魚類肌間骨發育分子調控機制及遺傳選育研究進展
2021-06-02 08:28:40王旭東聶春紅高澤霞
水生生物學報 2021年3期
關鍵詞:研究
王旭東 聶春紅 高澤霞 ,
(1. 華中農業大學水產學院,農業農村部淡水生物繁育重點實驗室/農業動物遺傳育種與繁殖教育部重點實驗室,武漢430070; 2. 長江經濟帶大宗水生生物產業綠色發展教育部工程研究中心,武漢 430070;3. 湖北省名優魚育種與健康養殖工程技術研究中心,武漢 430070)
肌間骨, 俗稱魚刺, 是由肌膈中的肌腱骨化而來, 僅存在低等真骨魚類中。我國是水產養殖大國,主要養殖大宗淡水魚類, 即青魚、草魚、鰱、鳙、鯉、鯽和魴[1], 都存在一定數量的肌間骨。肌間骨的存在不僅極大限制了其加工以及出口創匯, 而且也給食用這些魚類的人們帶來麻煩和一定的受傷風險。但因這些大宗淡水魚類味道鮮美、養殖技術容易掌握及歷史文化和傳統消費習慣等因素, 使得大宗淡水魚在我國水產養殖業中占據重要地位。據《2019中國漁業統計年鑒》統計, 2018年我國大宗淡水魚產量占到了全國淡水養殖魚類產量的66.45%[2]。因此, 對肌間骨的相關研究, 如肌間骨形成的分子機制的解析及培育無刺或少刺的大宗淡水魚的育種技術也逐漸成為當前研究的熱點。
長期以來, 人們對魚類肌間骨的研究主要集中在形態和數目方面[3—8]。近期, 常永杰等[9]發現刀鱭(Coilia nasus)和短頜鱭(C. brachygnathus)背部上方和腹部下方有肌骨桿(Myorhabdoi)。國內對于肌骨桿的研究報道較少, 該發現對我國魚類肌間骨類型研究做了進一步補充。Yang等[10]對12個亞科共592尾鯉科魚類肌間骨的發生進行了系統進化研究,研究揭示了鯉科魚類肌間骨的變異多樣性, 且與其系統發育密切相關, 但又受到多種環境因素的影響,對肌間骨的發生演化做了進一步補充。
隨著多學科的交叉融合, Fiedler等[11]研究了北大西洋鯡(Clupea harengus)肌間骨的微結構及相關屬性, 采用機械拉伸試驗、掃描電子顯微鏡法(Scanning electron microscopy, SEM)、寬角X射線散射法(Wide-angle X-ray scattering, WAXS)和micro-CT成像技術等方法評估了肌間骨組織礦物密度(Tissue Mineral Density, TMD)及肌間骨內部微觀結構, 進而對北大西洋鯡大體形和小體形兩個群體的肌間骨進行了比較分析。研究結果表明, 與其他類型的骨相比, 肌間骨具有十分特殊的剛度、強度和韌性。該研究為硬骨魚類骨骼生物力學和微觀結構特性提供了新的數據。
近年來, 隨著生物技術的快速的發展, 研究人員開始關注肌間骨骨化模式[12—14]、肌間骨相關的分子調控機制和少/無刺魚的培育等方面。何蘋萍等[15]、Lü等[16]、Nie等[17,18]和Wan等[19,20]研究了肌間骨骨化過程的分子機制及一些關鍵基因的調控作用。在育種方面, Tang等[21]和Xiong等[22]分別評估了鏡鯉(Cyprinus carpio var. specularis)和團頭魴(Megalobrama amblycephala)肌間骨的遺傳力, 為開發無或少肌間骨的相關品種奠定了遺傳學基礎。徐曉峰等[23]和Perazza等[24]分別發現無肌間骨突變體草魚(Ctenopharyngodon idella)和大蓋巨脂鯉(Piaractus brachypomus), 為探索肌間骨發生的分子機制提供了重要的實驗材料。因此, 本文綜合整理了有關肌間骨的研究資料, 主要從肌間骨相關的分子機制及育種前景等方面進行了全面闡述。
