。有研究表明,相對于氮鉀元素,我國苜蓿生產更易受到土壤磷元素匱乏的限制[3-5]。磷素限制不僅影響了苜蓿干草生產量及種子產量[6-8],而且降低了苜蓿養分、粗蛋白含量,降低了苜蓿草產品的品質及適口性[9]。土壤磷素的供給能力直接關系到苜蓿的越冬率及抗逆性,施用磷肥可以明顯提高苜蓿春季返青率,增加鹽漬苜蓿田的幼苗建植率及產量[10-11]。苜蓿根系與固氮菌的共生關系受到土壤磷素供給能力的影響,磷素限制會減少有效根瘤數量,進而減小單位面積苜蓿的固氮量,最終降低苜蓿固氮能力[12-13]。鑒于土壤磷素在苜蓿生產中的重要作用及我國苜蓿主產區土壤磷元素可利用性低的特點,有關磷素與苜蓿生長及生產的相關性研究一直是草學研究領域的熱點之一。
苜蓿生物量的形成與其形態發育密切相關,影響苜蓿產草量的主要形態性狀有單莖重、莖長、莖粗、側枝數、葉片數等[14]。據報道苜蓿葉片性狀及分枝,包括葉面積指數(LAI)、冠層結構、葉長、葉寬、葉角度、堅挺小葉、分枝數目(株數)等與植株的光攔截有關,是影響苜蓿產量及質量的主要形態性狀[15]。不同基因型苜蓿單株間生物量的差異受到其形態及側枝性狀的調控,在此調控過程中,側枝數量大于單個側枝重量的貢獻,而葉長的貢獻大于葉寬[16]。磷肥的施用可以通過增大苜蓿株高、增加側枝數量提高苜蓿單株生物量,進而提高苜蓿產量[17-18];但是有研究表明磷肥添加并沒有改變苜蓿的植株密度和側枝數量,其通過提高單個側枝重量對苜蓿生物量產生影響[19-20]。因此,改變單株生物量是土壤磷素供給能力改變苜蓿生產力的重要途徑,但是有關土壤磷素供給能力通過改變苜蓿形態性狀(葉片性狀、側枝性狀等)影響苜蓿單株生物量尚存在爭議。盡管國內外針對苜蓿形態特征開展了大量研究,但土壤磷素供給能力通過改變苜蓿形態特征進而影響其單株生物量的途徑尚缺乏足夠的研究。
鑒于土壤磷素供給能力對苜蓿生產的重要性及我國苜蓿主產區土壤磷素現狀,結合前人有關土壤磷素供給能力改變苜蓿生產力途徑的研究,采用室內盆栽試驗方法研究了苜蓿個體性狀對土壤磷素供給能力的響應,擬解決以下科學問題:1)苜蓿單株性狀對土壤磷素供給能力具有什么響應;2)土壤磷素供給能力如何通過莖葉及側枝性狀影響苜蓿單株生物量。
1 材料和方法
1.1 試驗材料
試驗于2017年5月初~7月初在農業部科爾沁牧草資源重點野外科學觀測試驗站溫室進行。為了有效研究土壤中磷元素對天然草原植物所起的作用,避免其他營養成分偏離正常水平而對試驗造成影響,試驗所用土壤采自于中國農業科學院廊坊試驗基地,該土壤有效磷含量長期嚴重缺乏(土壤有效磷(P2O5)含量為3.00 mg/kg,據徐明崗等[21]研究當土壤有效磷(P2O5)含量低于5 mg/kg時即為磷素嚴重匱缺狀態),而其他養分水平相對正常。土壤的基本理化性質為:全磷0.56 g/kg、全氮0.66 g/kg、有效磷(P2O5)3.00 mg/kg、堿解氮51.36 mg/kg、有機質1.11%、pH 7.50。將所有土壤充分混勻、過篩、風干,然后裝入直徑為18 cm的花盆,每盆2.25 kg風干土。供試紫花苜蓿品種為中苜2號,由中國農業科學院草原研究所牧草栽培研究室提供。
