中國被毛孢三種菌絲形態(tài)的超顯微特征觀察
2021-05-31 07:29:08夏櫻霞蘇強(qiáng)軍UWITUGABIYEVestine陳照禾
浙江農(nóng)業(yè)學(xué)報 2021年5期
謝 放,夏櫻霞,蘇強(qiáng)軍,UWITUGABIYE Vestine,陳照禾,周 剛
(蘭州交通大學(xué) 化學(xué)與生物工程學(xué)院,甘肅 蘭州 730070)
中國被毛孢被大多數(shù)學(xué)者認(rèn)為是冬蟲夏草菌的無性型[1-3]。在實驗室條件下培養(yǎng)時經(jīng)常在典型的基內(nèi)菌絲表面或邊緣長出密集的鈕狀菌絲團(tuán)和氣生菌絲。通過觀察實驗室人工培養(yǎng)的中國被毛孢長出原基的現(xiàn)象發(fā)現(xiàn),基內(nèi)菌絲表面出現(xiàn)的菌絲團(tuán)生長一定時間后可轉(zhuǎn)變?yōu)獒橆^狀原基,但當(dāng)氣生菌絲出現(xiàn)后該生物學(xué)過程不再發(fā)生。目前,三種菌絲的超顯微特征和產(chǎn)生機(jī)制未見報道。由于以往研究的中國被毛孢菌種大多是由冬蟲夏草經(jīng)組織分離而獲得的[4],形態(tài)差異明顯的氣生菌絲究竟是雜菌或其他因素的影響無法確證。經(jīng)組織分離獲得的菌株材料,通過挑取單菌絲方法進(jìn)行純化的難度大,要排除雜菌因素,必須是單孢分離方法獲得的純化菌種才可以進(jìn)行研究。本研究采用光斑標(biāo)記法[5]分離純化得到單孢菌株,以保證菌種的可靠性,從而排除雜菌可能的干擾。
在常見的食用菌栽培中,子實體發(fā)生的菌絲變化已經(jīng)研究清楚,菌絲需經(jīng)過扭結(jié)、針頭狀鈕結(jié)、菌蕾等過程才可形成子實體[6]。而中國被毛孢形成子實體的過程還沒有明確,培養(yǎng)過程中出現(xiàn)的菌絲團(tuán)扭結(jié)是否都會發(fā)育成針頭狀原基、菌蕾等結(jié)構(gòu)仍有待研究[7]。本實驗通過對單孢純化方法獲得的中國被毛孢菌株生長發(fā)育過程中的3種形態(tài)菌絲進(jìn)行掃描電鏡觀察,掌握不同菌絲的形態(tài)特征,了解其整個生長發(fā)育過程,排除雜菌干擾的可能,為今后人工培養(yǎng)過程中促進(jìn)菌絲分化出原基進(jìn)而形成子實體提供指導(dǎo),同時對更深入研究菌絲發(fā)育機(jī)理和調(diào)控奠定基礎(chǔ)。
1 材料與方法
1.1 冬蟲夏草單孢菌株的采集與分離
2019年5月從甘肅天祝藏族自治縣帶土采集完整的新鮮冬蟲夏草,于實驗室16~18℃中培養(yǎng),使其進(jìn)一步發(fā)育成熟并彈射子囊孢子。當(dāng)子實體上的子囊殼表面出現(xiàn)白色纖維狀孢子時,將滅菌玻璃紙袋套于子實體以收集子囊孢子,每3 d更換1次玻璃紙袋。用無菌水沖洗玻璃紙袋收集子囊孢子到試管中,稀釋至每毫升大約100個,吸取200 μL的孢子懸液置于1%水瓊脂固體培養(yǎng)基中并均勻涂布后于18 ℃黑暗培養(yǎng),每2 d觀察一次孢子萌發(fā)情況。利用光斑實時標(biāo)記法[5]分離萌發(fā)的單孢子轉(zhuǎn)接到牛奶培養(yǎng)基上,置于16 ℃下黑暗培養(yǎng)。