1 肌間骨發育相關基因調控
1.1 BMP家族基因
骨形態發生蛋白(Bone Morphogenetic Protein,BMP)是1965年由Urist首次從成人骨組織中提取的一種活性蛋白質, 除了BMP-1外, 均屬于轉化生長因子-β(Transforming growth factor, TGF-β)超基因家族[25,26]。目前, BMP家族基因已經超過30種,Zhang等[27]據dCHIP分析將已發現的人類14種骨形態發生蛋白分為3種類型: BMP2、BMP4、BMP6、BMP7和BMP9屬于成骨/成軟骨/成脂類型, BMP5、BMP11、BMP12、BMP13、BMP14和BMP15屬于肌腱發育類型, BMP3、BMP8和BMP10屬于BMP3類型[27]。BMP具有誘導成骨的作用, 被認為是誘導骨形成及骨分化最早期的信號分子。BMP蛋白主要由單體形式通過二硫鍵結合形成二聚體[28]。BMP的同二聚體和異二聚體被報道能與有成骨潛能及未分化的間充質干細胞表面受體相互作用, 從而促進間充質干細胞分化[25,26]。BMP9被認為是誘導骨髓間充質干細胞成骨的最有效因子之一[29]。
Zhang等[30]運用qRT-PCR技術初步分析了9個bmp基因在團頭魴肌間骨4個發育關鍵時期中的表達, 結果表明: 在肌間骨形成的早期階段,bmp3、bmp4、bmp5和bmp8a的表達量均上調。因此, 這些BMP家族基因可能參與刺激間充質細胞向肌間骨生長位點的遷移、聚集及成骨細胞的分化。在肌間骨快速發育時期, 只有bmp7b和bmp16表達急劇增加, 說明bmp7b和bmp16可能參與成骨細胞分化和肌間骨的發育成熟。在團頭魴肌間骨發育過程中, 表達量最高的bmp7bmRNA在肌間骨快速發育時期達到最高峰, 由此表明BMP-7b可能是肌間骨發育的強誘導因子, 用于誘導成骨細胞發育成熟。在肌間骨完全發育成熟的時期,bmp2a、bmp2b、bmp3、bmp4、bmp8a和bmp14相對于其他骨化時期的表達量顯著增加, 推測這些基因對維持肌間骨結構等方面有重要作用。Yang等[31]對羅非魚和團頭魴的bmp2a/b基因的研究表明,bmp2b主要在肌間骨、肋骨和鰭條等骨相關的組織表達, 而bmp2a主要在肌肉、脾和心臟等外周組織表達。由此說明,bmp2b可能與肌間骨的發生發育密切相關。Su和Dong[32]在轉錄和翻譯水平比較分析了bmp4基因在鯉和羅非魚的背部肌肉、腹部肌肉和尾部肌肉的表達情況, 結果表明bmp4基因在鯉背部和尾部肌肉這兩處含有肌間骨的組織中高表達, 其他4個組織也有表達, 但表達量遠遠低于在鯉的背部和尾部肌肉中的表達量。通過對有無肌間骨魚類和有無肌間骨組織的比較, 說明bmp4基因可能是調控肌間骨的重要基因。
1.2 肌腱發育相關基因