1.2 試驗方法
周德超[22]對京郊土壤磷肥添加后固定速率的研究表明,磷肥在添加后一晝夜的固定速率為34%~58%,2~30 d的固定速率為65%~76%。依據該固定速率、本土壤有效磷含量及全國養分含量分級標準[21],所用土壤設置(P2O5)0,30,90,180 mg P2O5/kg 4個磷添加梯度(P0、P1、P2、P3),分別代表很缺乏、中等、豐富、很豐富四個等級,每個處理6個重復,磷肥選用過磷酸鈣。將裝好土的花盆按照其施磷梯度計算好施磷量(每盆含風干土2.25 kg),然后將每盆所添加的磷肥與土壤充分混勻(一次性添足),最后給每盆澆同等量的水(澆透為止)。待土壤潮濕松軟后,每盆挖4個洞穴,每洞穴放3粒種子。出苗15 d后間苗,每盆保留長勢基本相同的幼苗4株,生長期間定期對每盆澆水,保證其水分的充足并及時拔除花盆里生長的雜草幼苗。
1.3 樣品采集與數據測定
待苜蓿生長到65 d時收獲,此時苜蓿生長速度明顯減慢。收獲時將花盆打破,輕輕抖掉根系上面的土壤,然后將每株的根系分開,并洗凈。首先對各苜蓿個體形態指標進行測定,利用游標卡尺測量主葉葉寬及節間粗,利用鋼尺對植物的株高、側枝總長、節間長、葉長進行測定。葉長、葉寬、節間長、節間粗測量每一主葉及每一節間后取平均值,側枝總長測量每一側枝長后取總和。待所有形態指標測量結束后,對每個單株進行根、莖、葉的分離,并置于60 ℃溫度烘箱烘干,然后稱重。
1.4 數據處理
各個性狀的可塑性指數PI參考Valladares等[23]的方法計算求得,為各處理間的最大值、最小值之差與最大值的比值,表示為PI=[max(i)-min(i) ] / max(i)。運用偏最小二乘回歸法建立個體地上生物量與各形態指標的回歸方程,并根據回歸方程的變量投影重要性指標(VIP),經計算可得各種影響要素的貢獻率(ω)[24]。計算方法:
ωXj=VIPj/∑VIPj
式中:q為自變量的個數,r(Y,th)為2各變量的協方差,Whj是軸Wh的第j個分量,ωX j為影響要素Xj的權重。
利用Excel 2010進行數據整理及簡單的數據分析,利用SPSS 19.0進行方差分析和相關分析,利用Sigmaplot 12.0作圖。方差分析法分析苜蓿各性狀在不同處理間的差異,Duncun方法進行多重比較;相關分析采用Pearson相關分析方法。
2 結果與分析
2.1 個體形態
