匍匐翦股穎HSP26.7基因啟動子的表達分析
2021-05-27 03:37:18魏佳興朱俊飛董康挺王亭亭秦玉嬌郭琦魏雨佳李軍霖邊秀舉孫鑫博
草原與草坪 2021年1期
關鍵詞:植物
魏佳興,朱俊飛,董康挺,王亭亭,秦玉嬌,郭琦,魏雨佳,李軍霖,邊秀舉,孫鑫博
(河北農業大學 農學院/河北省作物生長調控實驗室,河北 保定 071000)
熱激蛋白(Heat-Shock Protein,HSP)是生物在逆境脅迫的情況下(如干旱、高溫等不良環境),體內才開始合成或合成量急劇增加的具有分子伴侶活性的調節蛋白質[1-2]。植物體內的一些功能蛋白在逆境條件下常導致高級構象被破壞,從而影響蛋白質功能,最終影響生物的生理代謝[3]。分子伴侶雖不具有催化活性,但能協助失活蛋白重新恢復活力,構建正確的高級結構,從而減少不良環境引起的蛋白變性對生物體的影響[4]。因此,熱激蛋白是植物對抗逆境的重要蛋白質。根據熱激蛋白分子量的大小,熱激蛋白可分為HSP100、HSP90、HSP70、HSP60以及sHSP 5大家族[5-6]。小熱激蛋白(sHSP)是熱激蛋白中的一個重要家族,雖然其分子量較小,但種類豐富,在植物體內普遍存在[7]。研究發現熱激蛋白均對高溫脅迫具有敏感性[1]。小熱激蛋白的表達不僅限于高溫誘導,且一個亞家族的小熱激蛋白的表達方式與效應蛋白也不盡相同。除了受外界環境誘導外,許多小熱激蛋白的表達還具有時空特異性,即在植物不同的發育階段,小熱激蛋白的表達量也表現出明顯的變化。
匍匐翦股穎(Agrostisstolonifera)是一類性狀優良的多年生冷季型草坪草,耐陰能力較強,具有一定的耐踐踏以及耐低修剪能力,常被用作高爾夫球場的果嶺草,當修剪至3~5 mm高度時可以形成高質量草坪[8-9];耐寒但不耐熱,在絕大多數北方地區可安全過冬,但在河北省一些地區常出現“夏枯”現象,極大地影響了草坪的使用功能以及維護成本[10]。在匍匐翦股穎熱激蛋白的研究中發現較為抗熱的品種往往要比不抗熱品種多出1~2類熱激蛋白[11]。南京農業大學黃炳茹課題組對熱激蛋白的研究已經有了一定的成果[12],但是目前對匍匐翦股穎中小熱激蛋白啟動表達的信息傳遞方式以及與下游蛋白互作的具體模式尚不清晰。因此還需從微觀領域進一步研究小熱激蛋白的具體構造以及功能。Luo Hong課題組在匍匐翦股穎抗非生物脅迫的研究中鑒定出3個sHSPs:AsHSP17,AsHSP26.7和AsHSP26.8。