復合重金屬脅迫下5種植物微觀結構的變化
2021-05-27 03:37:14張云陳金發李海濤高容孫琪旗
草原與草坪 2021年1期
張云,陳金發,李海濤,高容,孫琪旗
(1.西昌學院 資源與環境學院,四川 西昌 615013;2.涼山彝族自治州林業和草原局,四川 西昌 615000 )
重金屬污染的土壤的治理與修復是生態學與環境科學近年來研究的熱點之一,其中植物修復是目前研究最多的一種有潛力、低成本的綠色土壤污染修復技術。植物細胞的微觀變化間接反映了植物的耐污力,而較強的耐活力是修復備選植物的重要條件之一[1]。長期生長在重金屬污染土壤中的植物,其體內各種生物結構會發生變化,而內部結構的變化特點最能體現出植物對環境的適應能力或環境因子對植物造成的影響[2]。近年來,國內外對異質環境下植物解剖結構變化方面的研究報道較多。崔國文等[3]發現在寒冷環境下,植物的葉脈突起度和氣孔密度會增大。孫琳旎等[4]發現長期處在干旱條件中的植物莖會為了適應環境而產生相應的變化,它們有較厚的皮層組織和排列緊密的細胞結構;易詩明等[5]在對蓖麻抵擋鉛鋅脅迫時的研究發現,蓖麻根系外皮層也會有加厚并出現木栓積累的現象。目前對生長在重金屬污染土壤中的植物莖葉解剖特征變化的報道較少,莖是植物輸送養分和水分的器官,葉片是與環境接觸面積最大的器官,兩者對外界的環境變化較為敏感,它們的解剖結構與植物對異質環境的適應性之間存在緊密關系[6]。
本試驗以某冶煉廠銅礦渣堆放地生長的5種常見植物為研究對象,以自然背景土壤中相應的5種植物做對照,對它們的莖葉進行顯微觀察,探討重金屬對植物解剖結構的傷害特點以及植物對重金屬的適應機理,為選育耐重金屬污染植物提供植物解剖學方面的參考。
1 材料和方法
1.1 試驗材料
試驗植物(experimental group,EG)與對照植物(control check,CK)分別采自于某冶煉廠銅礦渣堆放地和西昌學院周邊,時間為2018年9月。采集時選擇生長狀況良好的蓖麻、刺莧、鬼針草、空心蓮子草和紫莖澤蘭,植物的基本情況見表1,土壤的重金屬含量及理化性質見表2。
1.2 試驗方法
1.2.1 樣品的處理 將兩地采集的植物保濕帶回實驗室,先用自來水將表面污垢洗干凈,再用去離子水漂洗3遍,做好標簽放入4℃冰箱冷藏備用。

表1 試驗所用材料的基本情況

表2 基質的重金屬含量及理化性質
1.2.2 葉片下表皮氣孔結構的觀察 采用徒手切片法,先在潔凈的載玻片上加幾滴蒸餾水,然后摘取植株頂部第三片成熟葉,用鑷子輕微撕下葉片的下表皮放在載玻片的水滴上,等表皮完全展開后用蓋玻片輕輕蓋上,盡量不要有氣泡[7]。將制好的臨時裝片放在光學顯微鏡CX31(40×)下觀察并拍照記錄結果,然后用ToupView軟件測量氣孔的長度和寬度,計算長寬比。每種植物至少做3 次重復,結果以“平均值±標準差”的形式表示。
1.2.3 主葉脈橫切面的觀察 先在兩片刀片間沾幾滴清水,使兩片刀片貼緊對齊。右手捏緊刀片,從葉片主脈一端緩慢地切向另一端,注意刀片不要來回切割,把切下的切片放在裝有去離子水的干凈養皿中。用鑷子選取最薄的切片按1.2.2的方法在光學顯微鏡(10×)下觀察其結構,測量及統計的指標有葉脈厚度、葉片厚度和葉脈突起度。葉脈突起度=葉脈厚度/葉片厚度。
1.2.4 莖解剖結構的觀察 用1.2.3的方法切取植物第3~6位莖節制作成裝片在光學顯微鏡(10×)下觀察其結構,并測量統計莖表皮細胞大小和皮層厚度。
1.2.5 莖葉重金屬含量的測定 分別取1.2.1中保存的5種植物的莖葉,先在105℃的烘箱中殺青5 min,然后在70℃下烘至質量不變,用粉碎機粉碎并過80目篩。稱取0.2 g左右粉碎樣品放到消解罐中,精確記錄其質量,加入5 mL硝酸,用微波消解儀(上海屹堯,WX-4000)消解。等消解儀上顯示的溫度降到80℃以下后,將消解罐取出并放在通風櫥內。再加入2 mL H2O2至濃煙冒盡,然后將其移到50 mL容量瓶并用去離子水定容。定容后的樣品采用火焰原子吸收分光光度計(普析,TAS990)測定相應部位中銅、鉛和鋅含量。每個部位做3個平行樣,所有數據以“平均值±標準差”表示。
1.3 數據分析
試驗數據用excel進行整理,采用SPSS 23.0軟件進行數據的統計分析。
2 結果與分析
2.1 植物莖葉重金屬含量
對照植物的重金屬含量均在正常含量[8](Cu:5~30;Zn:10~100;Pb≤10;單位mg/kg)范圍內,滿足對照試驗要求。礦渣地的5種植物對于3種重金屬的總富集量除空心蓮子草外,均高于自然地的對應總富集量(圖1,圖2)。說明空心蓮子草向莖葉轉移重金屬的能力較差,具有重金屬的規避特征[9]。
2.2 葉解剖結構特征比較
2.2.1 氣孔結構 礦渣地植物與對照植物的葉表皮細胞形狀無較大差異,蓖麻、刺莧、空心蓮子草多為多邊形,鬼針草、紫莖澤蘭多為花瓣形(圖3)。