有研究認為肌間骨是由肌腱骨化而來的[33,34]。在肌腱發育過程中, Scleraxis(SCX)和Tenomodulin(TNMD)等作為肌腱發育的特異性轉錄因子受到廣泛關注。SCX是一個含有堿性螺旋-環-螺旋(basic Helix-Loop-Helix, bHLH)的轉錄因子, 是肌腱祖細胞最早的標志, 在肌腱的發育成熟中起了重要的作用[35]。在哺乳動物中, Tomoya等[36]利用scx-GFP轉基因小鼠研究了SCX在肌腱損傷后修復作用, 結果表明SCX是一個重要的加快修復損傷成熟肌腱的分子靶標。Shukunami等[37]應用TALEN介導基因編輯技術獲得scx–/–小鼠表現出肌腱發育不良的表型。同時, Shukunami等[37]也發現SCX是tnmd基因的轉錄調控因子, 在scx–/–小鼠的肌腱和韌帶中tnmd基因幾乎不表達。TNMD是一種肌腱/韌帶特異性標記物和抗血管生成分子, 屬于II型跨膜糖蛋白, 含有高度保守的c-末端富含半胱氨酸的結構域[38,39]。Yin等[39]發現tnmd–/–小鼠的肌腱干細胞基因表達譜顯著改變, 表現為細胞對I型膠原質黏附不足, 細胞外基質的收縮能力受損。
在硬骨魚類的研究中, Nie等[40]通過Crispr/Cas9基因敲除技術分別獲得了scxa–/–和scxb–/–斑馬魚突變體,scxa–/–斑馬魚與野生型斑馬魚相比背部肌間骨消失, 僅尾部含有少量肌間骨; 進一步通過轉錄組分析發現scxa–/–斑馬魚肌腱發育相關基因表達量均下調。而scxb–/–斑馬魚肌間骨表型與野生斑馬魚無異。該研究表明scxa基因可控制硬骨魚類肌間骨的數量性狀。陳宇龍等[41]研究了肌腱發育相關基因tnmd和xirp2a在團頭魴肌間骨不同發育時期的表達, 研究結果表明tnmd基因在團頭魴肌間骨快速發育時期表達量顯著高于其他時期, 而xirp2a基因在肌間骨發育各個時期沒有顯著差異。該研究指出tnmd基因可能對肌間骨的發育具有一定的調控作用。
1.3 硬化蛋白基因
硬化蛋白(Sclerostin, SOST)是一種含有213個氨基酸的糖蛋白, 在骨細胞中特異性表達[42], 抑制成骨細胞的活動, 其主要作用是抑制Wnt/β-catenin通路, 從而抑制骨的形成。在sost基因敲降的小鼠中可以觀察到高骨量的表型, 并且抗骨質流失[43]。硬化蛋白抗體能夠刺激骨形成, 這為治療骨質疏松提供了一種新的方法[44]。
硬化蛋白基因在硬骨魚類肌間骨中的研究較少。楊敏璇等[45]利用分子生物技術體外克隆鯽(Carassius auratus)sost基因全長, 并構建原核表達載體, 體外表達硬化蛋白。結果表明, 隨著時間的遞增, 硬化蛋白表達量逐漸增大, 誘導時間為4h左右其表達量最高。該研究表明, 信號肽存在對sost基因的蛋白表達存在抑制影響。房連聰[46]對淇河鯽肌間骨發育時期的研究推測, SOST可能參與淇河鯽肌間骨骨化負調控, 對骨骼發育存在抑制作用。王良炎等[47]對淇河鯽成魚不同肌間骨相鄰肌組織的研究發現,sost在肌隔組織中大量表達且在淇河鯽背部肌肉中存在差異性表達。田雪等[48]以淇河鯽仔稚魚為研究對象, 檢測sost基因在肌間骨不同骨化階段的mRNA和蛋白表達變化情況, 結果表明該基因在肌間骨完全成熟后表達量達到最高,推測sost基因與鯽的肌間骨生長有一定相關性。
1.4 其他基因
在脊椎動物中, 肌節同源盒(Muscle segment homeobox, MSX)在上皮-間充質的交互作用中發揮重要作用, 促進器官的形成[49]。Lü等[16]運用整胚原位雜交和qRT-PCR技術研究Msx-C在唇?(Hemibarbus labeo)肌間骨發生過程中的作用。Msx-C基因在唇?26—41 dph(Day post hatching)在肌膈中表達, 該基因與唇?肌間骨骨化過程(35—62 dph)一致, 表明Msx-C基因在唇?肌間骨形成過程中發揮重要作用。