苜蓿個體株高、節間數、節間粗、葉長均表現為P0處理顯著小于P1,P2,P3處理(P<0.05),而P1、P2、P3處理之間差異性不顯著(P>0.05);側枝總長表現為P3處理顯著大于P0,P1,P2處理,同時P0處理顯著小于P2處理,P0與P1處理間及P1與P2處理間差異不顯著;葉寬在P0處理下最小,但是P0與P3處理間差異不顯著,P0處理顯著小于P1,P2處理;P3處理具有最大的葉長與葉寬比值(葉長/葉寬),顯著大于P0,P1,P2處理,P0處理顯著小于P1,P2處理,P1與P2處理間無顯著差異;苜蓿個體節間長、節間長/節間粗在各個處理間均無顯著性差異(圖1)。

圖1 苜蓿個體形態對土壤磷元素供給能力的響應Fig.1 Responses of alfalfa individual traits to soil phosphorus supply capacity注:P0、P1、P2、P3分別代表P2O5 0、30、90、180 mg/kg磷肥添加量;不同小寫字母代表苜蓿個體形態性狀在處理間存在顯著差異(P<0.05);PH:株高;BL:側枝總長;SN:節間數;SL:節間長;SD:節間粗;LL:葉長;LW:葉寬;SLD:節間長/節間粗;LL/LW:葉長/葉寬。以下圖表符號含義與本圖相同
2.2 個體生物量及其分配
苜蓿個體各構件鮮重,包括葉鮮重、莖鮮重、根鮮重、地上鮮重及總鮮重均隨著土壤磷素供給能力的增強而呈增加趨勢(圖2A);在最小磷素供給水平P0處理下最小,顯著低于其他處理(P<0.05);在最大磷素供給水平P3處理下達到最大值,但P3處理與P1,P2處理差異未達到顯著水平(圖2A)。苜蓿個體各構件干重,包括葉干重、莖干重、根干重、地上干重及總干重對土壤磷素供給能力的響應趨勢與鮮重基本相同,在P0處理下最小,且顯著低于其他處理(圖2B);在P3處理下達到最大值,顯著高于P0,P1處理,但與P2處理差異不顯著(圖2B)。
苜蓿個體生物量分配對土壤供磷能力的響應不敏感,根冠比、根莖比、根葉比及莖葉比在處理間均無顯著差異(P>0.05)(圖3)。相關分析表明苜蓿個體生物量分配受莖干重、根干重調控,莖干重與根冠比、根莖比呈顯著相關(P<0.05),與莖葉比呈極顯著相關(P<0.01);根干重與根冠比、根莖比、根葉比呈顯著相關;葉干重、地上干重及總干重與各分配指標無顯著相關(表1)。
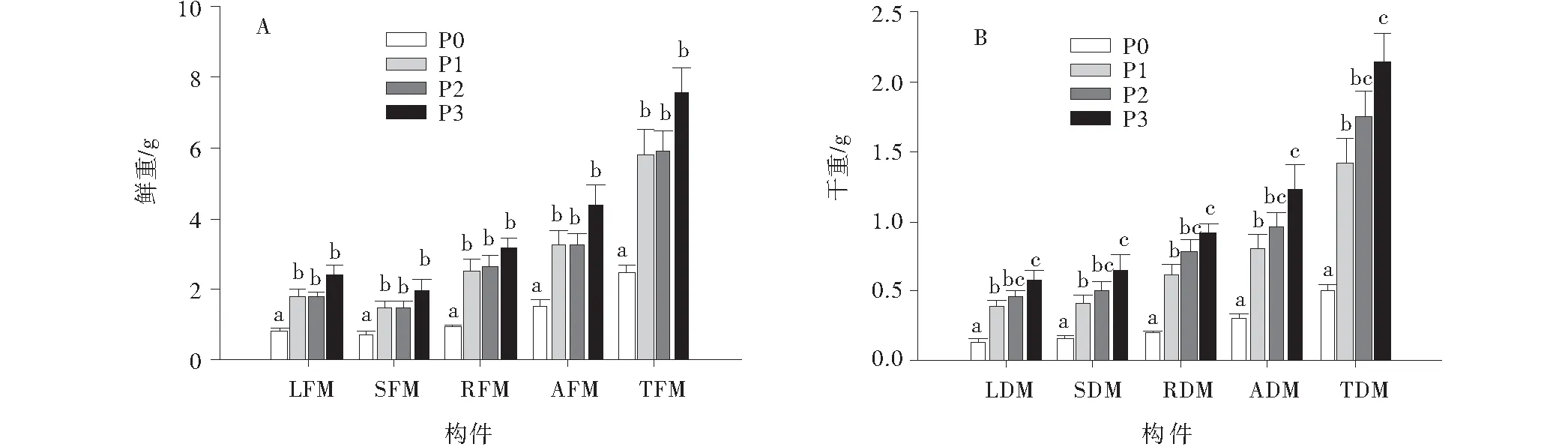
圖2 苜蓿各器官鮮重與干重對土壤磷元素供給能力的響應Fig.2 Responses of fresh weight (A) and dry weight (B) of alfalfa organs to soil phosphorus supply capacity注:LFM:葉鮮重;SFM:莖鮮重;RFM:根鮮重;AFM:地上鮮重;TFM:總鮮重;LDM:葉干重;SDM:莖干重;RDM:根干重;ADM:地上干重;TDM:總干重