圖1 自然地植物莖葉重金屬含量Fig.1 Heavy metals content in stems and leaves of natural plants

圖2 礦渣地植物莖葉重金屬含量Fig.2 Heavy metals content in stems and leaves of slag field plants

圖3 植物葉下表皮氣孔結構Fig.3 Stomatal structure of lower epidermis of plant leaves注:圖片中所有符號表示含義見表1,下同
5種礦渣地植物的氣孔開放度都相對較小。氣孔寬度在兩種生境中沒有顯著差異。氣孔長寬比除刺莧外,其他4種植物均存在顯著差異。除刺莧外的4種礦渣地植物氣孔長度均顯著大于自然地植物,其中鬼針草氣孔長度的差異極顯著(表3)。
2.2.2 主葉脈結構 2種生境下5種植物的主葉脈解剖結構沒有差異(圖4)。表皮位于葉脈的最外層,表皮以內是大量的基本組織,基本組織的細胞較大。維管束呈半環形,木質部靠上方,韌皮部靠下方。
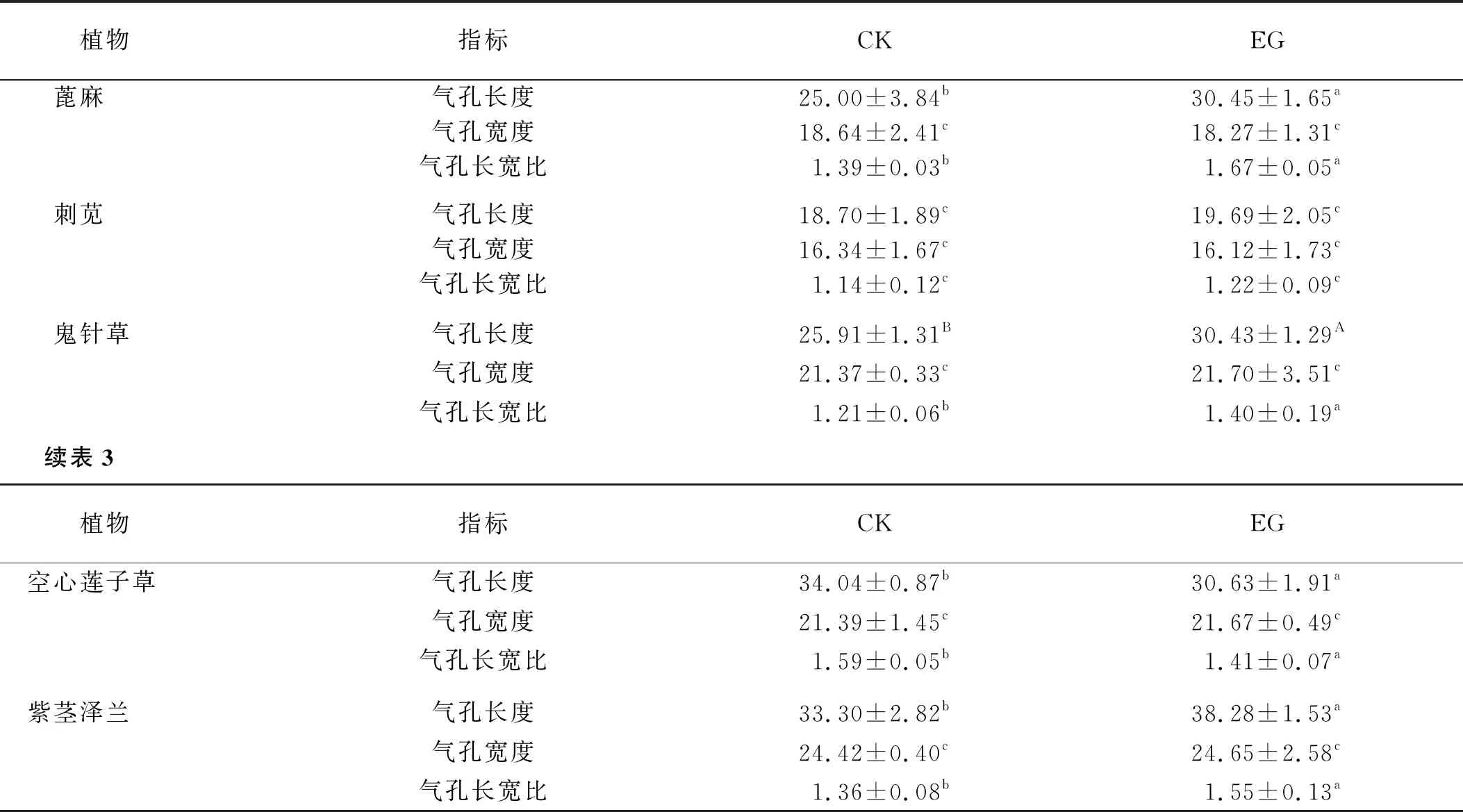
表3 兩種生境中植物葉片氣孔特征比較
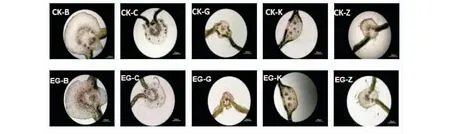
圖4 植物主葉脈橫切結構Fig.4 Transverse structure of main leaf vein of plant
與對照植物相比,礦渣地植物的葉脈及葉片厚度無顯著性差異。鬼針草主葉脈的突起度顯著大于自然地植物,而蓖麻、刺莧、空心蓮子草和紫莖澤蘭葉脈突起度在2種生境中的差異不顯著(表4)。