TWIST是高度保守的堿性螺旋-環-螺旋(Basic helix-loop-helix, bHLH)家族中的轉錄因子, 參與中胚層衍生組織(包括骨骼)的發育[50]。脊椎動物有兩種twist基因, 即twist1和twist2[51]。研究發現,twist1和twist2對小鼠骨骼的發育具有調控作用[52]。陳潔等[53]通過qPCR研究twist1和twist2在唇?肌間骨發育的4個關鍵時期的表達情況, 結果表明twist1表達量隨肌間骨發育逐漸降低, 肌間骨快速發育和成熟時期twist1表達量與肌間骨未發育時期相比顯著降低;twist2從肌間骨未發育時期到肌間骨快速發育時期表達量并未發生顯著變化, 到肌間骨完全成熟時期其表達量才顯著降低。這表明twist1/2基因的表達與肌間骨骨化存在一定的相關性。
當前, 對于魚類骨骼發育的研究還比較缺乏,研究進展情況落后于在人類和哺乳動物的相關研究。同時, 魚類相對于人類和哺乳動物的進化地位低, 相同基因在進化過程中其功能可能會發生相應的改變。而且在活體水平的基因編輯會或多或少的受到補償效應的作用, 從而對表型觀察等產生影響。綜合來看, 魚類肌間骨發育相關基因仍有待挖掘和探索。
2 肌間骨相關組學分析
目前, 大多數肌間骨的研究主要是集中在個體發生發育的形態學分析, 對肌間骨發育分子調控方面的研究相對較少。近些年, 隨著高通量測序技術和分子技術的發展, 越來越多的研究人員開始運用組學技術和分子技術揭示肌間骨發生發育的分子機制(表 1)。
2.1 轉錄組
基于高通量測序技術, Wan等[19]首次比較分析了6月齡團頭魴肌間骨(IBs)和包含肌間骨的結締組織(CT)的miRNA分子特征, 獲得218個已知miRNA基因的序列和相應表達水平。結果表明MiR-206是成骨細胞分化的關鍵調控因子, 對成骨細胞分化起到抑制作用。通過stem-loop實時熒光定量對11個差異表達miRNA在9個組織上進行驗證, 結果表明mam-miR-221、mam-miR-222a、mam-miR-92a和mam-miR-26a在肌間骨和含肌間骨的結締組織中表達量相對較高, 說明其在肌間骨發育中發揮一定作用。
隨后, Wan等[20]比較分析了團頭魴肌間骨發育關鍵時期(S1. 肌間骨未發生; S2. 肌間骨初步發生;S3. 肌間骨快速發育; S4. 肌間骨發育完全)mRNA和miRNA的分子特征, 并對二者進行整合分析。結果表明TGF-β信號通路與MAPK信號通路、ERK信號通路互作, 調節成骨細胞分化。在團頭魴肌間骨4個發育關鍵時期中, 有22個miRNA持續上調和18個miRNA持續下調。通過整合分析發現一定數量的與肌間骨形成有關的mRNA-miRNA互作對,在細胞水平驗證了miR-133對靶基因Tgfbr1a的調控作用。
何蘋萍等[15]對禾花鯉(C. carpio var. Quanzhounensis)和建鯉(C. carpio var.Jian)肌間骨進行miRNA的高通量測序, 據差異表達結果推測, 肌間骨的發育可能是通過下調促進成骨miRNA的表達量及上調抑制成骨miRNA的表達量來抑制成骨過程, 從而維持禾花鯉肌間骨細小及柔軟的特性。Peng等[54]比較分析禾花鯉和建鯉肌間骨mRNA和microRNA轉錄組數據發現7個差異表達基因和12個差異表達miRNA參與禾花鯉骨礦化調控過程。隨著團頭魴全基因組測序的完成, Liu等[55]對3種關鍵組織(肌間骨、肌間骨外周結締組織和背部白肌)進行了基因差異表達分析, 進一步確認了所觀察到的基因表達差異確實與肌間骨的形成和功能有關。