1.2 培養(yǎng)基
本研究采用1%水瓊脂培養(yǎng)基(瓊脂10 g,加入蒸餾水定容至1000 mL,pH自然)和牛奶培養(yǎng)基(馬鈴薯200 g,牛奶200 mL,葡萄糖20 g,磷酸二氫鉀1 g,硫酸鎂0.2 g,酵母粉1 g,水解乳蛋白5 g,復(fù)合維生素B1一片,瓊脂15 g,加入蒸餾水定容至1000 mL,pH自然),配置的培養(yǎng)基經(jīng)121℃高壓滅菌后備用。
1.3 中國被毛孢三種菌絲的形態(tài)觀察
每7 d觀察1次牛奶培養(yǎng)基上的單孢菌落生長情況,參照文獻(xiàn)[8-9]從宏觀上對菌落生長速度、菌落顏色、色素分泌、菌絲質(zhì)地等方面進(jìn)行形態(tài)鑒定并拍照記錄。微觀上使用光學(xué)顯微鏡和掃描電子顯微鏡對菌絲粗細(xì)、彎曲度、膨大部分、菌絲間關(guān)系、菌絲表面特征等方面進(jìn)行觀察分析并拍照記錄。
1.4 三種菌絲形態(tài)的掃描電鏡觀察
本研究采用掃描電子顯微鏡觀察菌株TZ8-1的3種常見菌絲形態(tài)。分別挑取氣生菌絲、菌絲團(tuán)、基內(nèi)菌絲于2.5%戊二醛中4℃固定12 h;0.1 mol·L-1的磷酸緩沖液漂洗3次,每次15 min;各體積分?jǐn)?shù)梯度乙醇(30%、50%、70%、90%、95%)脫水1次后用100%乙醇脫水2次,每次20 min;用乙醇和乙酸異戊酯混合液(體積比1∶1)處理30 min;最后用乙酸異戊酯處理2 h后置于冷凍干燥機(jī)中處理15 h;干燥后的菌塊固定于樣品臺上進(jìn)行噴金處理,置于掃描電鏡下觀察、測量3種形態(tài)菌絲的形狀和大小并拍照。
2 結(jié)果與分析
2.1 三種菌絲的宏觀形態(tài)特征
本研究發(fā)現(xiàn),菌株TZ8-1在牛奶培養(yǎng)基上生長緩慢。培養(yǎng)60 d時表現(xiàn)為革質(zhì)狀的基內(nèi)菌絲(圖1-A),呈蚯蚓糞狀,質(zhì)地堅韌,多隆起褶皺,呈現(xiàn)棕黃色,且整個菌落會從中間部位隆起并蔓延,菌落背部凹陷,有少量褐色色素分泌至培養(yǎng)基。培養(yǎng)120 d時,在基內(nèi)菌絲表面形成若干致密且表面粗糙的絨狀白色菌絲團(tuán)(圖1-B)。同一條件下繼續(xù)培養(yǎng)180 d時白色疏松的氣生菌絲(圖1-C)開始增多,并逐漸延伸至整個菌落表面。
2.2 三種菌絲的光學(xué)顯微鏡觀察
由圖2可以看到,中國被毛孢三種不同形態(tài)的菌絲存在顯著差異。基內(nèi)菌絲(圖2-A)的典型特征為:菌絲隔膜不明顯,菌絲較粗,排列緊密,分支多,彎曲度高,菌絲之間多以相互纏繞的方式分布。
菌絲團(tuán)(圖2-B)的典型特征為:菌絲隔膜不明顯,隔膜間距和菌絲粗細(xì)不均勻,分支多、彎曲度高,菌絲之間交錯、重疊現(xiàn)象明顯,菌絲雜亂生長,排列無規(guī)律性。說明菌絲有扭結(jié)的趨勢,但外觀沒有呈現(xiàn)明顯的針頭狀,說明原基的分化還受其他因素的調(diào)節(jié)或刺激。