表4 植物主葉脈橫切結構特征
2.3 莖解剖結構特征比較
2.3.1 表皮細胞 受重金屬影響,礦渣地植物莖表皮的細胞呈長方形或不規則,大小不一,含葉綠體少(圖5)。除蓖麻外,其他4種植物莖表皮細胞寬度在2種生境中無顯著差異;除鬼針草和紫莖澤蘭外,另外3種植物莖表皮細胞長度均顯著小于對照。2種生境下鬼針草與紫莖澤蘭的皮層厚度無顯著性差異,其余3種植物的皮層厚度均在0.01水平上差異顯著(表5)。
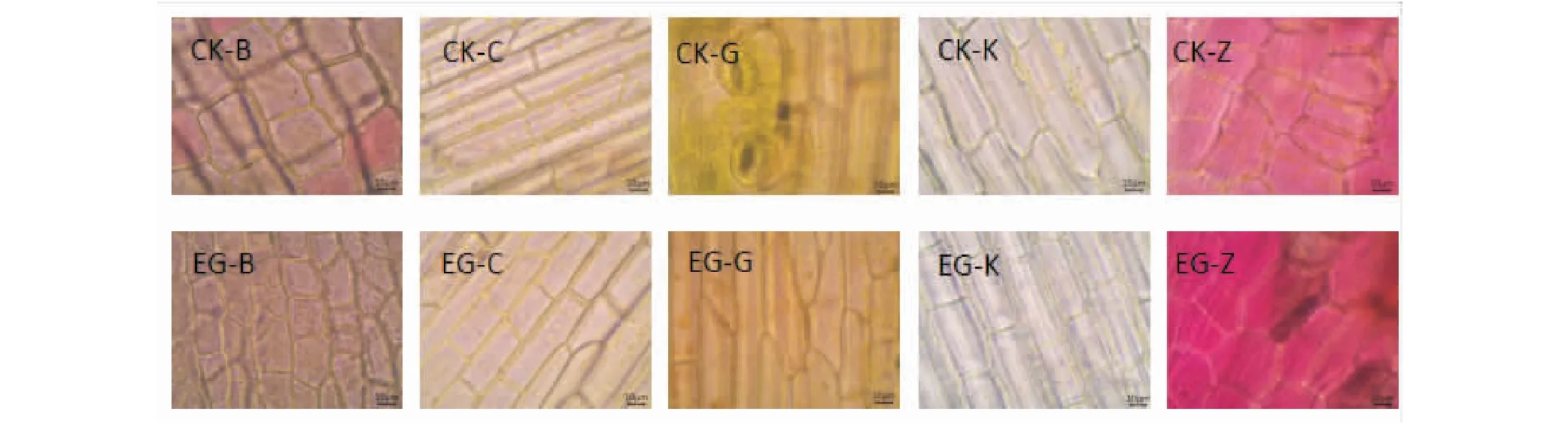
圖5 植物莖表皮細胞解剖結構Fig.5 Anatomical structure of plant stem epidermal cells