2.2 蛋白組
Nie等[17]通過iTRAQ LC-MS/MS蛋白組學定量分析技術構建了魚類肌間骨和肋骨的第一張蛋白組圖譜。經蛋白組學分析發現, 一齡團頭魴肌間骨和肋骨之間的93個差異表達蛋白造成一齡團頭魴肌間骨和肋骨的化學組成差異, 二齡團頭魴肌間骨和肋骨間存在154個差異表達蛋白造成上述2種骨間的化學組成差異; 與破骨細胞分化相關的玻連蛋白b前體和金屬蛋白酶2在肋骨中顯著上調表達; 在團頭魴的不同年齡階段(一齡、二齡)發現13個肌間骨發育相關候選蛋白和11個肋骨發育相關候選蛋白。通過KEGG分析在235個通路中共定位了2023個蛋白, 其中在肌間骨發育相關信號通路, 如MAPK信號通路、鈣離子信號通路、TGF-β信號通路、Wnt信號通路及破骨細胞分化途徑分別鑒定出60、41、20、17和11種蛋白。通過GO注釋分析,HCM信號通路為差異表達蛋白最多的通路。在肋骨和肌間骨中共同表達45個蛋白, 僅有LAMA2表達上調, 其余44種蛋白表達下調。基于蛋白組數據,Nie等[17]對ECM、MAPK、GnRH及鈣離子通路中蛋白的表達模式進行分析。在ECM信號通路中鏈狀膠原蛋白a1(V)等18個蛋白在一齡團頭魴肋骨組織中高表達; 肌腱蛋白樣蛋白、層黏連蛋白亞基a-2和tnc蛋白在一齡團頭魴肌間骨中的表達量最高。
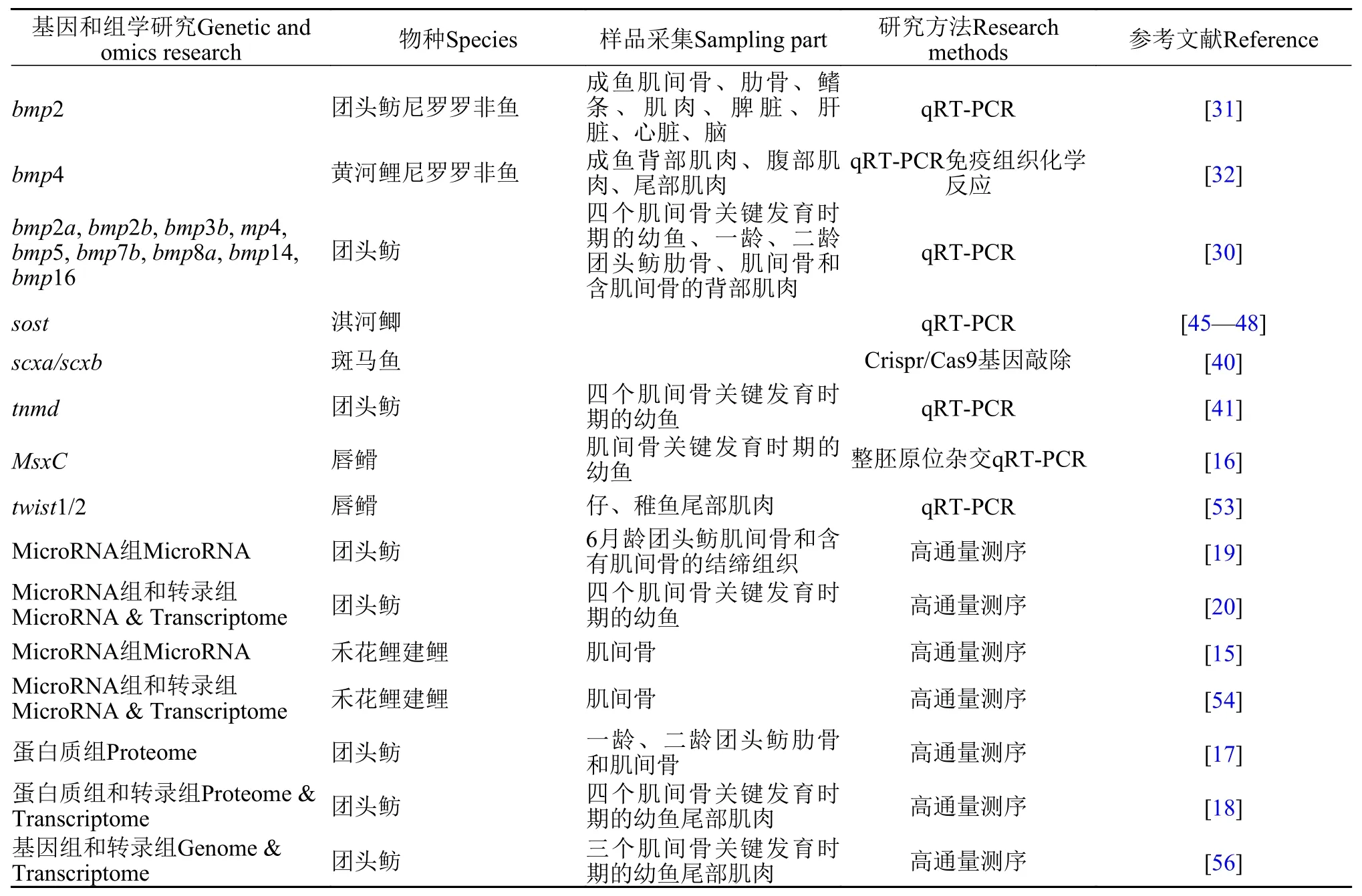
表 1 低等硬骨魚類肌間骨相關基因和組學研究Tab. 1 Summary of studies on genes or omics related to intermuscular bone in lower teleosts
2.3 多組學整合分析
在肌間骨的組織結構和成分研究方面, Nie等[17]通過HE染色切片法比較分析了一齡和二齡團頭魴肌間骨和肋骨, 切片結果表明兩種骨組織均含有三種典型的骨形成細胞, 即破骨細胞、成骨細胞和骨細胞, 但在一齡和二齡團頭魴肋骨中均發現一個富含破骨細胞的骨腔, 而肌間骨不存在類似骨腔。通過使用阿利新藍染色(用于軟骨細胞染色)和茜素紅染色(用于組織鈣鹽反應)等不同的染色方法分析,Nie等[18]對團頭魴肌間骨發育關鍵時期的個體進行整體染色觀察的研究, 結果表明肌間骨礦化從S2時期開始, S2到S4時期鈣沉積量持續增加; 但在肌間骨發育過程中無軟骨細胞形成, 表明肌間骨的骨化方式為膜內骨化。隨后, Nie等[18]綜合了組織學、轉錄組和蛋白組數據全面分析了肌間骨的骨化模式, 通過同位素標記相對和絕對定量(isobaric tags for relative and absolute quantification, iTRAQ)技術鑒定得到3368個蛋白, 功能注釋表明MARK信號通路和鈣離子信號通路是骨發生過程中的主要信號通路。經蛋白組學和定量表達分析發現, 軟骨細胞相關蛋白(COLLAGEN9A1、STAT1和TNC)在肌間骨發育過程中的表達量無顯著差異, 而與成骨細胞和骨形成相關蛋白(ENTPD5、CASQ1A、PVALB、ANXA2A和ANXA5)表達量在不同發育時期間具有顯著的差異性, 從蛋白組學的角度進一步驗證了肌間骨骨化不經歷軟骨階段的觀點。