圖3 苜蓿個體生物量分配對土壤磷元素供給能力的響應Fig.3 Responses of alfalfa individual biomass allocation to soil phosphorus supply capacity注:RAR:根冠比;RSR:根莖比;RLR:根葉比;SLR:莖葉比

表1 苜蓿個體各構件生物量與構件生物量分配的相關分析
2.3 個體含水量
苜蓿個體各構件含水量對土壤供磷能力響應敏感,均隨著土壤磷素供給能力的改善而呈降低趨勢;葉含水量、莖含水量、地上含水量、總含水量均表現為P0處理顯著高于其他3個磷肥添加處理(P<0.05),P1處理顯著高于P2、P3處理,P2、P3處理間差異不顯著(P>0.05);根含水量表現為P0、P1處理顯著高于P2、P3處理,P0、P1處理間及P2、P3處理間無顯著性差異(P<0.05)(圖4)。相關分析表明苜蓿含水量隨著個體株高、干重的增大而降低,苜蓿個體各構件含水量與其株高、葉干重、莖干重、根干重、地上干重及總干重均呈極顯著相關關系(P<0.01)(表2)。

圖4 苜蓿個體含水量對土壤磷元素供給能力的響應Fig.4 Responses of alfalfa individual water content to supply soil phosphorus capacity注:LMC:葉片含水量;SMC:莖含水量;RMC:根系含水量;AMC:地上含水量;TMC:總含水量.
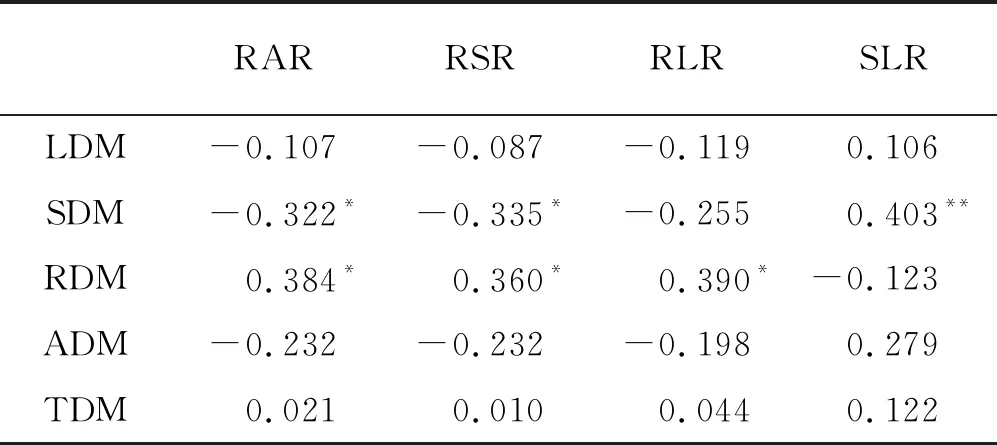
表2 苜蓿個體構件含水量與個體大小的相關分析
2.4 個體地上干重的構成因素
偏最小二乘法分析表明苜蓿個體各形態指標(側枝總長、株高、節間粗、葉長、節間數、節間長及葉寬)對其地上干物質積累量的變量投影重要性(VIP)分別為1.22,1.10,1.04,0.99,0.99,0.79,0.78(圖5A),貢獻率分別為17.66%,15.92%,15.05%,14.33%,14.33%,11.43%,11.29%(圖5B);結合本文2.1中苜蓿個體形態對磷處理的響應,土壤供磷能力主要通過苜蓿側枝總長、株高、節間粗、葉長及節間數來影響其地上干重,其中側枝總長及株高的貢獻相對較大。

圖5 個體形態性狀對地上干重的投影重要性(A)和貢獻率(B)Fig.5 The importance in projection values (A) and contribution rate (B) of morphological traits
3 討論
形態性狀既可以反映苜蓿單株生物量的大小,也可以通過光攔截影響其光合能力[14-15]。形態性狀的差異是不同品種或不同環境下苜蓿單株生物量差異的主要誘導途徑之一,例如,側枝數量及葉長的不同導致了品種間苜蓿單株生物量的差異[16]。本研究表明土壤磷環境的改善可以促進苜蓿單株生長,顯著增加苜蓿個體生物量,提高株高、側枝總長、節間數、節間粗、葉長(寬)。進一步分析表明側枝總長、株高對苜蓿單株生物量構成具有較大的貢獻,因此土壤磷環境的改善主要通過增加側枝總長和株高提高苜蓿單株生物量。