前期的研究表明,AsHSP26.7的表達受到高溫脅迫的誘導,并且分別在過表達水稻SUMOE3連接酶基因OsSIZ1和轉水稻microRNA393的轉基因匍匐翦股穎中的表達有所增加,且轉基因植株的抗熱性都有所增加[13-15]。因此,AsHSP26.7基因在匍匐翦股穎響應高溫脅迫中起著重要作用。然而,目前尚未發現對于AsHSP26.7啟動子的研究報道。
本試驗通過基因克隆得到HSP26.7的啟動子序列,并進行啟動子順式作用元件分析、轉基因擬南芥GUS組織化學染色和轉基因擬南芥脅迫后的表達分析,來闡述該基因啟動子的表達模式,從而為探討HSP26.7基因的功能提供一定的理論和分子基礎。
1 材料和方法
1.1 試驗材料
采用匍匐翦股穎Penn-A4品種,擬南芥為Columbia生態型,兩者皆由河北省作物生長調控實驗室自行保存,于人工氣候室進行養護繁殖,培養條件為:23℃,16 h光照,光照強度為6 000 lx。
實驗所用pEASY-T1 Simple Cloning Vector,PCRmix,限制性內切酶均購于Takara公司,GUS試劑采購于奧博萊公司,載體pCAMBIA1301、大腸桿菌DH5α感受態細胞和農桿菌感受態細胞GV3101均由本實驗室自行保存。
1.2 實驗方法
1.2.1 匍匐翦股穎HSP26.7啟動子的克隆 利用CTAB法分離純化得到匍匐翦股穎的DNA,從NCBI上查詢得到HSP26.7的起始密碼子之前的2 403 bp的序列作為啟動子,根據所得到的序列信息設計了其前后引物HSP26.7pro-F1和HSP26.7pro-R1,并以匍匐翦股穎DNA為模板對HSP26.7基因啟動子進行體外擴增。擴增所得產物連接T載體并測序,序列無誤后進行后續實驗。
HSP26.7pro-F1:5′- TA TGA AAG GCA TCA AAT AGC TCT-3′
HSP26.7pro-R1:5′- TG CTC AAG CGA GAA TCA CAG -3′
1.2.2 生物信息學分析 通過啟動子序列分析軟件PlantCARE數據庫將已知的HSP26.7的基因啟動子序列與一些已經被證實了功能的順式作用元件進行比對,尋找序列中可能參與環境脅迫、信號通路和生長發育相關的順式作用元件。