氣生菌絲(圖2-C)的典型特征為:菌絲平直、隔膜明顯,菌絲粗細(xì)均勻,排列疏松,分支少,次級菌絲多且以并列方式分布,彎曲度低,菌絲之間相互融合、交錯、重疊現(xiàn)象不明顯,生長較為整齊,具有方向性。同時在菌絲頂端或中部以出芽的方式形成大量的分生孢子(圖2-C中箭頭1、2所指部分),多數(shù)呈腎形,少數(shù)以橢圓形、長橢圓形及近球形等形態(tài)存在,形成后掉落或附著在菌絲上,說明氣生菌絲是中國被毛孢無性孢子繁殖的主要狀態(tài)。這一現(xiàn)象暗示著在培養(yǎng)后期,隨著營養(yǎng)和水分的消耗,氣生菌絲為了爭奪營養(yǎng)空間加速生長,進(jìn)而產(chǎn)生了大量的分生孢子。
綜上,3種菌絲的顯微形態(tài)差異較大。基內(nèi)菌絲粗且多分支,彎曲度高,排列緊密且相互纏繞。構(gòu)成菌絲團(tuán)的菌絲粗細(xì)不一,多分支,排列無規(guī)律。氣生菌絲粗細(xì)均勻,少分支且彎曲度低,排列疏松。
2.3 三種菌絲的掃描電鏡觀察
本研究對3種菌絲的掃描電鏡(圖3)觀察可以確定菌絲的粗細(xì)、彎曲度、膨大部分、菌絲間關(guān)系、菌絲表面特征等微觀特征。對3種形態(tài)菌絲的特征描述如表1所示。

A,基內(nèi)菌絲;B,菌絲團(tuán);C,氣生菌絲。標(biāo)尺為5 μm。A,Substrate mycelium;B,Hyphae kont;C,Aerial mycelium. Bar=5 μm.圖2 中國被毛孢TZ8-1三種菌絲的光學(xué)顯微圖Fig.2 Optical micrographs of three kind of mycelium of Ophiocordyceps sinensis TZ8-1
圖3-A顯示基內(nèi)菌絲排列緊密,圖3-B中箭頭所指為基內(nèi)菌絲的“H”型融合,與毛雄明等[10]的觀察結(jié)果一致。圖3-C中箭頭所指為部分基內(nèi)菌絲發(fā)生重疊。基內(nèi)菌絲發(fā)生菌絲的“H”型融合和重疊,且菌絲較粗、多分枝,這種生長方式增強(qiáng)了菌絲對環(huán)境的適應(yīng)能力,達(dá)到吸收更多營養(yǎng)物質(zhì)的目的,有利于菌絲網(wǎng)絡(luò)的形成[11],為中國被毛孢進(jìn)一步的生長發(fā)育奠定基礎(chǔ)。
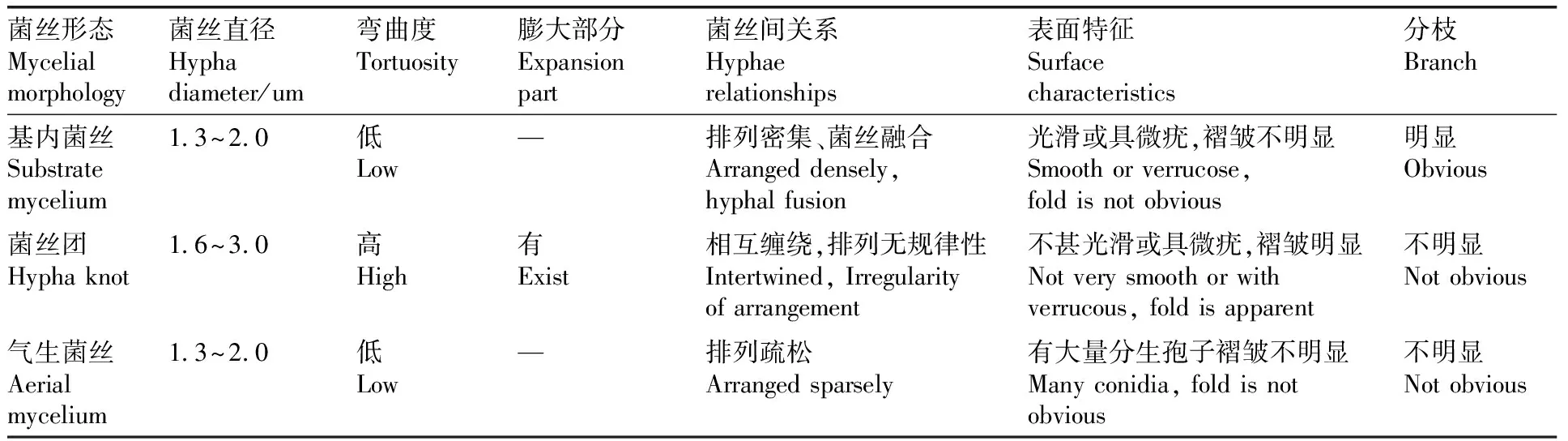
表1 中國被毛孢TZ8-1三種菌絲的掃描電鏡特征

A—C,基內(nèi)菌絲;D—F,菌絲團(tuán);G—I,氣生菌絲。標(biāo)尺:A,D,G為20 μm;B,E,H為10 μm;C,F(xiàn),I為 5 μm。A-C,Substrate mycelium;D-F,Hypha knot;G-I,Aerial mycelium. Bar: A,D,G=20 μm;B,E,H=10 μm;C,F(xiàn),I=5 μm.圖3 中國被毛孢TZ8-1三種菌絲的掃描電鏡圖Fig.3 Scanning electron microscopy of three kind of mycelium of Ophiocordyceps sinensis TZ8-1