生物量分配的改變是植物響應環境變化的適應策略之一,在土壤養分或水分限制下,植物會加大根系分配以獲取較大的根系吸收面積[25-26]。研究表明苜蓿通過增加根系分配適應水分限制[27-28]。但是,也有研究發現苜蓿生物量分配并不隨環境因子的變化(氮、磷、水分、二氧化碳等)而改變[19,29-30]。本研究表明苜蓿生物量分配在各處理間無顯著差異,磷素在苜蓿生物量分配過程中無調控作用。因此,根系生物量分配的增加不是苜蓿適應低磷環境的策略之一,其可能通過其他策略適應低磷環境,如根際酸化、改變根系形態等[31-32]。眾多研究中所出現的差異性結果表明植物(苜蓿)生物量分配的調控是一個非常復雜的過程,其不僅與植物類型有關,還受到植物生長階段、環境因子脅迫強度及多因子間交互作用等因素的影響[13,33-37]。本研究表明苜蓿生物量分配與生物量多少有關,主要受根、莖干重的調控。為了充分厘清苜蓿生物量分配對環境因子的響應機制,不同苜蓿品種、不同生長階段的多因子交互試驗還有待進一步開展。
本研究表明,隨著土壤供磷能力的改善,苜蓿植株含水量顯著下降,且同干重呈顯著負相關關系,這與李星月等[38]、劉雪云等[39]、李玉娥等[40]、劉芳等[41]針對植物含水量的研究結果一致。一般來說,組織含水量越大,光合速率越快,干物質積累越多,但這與本研究結果中土壤磷素供給能力的改善能夠提高苜蓿生物量不一致。植物組織含水量代表了其幼嫩程度,與植物發育階段(物候期)有關,植物發育越成熟,含水量越低[42-44]。因此,本研究結果中苜蓿植株含水量在磷素營養改善條件下顯著降低可能與磷素營養改變了苜蓿植株的物候期有關。磷對植物物候期的影響與氮、水分等其他非生物因素相反,在磷缺乏時,植物趨向于推遲發育,延遲其物候期[45-47];而在其他非生物環境因子脅迫下,植物趨向于提前其物候期,加速完成生命周期[48]。關于苜蓿物候期對土壤供磷能力的響應在國內外鮮見報道,杜文華[49]研究表明施肥對紫花苜蓿的物候期無影響,但是總體而言,苜蓿作為多年生植物,其物候期對磷環境的響應及響應機理缺乏足夠的研究。本研究認為苜蓿物候期對土壤磷素供給能力的響應與擬南芥、玉米等一年生植物具有相同的響應趨勢,即在磷缺乏時,苜蓿趨向于推遲其發育,延遲物候期[48]。
4 結論
土壤磷素供給能力的改善可以通過改變苜蓿側枝總長、株高、節間粗、節間數及葉長提高苜蓿生物量,其中側枝總長和株高是其最主要途徑;苜蓿單株生物量分配對土壤供磷能力響應不敏感,其受苜蓿生物量的調控,相對于葉干重來說,根、莖干重是苜蓿單株生物量分配的調控因子;苜蓿單株含水量隨土壤供磷能力的增強而顯著下降,這可能與磷缺乏延遲苜蓿物候期有關。