Chen等[56]通過全基因組和轉錄組綜合分析, 獲得了MAPK信號通路、鈣離子信號通路、ECM受體互作及Notch信號通路等團頭魴肌間骨發育相關信號通路, 尤其是ECM受體互作和Notch信號通路在全基因組和轉錄組兩種分析中均得到富集。隨后在轉錄水平驗證了以上通路中的相關基因(itgal、itga6a、lamb1b、col6a3、dvl1a和oser1), 結果表明這些基因均在團頭魴肌間骨發育S3時期比S1時期顯著高表達, 與轉錄組水平相同。
近期, 基于在大蓋巨脂鯉養殖群體中發現了缺失肌間骨的個體[24], 研究團隊對突變體進行了全基因組關聯分析[57], 篩選到了13個與減少骨量、促進骨形成、抑制骨吸收和骨礦化等相關功能相關的基因, 包括actn3b、adamtsl2、atp6v0a1a、atp6v0ca、dchs1b、ebf3a、efnb1、nmu、ntn4、pde4d、plek、wisp1b和xpr1b。
基于魚類肌間骨相關組學和基因表達信息, 總結出從肌腱源干細胞-成骨細胞前體細胞-成骨細胞-肌間骨形成過程中發揮一定作用的通路及相關基因(圖 1), 為進一步探討魚類肌間骨發育的機制提供了分子基礎信息。
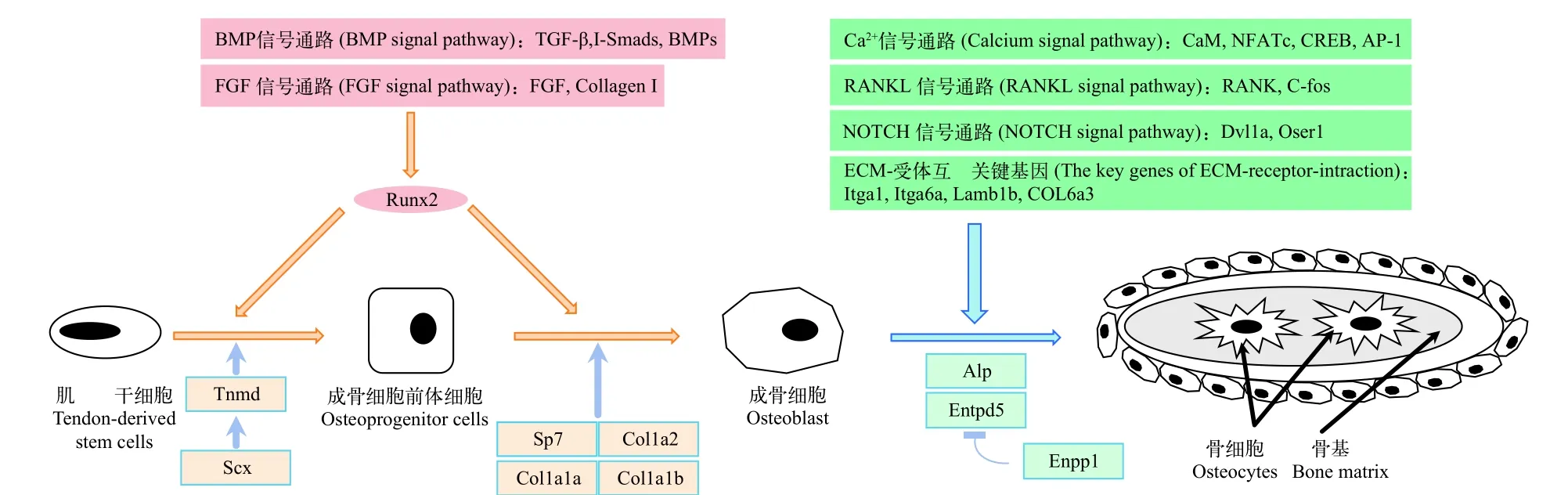
圖 1 硬骨魚類肌間骨發育相關分子機制Fig. 1 The molecular mechanism related to intermuscular bone in teleosts
3 肌間骨性狀的遺傳選育
目前, 國內大宗淡水魚類及印度、巴西等國家主要養殖的魚類品種均含有肌間骨, 對魚類經濟效益和食用安全產生了一定的影響。一些研究學者致力于運用育種手段來減少或去除魚類肌間骨, 已取得一些明顯的進展[58](表 2)。
3.1 選擇育種
自20世紀70、80年代起, 以群體選育和家系選育為主的選擇育種已廣泛應用于水生生物的遺傳改良工作中[59], 而研究人員對減少肌間骨數量方法探究開始于1967年[60]。雖然這些方法探究沒有獲得少肌間骨魚類品種[58], 但許多研究發現在鯉科魚類中肌間骨數量存在顯著的遺傳變異現象[60,61], 這種遺傳變異現象表明選擇育種可應用于減少肌間骨數量的相關研究[58]。
近些年, Cao等[62]對鏡鯉、柏氏鯉(C. pellegrini)與黑龍江鯉(C. carpio)的雜交鯉、荷包鯉的耐寒品系等3種魚的肌間骨數量和形態變異情況進行比較分析, 與其他兩種魚相比, 鏡鯉肌間骨的數量最少, 變異系數也較高。結果表明鏡鯉是一種可用于選擇育種減少肌間骨的材料, 并且認為通過選擇育種可以有效減少魚體后部肌間骨數量。Tang等[21]首次對鏡鯉肌間骨進行遺傳力和數量性狀位點(Quantitative Trait Loci, QTLs)分析, 結果表明鏡鯉肌間骨數量為中等遺傳力性狀并發現8個肌間骨QTLs和29個肌間骨相關的候選基因。該研究為鯉肌間骨分子機制的深入研究提供了資料。Xiong等[22]研究表明團頭魴整體肌間骨數量變異系數為6.98%, 而脈弓小骨數量變異系數高達13.63%,顯著高于髓弓小骨數量的變異系數(6.60%)。對83個全同胞家系團頭魴的肌間骨數量遺傳力進行評估顯示, 髓弓小骨數量為低遺傳力(0.13), 脈弓小骨數量為中等遺傳力(0.37)。