圖3-E中箭頭所指部分為菌絲的膨大部分,此現(xiàn)象發(fā)生在基內(nèi)菌絲完成營養(yǎng)生長后出現(xiàn)的菌絲團(tuán)中,圖3-F中箭頭所指為菌絲的扭結(jié)部分,在其他食用菌相關(guān)研究中,刺芹側(cè)耳在給予光照刺激后菌絲發(fā)生扭結(jié),形成原基,進(jìn)而分化成菇蕾[12]。金針菇菌絲體受到機(jī)械損傷后通過上調(diào)表達(dá)疏水蛋白來誘導(dǎo)菌絲的扭結(jié)進(jìn)而形成子實體[13]。
圖3-I中1所指為氣生菌絲表面所附著的分生孢子,其數(shù)量多、分布廣。這表明在營養(yǎng)匱乏的后期為爭奪營養(yǎng)空間,氣生菌絲不斷延伸并產(chǎn)生大量的分生孢子加速繁殖。圖3-I中2為菌絲發(fā)生的褶皺部分,猜測其原因是樣品制備過程中脫水或干燥環(huán)節(jié)操作不當(dāng)造成的。
3 討論與結(jié)論
關(guān)于分生孢子的產(chǎn)生方式。本研究發(fā)現(xiàn)分生孢子在菌絲頂端或中部直接以出芽的方式形成,這不同于王燦[14]與何蘇琴等[15]的報道,中國被毛孢的分生孢子產(chǎn)生于已分化的分生孢子梗或具有一定形狀小梗上。但與肖巖巖等[16]的觀察結(jié)果一致,即分生孢子可通過在菌絲兩側(cè)或頂端產(chǎn)生芽生孢子的方式產(chǎn)生。與前兩者不同的原因可能是所用菌種來源于甘南藏族自治州且經(jīng)組織分離獲得,而與后者相同的原因是所用菌種由單個子囊孢子分離而來。因而,菌種的產(chǎn)地來源、分離方法和培養(yǎng)方法都是造成差異的因素。此外,取樣時間也可能對分生孢子產(chǎn)生方式的觀察有一定的影響,其具體原因有待進(jìn)一步深入研究。
菌絲團(tuán)中的菌絲膨大現(xiàn)象可能是作為一個臨時儲藏營養(yǎng)的場所[17],蓄積營養(yǎng)為進(jìn)一步發(fā)育成針頭狀原基做準(zhǔn)備。膨大部分是否為扭結(jié)出現(xiàn)針頭和菌蕾的前期征兆還未可知。因取樣時間限制了后期進(jìn)一步觀察菌蕾的形成,故菌絲膨大的原因有待深入研究。
三種菌絲的發(fā)生順序有時間差異,即與培養(yǎng)時間或營養(yǎng)脅迫有關(guān),如香菇菌絲在營養(yǎng)消耗至不適于繼續(xù)生長時菌絲發(fā)生扭結(jié)形成菌絲團(tuán),最終發(fā)育成原基[18]。
本研究發(fā)現(xiàn),菌絲團(tuán)未能扭結(jié)成為針頭狀原基,推測原因是上述培養(yǎng)基不能滿足發(fā)育成針頭狀的營養(yǎng)條件,需要其他因素的協(xié)助刺激。其內(nèi)在機(jī)理有待進(jìn)一步探討。
綜上所述,中國被毛孢在固體培養(yǎng)基上出現(xiàn)的3種菌絲形態(tài)與菌株純度無關(guān),是菌種的特性之一。三種菌絲的顯微形態(tài)差異明顯,氣生菌絲是中國被毛孢無性孢子繁殖的主要狀態(tài)。 且分生孢子的產(chǎn)生方式是芽生。