1.2.3 植物表達載體的構建 為了檢測HSP26.7基因啟動子是否成功被克隆以及能否在植物體內正常表達,本實驗構建了HSP26.7基因啟動子與GUS報告基因融合的表達載體pHL-pHSP26.7-GUS/35S-bar(圖1)。具體過程為:以HSP26.7pro-F1和HSP26.7pro-R1為前后引物,在前后引物的兩端分別添加BamH I酶切位點,用1.2.1的方法進行體外擴增。通過電泳檢測所得的條帶與預想大小是否一致,并將所需的目標條帶切割分離后連接pEASY-T1 Simple Cloning Vector,挑選連接方向正確的質粒,用BamH I進行單酶切,酶切產物經過分離后再連入pSB表達載體。體系如下:

2×Rapid Ligation Buffer10μLpSB1 μL回收片段9 μLT4 ligase1 μL
連接后在人工培養箱內4℃,放置12 h。用連接液轉化大腸桿菌DH5α感受態細胞,并通過菌落PCR檢測轉化是否成功,連接方向是否正確,再提取其質粒。將序列正確的重組質粒用凍融法轉入農桿菌LBA4404中。

圖1 植物表達載體pHLHSP26.7pro-GUS/35S-bar的構建Figure 1 Construction of plant expression vector pHLHSP26.7pro-GUS/35S-bar
1.2.4 擬南芥的轉化與檢測 用花序浸染法[16]轉化擬南芥野生型。T0代種子收獲后,播種于育苗盤,用草丁膦除草劑進行轉基因陽性苗的篩選。篩選出的陽性苗進行轉基因PCR檢測。以擬南芥陽性苗基因組DNA為模板,以引物HSP26.7F和HSP26.7R對HSP26.7進行體外擴增,以引物barF和barR對bar基因( X17220.1)進行體外擴增。
barF:5′-GTCTGCACCATCGTCAACCACTAC-3′barR:5′-GTCCAGCTGCCAGAAACCCAC-3′
PCR擴增體系:

10× PCR buffer2.5 μLdNTP mix (2.5mM each)0.5 μLHSP26.7F/barF (10μM)1.0 μLHSP26.7R/barF (10μM)1.0 μLDNA模板1.0 μLTaq DNA polymerase(2.5 U/mL)0.5 μLddH2O補至25μL
反應條件:

1.2.5 轉基因擬南芥植株的GUS組織化學染色 將WT擬南芥和轉化后的擬南芥移栽至苗床,基質為蛭石/營養土混合物(v/v=3∶1),在溫室內培養,光照強度3 000 lx,16 h光照,20℃。保持良好的肥水供應使其生長勢保持一致,待到開花時進行處理。熱處理:植株置于40℃培養箱中處理24 h;鹽處理:將植株根部浸泡在175 mmol/L NaCl溶液8 h;干旱處理:將植株從基質中挖出,洗凈根部,用濾紙吸去植株上的水分,將植株放置在實驗室的濾紙上2 h。處理后將植株進行GUS組織化學染色。
GUS染液的配制(10 mL):

0.1 mol/L 磷酸緩沖液(pH7.2)5 mL10% Triton X0.2 mL0.1 mol/L 鐵氰化鉀0.2 mL0.1 mol/L 亞鐵氰化鉀0.2 mL0.1 mol/L X-Gluc0.2 mLddH2O補至10 mL
將生長正常的擬南芥植株取樣迅速浸泡在上述染液中,真空浸泡30 min,37℃避光溫育12 h。棄染液,95%乙醇脫色,再將樣品放入70%乙醇中待樣品恢復原有形態后進行切片觀察。染出藍色的組織制作石蠟切片,置于顯微鏡下觀察。
2 結果與分析
2.1 匍匐翦股穎HSP26.7啟動子的克隆與序列分析
以HSP26.7pro-F1和HSP26.7pro-R1分別為前后引物,用提取的匍匐翦股穎DNA為模板進行體外擴增。電泳結果表明,在約2 500 bp處有一明顯條帶,其大小與預期結果相似(圖2)。將此膠切下來由Takara公司測序,測序結果用PlantCARE軟件進行順式元件比對分析。結果顯示HSP26.7基因啟動子不僅具有響應高溫脅迫的HSE元件和響應干旱脅迫的MBS元件等與植物抗逆性有關的順式元件,還有RY-element,Skn-1 motif等參與種子生殖生長的順式元件(表1)。
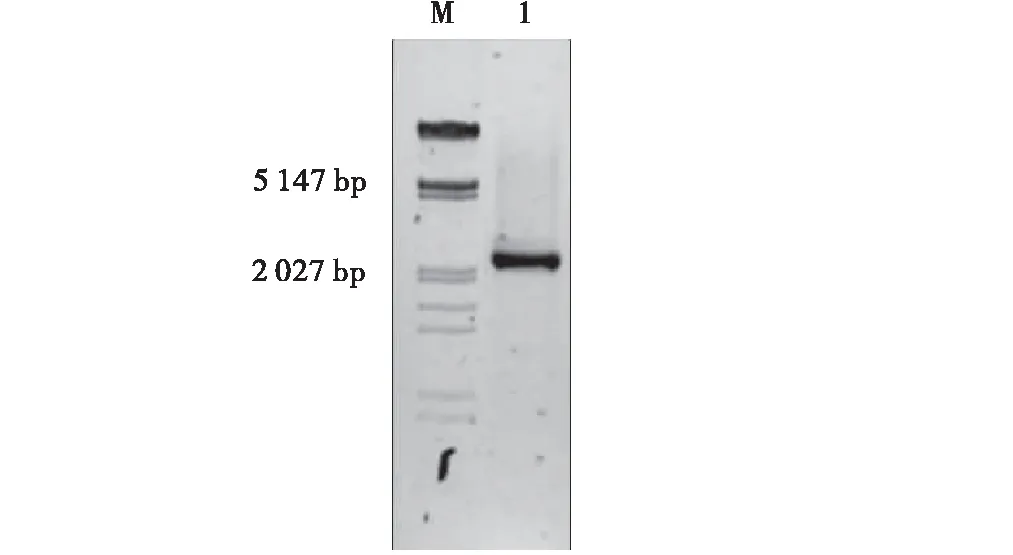
圖2 匍匐翦股穎HSP26.7基因啟動子的克隆Fig.2 Cloning of HSP26.7 gene promoter from Agrostis stolonifera L.(M: Marker;1:HSP26.7 gene promoter)注:M:Marker;1:HSP26.7基因啟動子
2.2 轉基因擬南芥的PCR檢測結果
電泳檢測結果顯示,在2 500 bp處有一明顯條帶,表明HSP26.7啟動子已經整合到擬南芥基因組中(圖3)。
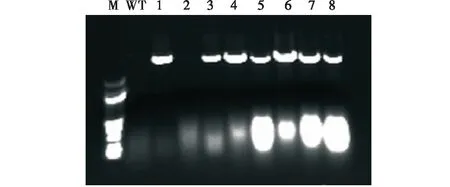
圖3 轉基因擬南芥PCR檢測結果Fig.3 PCR detection results of transgenic Arabidopsis thaliana