Xiong等[22]認為可以通過家系選擇育種來有效減少脈弓小骨數量。在20世紀90年代后期, DNA分子標記輔助育種的出現加快了育種進程[63]。馬吉敏等[64]從200個微衛星位點中篩選出149個多態性位點, 經關聯性分析獲得8個與鏡鯉肌間骨數量顯著相關的微衛星位點。Wan等[65]利用混合分組分析(Bulked segregant analysis, BSA)方法檢測團頭魴肌間骨極少(<100)和極多(>130)兩個群體中的SNPs, 共確定了6074個SNPs且最終篩選出181個與團頭魴肌間骨數量相關的候選SNP位點。該研究首次探索了SNPs位點與肌間骨數量的關聯性, 為肌間骨數量相關的分子標記輔助育種奠定了基礎。
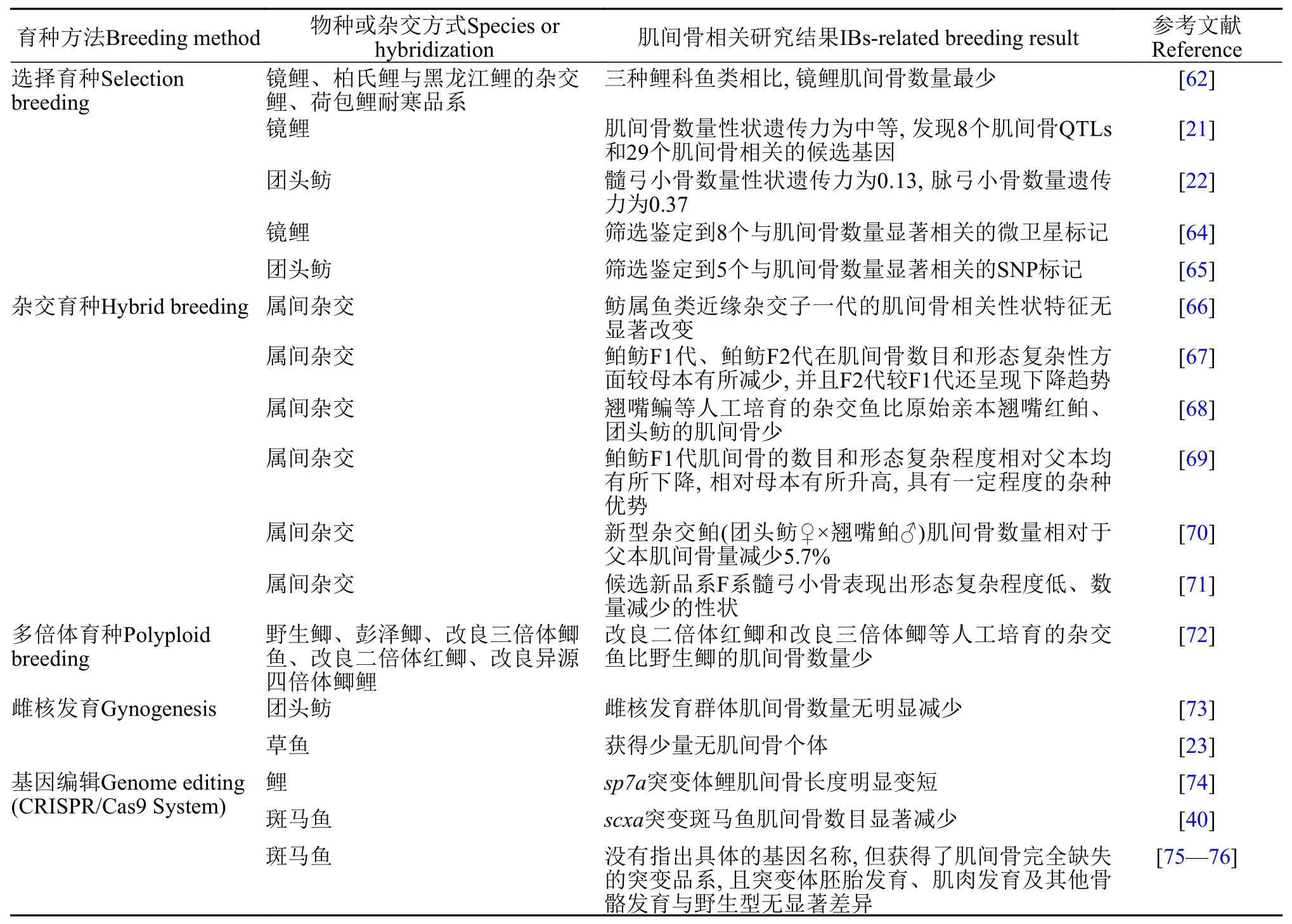
表 2 魚類肌間骨性狀相關的遺傳改良研究Tab. 2 Summary of studies on genetic improvement of fishes related to intermuscular bone traits
3.2 雜交育種
除選擇育種外, 雜交育種也是最基礎且得到廣泛應用的育種技術之一[59]。一些研究人員對不同魚類進行雜交, 對獲得的后代進行肌間骨相關分析研究。聶春紅等[64]對團頭魴與其他3種魴屬魚類[三角魴(M. skolkovii)、廣東魴(M. terminalis)和厚頜魴(M. pellegrini)]正反交及親本自交后代共10 種雜交組合子代的肌間骨數目、形態、分布和長度進行了比較分析。研究結果表明, 魴屬魚類近緣雜交子一代的肌間骨相關性狀特征無顯著改變。蔣文枰等[67]對鲌魴F1代、鲌魴F2代和母本翹嘴鲌(Culter alburnus)、父本團頭魴肌間骨的數目、形態和分布進行統計分析, 研究表明鲌魴F1代、鲌魴F2代在肌間骨數目和形態復雜性方面較母本有所減少, 并且F2代較F1代還呈現下降趨勢。鐘澤洲[68]對團頭魴、翹嘴紅鲌、鲌魴F1代及翹嘴鳊4種不同雜交魚的肌間骨的數目、形態和分布進行研究, 結果說明翹嘴鳊等人工培育的雜交魚比原始親本翹嘴紅鲌、團頭魴的肌間骨少。Guo等[69]研究廣東魴(♀)和翹嘴鲌(♂)屬間雜交F1代的肌間骨數目形態特征, 發現鲌魴F1代肌間骨的數目和形態復雜程度相對父本均有所下降, 而相對母本有所升高, 具有一定程度的雜種優勢。Wu等[70]用遠緣雜交技術培育的新型雜交鲌(團頭魴♀×翹嘴鲌♂)肌間骨數量相對于父本肌間骨量減少5.7%。李志等[71]比較分析了不同月齡異育銀鯽A+品系和候選新品系F系的肌間骨形態數量差異, 研究結果表明, 候選新品系F系髓弓小骨表現出形態復雜程度低、數量減少的性狀,相對于A+系有利于食用的優勢。
3.3 多倍體和雌核發育育種
多倍體育種和人工雌核生殖屬于細胞工程育種技術, 是在細胞或染色體組水平上進行遺傳改良的育種技術[59]。黎玲等[72]比較了5種不同倍性魚肌間骨的數目、形態和分布, 結果表明改良二倍體紅鯽和改良三倍體鯽等人工培育的雜交魚比野生鯽的肌間骨數量少, 為培育少或無肌間骨魚類提供一種育種方法。關檸楠等[73]對86尾雌核發育團頭魴群體肌間骨的數目、形態、分布和長度進行了比較分析。結果表明, 與正常的團頭魴群體肌間骨的數目相比, 該群體的肌間骨的數目并沒有明顯減少。徐曉鋒等[23]通過人工誘導雌核發育純合二倍體草魚(Ctenopharyngodon idellus)使突變基因的性狀顯現, 在5尾散鱗突變體草魚中篩選出了1尾正常發育的無肌間骨的草魚。