表5 植物莖解剖結構
2.3.2 莖橫切結構 5種礦渣地植物莖的橫切面結構相似,都能明顯的分辨出表皮、皮層和維管柱(圖6)。礦渣地植物莖的維管束與對照組差別大,發育受到了影響,皮層較厚(表5)。
2.4 植物莖葉中重金屬含量與解剖結構相關性分析
植物莖中銅的含量與葉片厚度呈極強正相關,葉中的銅與氣孔寬度、表皮細胞寬呈極強負相關。植物莖中鋅的含量與氣孔寬度呈極強正相關,葉中的鋅與氣孔長度、氣孔長寬比、表皮細胞長呈極強正相關。植物莖中的鉛與氣孔長度呈極強負相關(表6)。
與其他4種植物蓖麻、刺莧、空心蓮子草、鬼針草相比,礦渣地生境對紫莖澤蘭的影響相對較小,葉脈突起度、皮層厚度與自然地差異不顯著。
3 討論
植物通過調整內部結構來適應逆境,這種調節過程可以反映植物對環境變化的適應能力[10]。莖有為植物體輸送養料的作用,其組織構造對外界環境的反應較為敏感,其變化反映了植物莖對環境條件的適應以及環境對莖造成的影響,也是植物整體對環境條件的適應的結果[11]。本實驗中,植物莖對重金屬脅迫的反應主要表現為表皮細胞變小、皮層加厚。蓖麻、刺莧、空心蓮子草的表皮細胞長度和皮層厚度在自然地和礦渣生境中均存在極顯著差異。植物表面細胞變小、排列更緊密可能是由于重金屬導致其生長變慢[12]。

圖6 植物莖橫切結構Fig.6 Transverse structure of plant stem

表6 5種植物莖葉中重金屬含量與解剖結構相關性
氣孔保衛細胞是一種高度分化的細胞,它有復雜的信號轉導機制,能夠非常靈敏地感受各種生物和非生物刺激,并轉化成胞內信號來控制氣孔的大小,從而調節植物的生理狀態[13-14]。重金屬脅迫下,5種植物葉下表皮氣孔長寬比增大,氣孔開放程度減小,這是因為植物體內信號在進行傳導的過程中受到了重金屬離子的干擾或因重金屬離子導致保衛細胞活性降低而使氣孔形態和氣孔運動發生了異常。植物體內外氣體交換和進行蒸騰作用都要依靠氣孔,氣孔開放度的減小不利于氣體交換,這種變化必定會影響植物呼吸作用和光合作用等重要生理過程,從而使植物的生長發育也受到影響[15-16]。而氣孔長度增大,說明葉片在減少水分被蒸騰的同時,盡可能地維持較大的氣體交換速率,以減少重金屬對植物的迫害[17]。葉脈突起度反映了葉片與葉脈所代表的同化組織和輸導組織的相互協調、相互制約的關系,并且能夠靈敏地對外界環境變化作出響應[18]。從葉脈解剖結構來看,礦渣地植物與自然地植物的葉脈突起度差異并不明顯,重金屬并沒有對植物葉脈造成重大損傷。
植物莖葉中的鋅與氣孔寬度、氣孔長度、氣孔長寬比呈極強正相關的研究結論與SAIER[19]的研究一致,顯示表皮細胞的液泡化促進了Zn2+的積累。較高濃度的銅離子會降低葉面氣孔開度[20],氣孔寬度、表皮細胞寬與植物葉面中的銅含量呈極強負相關也說明了這一點。植物莖中的鉛與葉片氣孔長度呈極強負相關,可能是因為Pb 在向上部轉運過程中除部分被沿途輸導組織固定外,其余部分會隨著蒸騰流最終在葉片表層的氣孔和水孔附近過量累積,從而造成了對氣孔的影響[21]。
4 結論
礦渣堆放地的5種植物為了適應環境脅迫,莖葉解剖結構與對照組相比均發生了相應的變化。與自然地生境中的植物相比,礦渣地5種植物以較為細長的氣孔器、較厚的莖皮層組織等特征來適應重金屬脅迫的環境。從2種生境的莖葉解剖結構差異性來看,礦渣地生境對紫莖澤蘭的生長影響較小。
猜你喜歡
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