2.3 轉基因擬南芥GUS分析
實驗結果表明,只有在高溫誘導下的擬南芥植株才能啟動GUS基因的表達,在正常生長以及干旱和鹽脅迫的情況下并未檢測到GUS的表達(圖4)。染色結果表明,染色區域主要集中在植株頂部的生殖器官當中(花或剛剛抽穗的角果中),還有少量染色區域集中在植株葉片的基部(圖4A)。生殖器官中花藥和花絲頂部以及雌蕊的柱頭內的表達相對較多(圖4B)。花藥和柱頭染色較重的區域集中在花藥和柱頭的頂部(圖4C)。
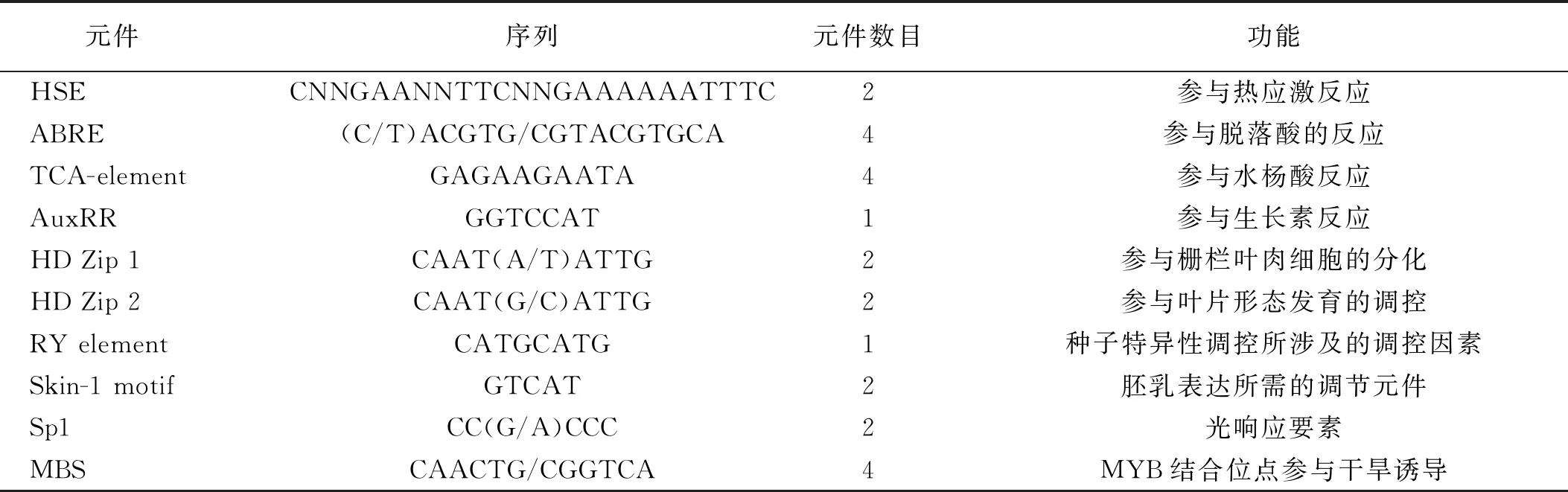
表1 匍匐翦股穎HSP26.7基因啟動子部分順式元件預測
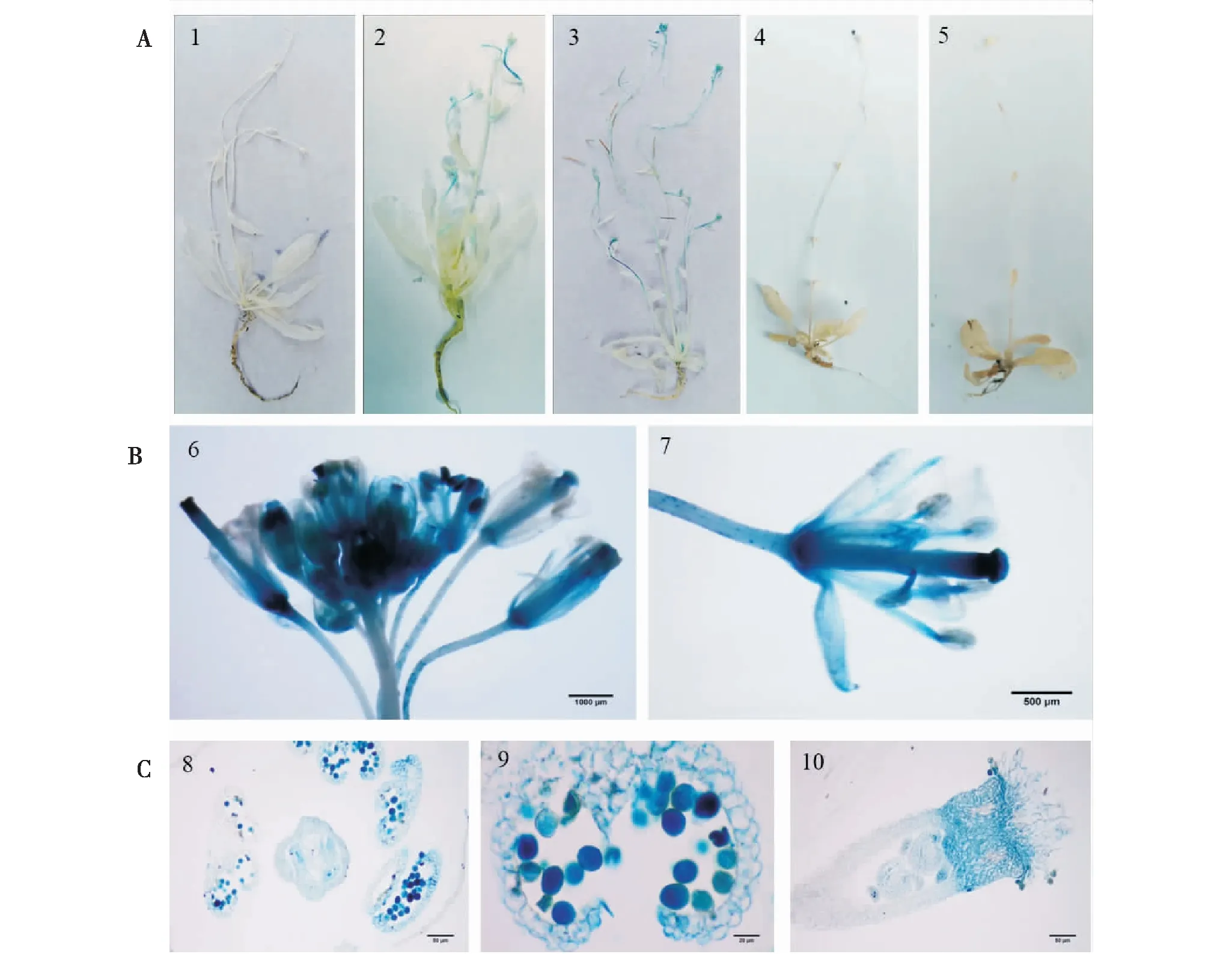
圖4 HSP26.7基因啟動子轉基因擬南芥的GUS組織染色Fig.4 Analysis of GUS activity in HSP26.7 promoter transgenic Arabidopsis注:A:轉基因擬南芥的GUS染色。1:正常生長的植株;2:40℃處理8h;3:40℃處理24 h;4:鹽處理8h;5:干旱處理2 h。B:轉基因擬南芥熱處理24 h花的GUS染色。6花序;7小花。C:轉基因擬南芥熱處理24 h花器官GUS染色。8:花藥和柱頭;9:花藥和花粉粒;10:柱頭。
3 討論
小熱激蛋白是植物對抗逆境脅迫的一種重要的蛋白因子,屬于分子伴侶的一種,幫助靶蛋白完成正確的折疊,形成正常的構象,從而使靶蛋白正常發揮功能,在植物面對高溫脅迫時起到重要作用[1-2]。小熱激蛋白相關功能的研究在草坪草中也有一定的進展[12],其中,匍匐翦股穎是一類性狀極其優良的草種,但由于其耐熱性差,導致夏季的養護管理成本較高,極大地影響了應用[10]。小熱激蛋白的過表達則為改善匍匐翦股穎在高溫下的性狀表現提供了一種可能。盡管已有資料指出小熱激蛋白會受外界環境以及某些生理生化條件誘導,但是對于小熱激蛋白感受逆境信號的機理以及改善植物抗逆性的機制尚不明確,本研究明確了小熱激蛋白的表達部位以及表達的時空特異性,為HSP26.7基因的啟動子的研究提供了理論基礎。