3.4 基因編輯育種
此外, 隨著基因編輯技術的發展, Zhong等[74]使用CRISPR/Cas9基因編輯技術敲除鯉sp7a基因, 結果表明sp7a–/–突變體存在明顯的肌間骨變短的現象, 但突變體同時存在骨缺損, 包括鰓蓋、上頜骨功能不全和背部彎曲等問題。Nie等[40]通過基因編輯技術分別獲得了scxa基因突變斑馬魚, 突變體與野生型斑馬魚相比背部無肌間骨, 僅尾部含有少量肌間骨, 總數較野生型減少約70%, 個別突變體雖然存在肋骨發育缺陷和肌膈發育缺陷, 但整體生命活動和繁殖能力與野生型沒有明顯差異。楊建等[75,76]同樣通過基因編輯技術獲得了無肌間骨的斑馬魚突變品系, 雖然作者沒有指出具體的基因, 但通過qRT-PCR對比分析5個肌肉特異性基因(mef2、myf5、myod、myog和sox6)和6個骨骼發育相關基因(bmp2a、bmp4、smad1、smad4a、runx2a和sp7)在無肌間骨突變品系和野生斑馬魚中的表達情況, 結果表明肌間骨存在與否對斑馬魚胚胎發育以及肌肉發育無顯著影響; 肌間骨缺失對斑馬魚骨骼發育無顯著影響, 只在胚胎發育時期影響骨骼相關基因表達水平的變化, 結合組織學染色結果推測,肌間骨缺失對斑馬魚骨骼發育無顯著影響。
4 研究展望
世界上對魚類肌間骨的遺傳育種研究最早始于1967年, 之后近50年, 這方面的研究進展比較慢。從圖 2可以看出, 自2010年后, 越來越多的研究人員開始圍繞魚類肌間骨分子遺傳機制和遺傳育種開展研究。研究肌間骨的分子機制對揭示魚類肌間骨的系統進化有著重要意義, 隨著分子編輯技術和單細胞測序技術的快速發展, 通過開展有肌間骨和無肌間骨魚類肌膈中肌腱組織細胞類型的確定和基因表達差異性研究, 篩選鑒定誘導有肌間骨魚類肌腱源性干細胞向成骨細胞分化的關鍵基因;通過基因過表達和基因功能缺失等研究方法明確基因的調控功能, 闡明魚類肌間骨發育的分子機制,進一步可通過CRISPR/Cas9基因編輯技術培育少或無肌間骨的魚類。
此外, 肌間骨的遺傳力為中等遺傳力且一些與肌間骨數量相關的標記基因也被篩選鑒定, 這些標記基因可以被用來指導少肌間骨魚類的選育, 尤其是鯉科魚類。隨著科學技術的快速發展和測序成本的降低, 二代/三代測序技術與全基因組關聯研究(Genome wide association studies, GWAS)技術也可以用來確定與肌間骨數量相關的標記基因, 進一步提高少肌間骨魚類選育的準確性。無肌間骨的草魚或巴西大蓋巨脂鯉可以用來與正常個體雜交,在獲得F1代基礎上進而形成F2家系, 用于QTL定位肌間骨有/無的關鍵基因, 為后續研究肌間骨的分子機制以及開展無肌間骨的魚類選育奠定基礎。中國的水產養殖已為世界做出重要貢獻, 若能培育出無“小刺”的主養大宗魚, 必將成為對世界水產的又一重大貢獻。

圖 2 硬骨魚類肌間骨相關分子機制及遺傳育種研究時間進程Fig. 2 The time course of molecular mechanism and genetic breeding related to intermuscular bone in teleosts
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