啟動子是位于結構基因上游、能夠與RNA聚合酶特異識別并結合的一段DNA序列,雖不具有轉錄與翻譯活性,卻是在轉錄水平上基因表達調控的中心[17]。本實驗發現HSP26.7基因的啟動子序列,設計特定的引物,將其克隆出來并轉化了擬南芥。經過序列分析發現該序列除了具有基本的啟動子特征外,還有包含很多特異的順式作用元件[18],響應高溫誘導的順式元件和與響應干旱誘導的順式元件,由此推測sHSP26.7不僅僅受高溫誘導表達,在干旱處理下也可能會表達[19]。除此之外本研究還在啟動子序列中找到了RY-element和Skin-1 motif等生殖生長的相關順式元件。由此推測sHSP26.7可能在植物不同的生長階段的表達不同,可能在生殖生長階段表達較高。
在幾種常見的逆境(高溫、干旱、鹽處理)脅迫處理下進行GUS染色,結果表明僅在熱脅迫條件下GUS基因的表達并顯色,即HSP26.7基因啟動子才能激活,說明高溫誘導能夠激活或增加熱激蛋白基因的表達,這與擬南芥HSP70-1、高粱HSP70、葉用萵苣LsHSP70-2711等基因相同[20]。染色結果還進一步表明,染色區域主要集中在花器官的頂部。這與臘梅熱激蛋白HSP1的啟動子類似,該基因啟動子在葉和根中幾乎檢測不到GUS活性,而在莖、花瓣、雄蕊和雌蕊中檢測到GUS的表達[21]。此外在大豆GmbZIP33基因轉基因擬南芥中GUS的表達也與本研究相似[16]。本研究結果表明,植物的不同部位HSP26.7基因啟動子的表達模式不同。本研究還發現GUS主要在花藥內的花粉粒和柱頭頂端的組織中表達。而HSP26.7基因啟動子也含有與生殖發育相關的順式元件(表1),這些結果表明HSP26.7基因很可能與植物的生殖生長,種子發育密切相關。HSP26.7啟動子序列中也含有響應干旱的元件,但是本研究中干旱處理后GUS表達量并沒有顯著變化,可能是由于試驗處理方式、時間或者是不同物種而出現的結果,其具體原因還需要做進一步的研究和探討。
4 結論
本實驗克隆了匍匐翦股穎中HSP26.7基因的啟動子,并研究了其生物學功能。結果表明,HSP26.7啟動子含有高溫誘導相關的順式作用元件(HSE,AT-rich element和TATA等),也含有與生殖生長相關的順式作用元件(RY-element和Skin-1 motif等)。GUS染色結果說明,HSP26.7基因啟動子只有在高溫脅迫下才能被檢測出來,并且主要表現在花粉粒、柱頭組織及葉基中。說明HSP26.7只在高溫脅迫下表達且具有組織特異性,主要在生殖器官中表達,可能與植物的生殖生長有關。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
