綠原酸對蘋果采后灰霉病抗性的影響
2021-06-03 02:38:38王大將張夢宇岳正洋周會玲
食品科學 2021年9期
王大將,張夢宇,岳正洋,周會玲*
(西北農林科技大學園藝學院,陜西 楊凌 712100)
灰霉病是由一種壞死性和營養性病原性真菌灰葡萄孢(Botrytis cinereaPers.)侵染所導致的病害,是植物中最普遍和最具破壞性的病害之一,可致使500多種植物發生病害,給農業生產造成嚴重損失[1]。灰葡萄孢主要危害蘋果的嫩莖、葉和果實[2],可通過寄生在植物殘體內或以菌核的形態在土壤深處度過寒冬等惡劣環境,具有極高的生存能力[3],防治難度極大。目前防治灰霉病主要依靠使用化學殺菌劑,然而化學試劑的大量使用造成越來越多的食品安全和環境破壞等問題,并且由于灰葡萄孢的遺傳靈活性和高進化潛力,殺菌劑的效果常常會被大量具有抗藥性的田間灰葡萄孢菌株削弱[4-5]。因此,探究蘋果自身物質對灰霉病的抗性是蘋果灰霉病防治的重要手段。綠原酸(chlorogenic acid,CGA)又稱3-O-咖啡酰奎尼酸,是由咖啡酸的1位羧基和奎尼酸的3位羥基縮合成酯的天然產物[6],具有抑菌消毒、活血降壓、抗氧化、防衰老等功效[7],廣泛應用于多個行業。研究發現,植物中提取的綠原酸對根霉、大腸桿菌、青霉菌均有較強的抑制作用[8]。Jiao Wenxiao等[9]研究發現,水蜜桃果實經綠原酸浸泡處理后接種擴展青霉,其在25 ℃貯藏期間的損傷直徑和腐爛指數明顯降低,綠原酸處理對果實硬度、可溶性固形物含量、pH值、可滴定酸含量等品質指標也有顯著的改善作用。除此之外,Vali?as等[10]研究馬鈴薯塊莖中的綠原酸合成途徑時發現,綠原酸及其衍生物可以清除羥自由基和超氧陰離子自由基,減輕細胞膜脂的損傷和延緩細胞的衰老凋亡過程,提高植物的 抗病性[11]。作為蘋果中含量較高的酚類化合物,綠原酸能否作為外源誘導因素增強蘋果對灰霉病的抗性鮮有報道。本實驗以‘富士’蘋果為研究對象,以外源綠原酸浸泡處理后損傷接種灰葡萄孢,通過分析蘋果發病過程中相關酶活性和物質含量的變化,探討綠原酸在采后蘋果灰霉病抗性中的作用,以期為綠原酸在采后蘋果灰霉病的防治提供一定的理論依據。
1 材料與方法
1.1 材料、菌株與試劑
富士蘋果(Malus domestica‘FuJi’)采于西北農林科技大學白水蘋果試驗站,要求大小均勻,成熟度、著色基本一致,且無病蟲害和機械損傷。
灰葡萄孢(Botrytis cinereaPer.)分離自感染灰霉病的蘋果果實,并進行純化。將灰葡萄孢接種于PDA培養基中,并在25 ℃恒溫避光條件下培養7 d,用含體積分數0.05% Tween-80的無菌水稀釋成105個/mL的孢子懸浮液備用。
L-苯丙氨酸、苯甲基磺酰氟化物(phenylmethyl sulfonylfluoride,PMSF)、二硫蘇糖醇(dithiothreitol,DTT) 美國Sigma公司;綠原酸(純度98%) 上海源葉生物公司;聚乙烯吡咯烷酮(polyvinylpyrrolidone,PVP)、乙二胺四乙酸二鈉、β-巰基乙醇、Tris-HCl、丙三醇、TritoX-100、磷酸、無水醋酸鈉、冰醋酸、鹽酸羥胺、亮抑酶肽均為國產分析純。
1.2 儀器與設備
A11型液氮研磨儀 德國IKA公司;BSM-220.3電子 天平 上海卓精電子科技有限公司;Infinite M200pro全波長多功能酶標儀 瑞士Tecan公司;JXN-26高速冷凍離心機 美國貝克曼庫爾特公司;HH-6數顯恒溫水 浴鍋 常州國華電器有限公司。
1.3 方法
1.3.1 接種實驗
在含有綠原酸質量濃度分別為0(對照)、100、200、300 μg/mL和500 μg/mL的PDA培養基上,分別接種20 μL 105個/mL灰葡萄孢懸浮液,放入培養箱中(25 ℃、黑暗)進行培養,并觀察記錄菌落直徑,根據菌落直徑篩選出合適的抑菌質量濃度,重復3 次。
在離體實驗基礎上,最終確定300 μg/mL為綠原酸的最佳抑菌質量濃度。蘋果果實經體積分數70%乙醇溶液表面消毒后分為兩組,每組60 個果實,一組用綠原酸溶液浸泡30 min,另一組(對照組)用清水進行相同處理,浸泡后取出在通風條件下自然風干,然后誘導24 h。用直徑3 mm消毒釘在蘋果赤道部位陰陽兩面均勻刺2 個傷口(3 mm×3 mm),在傷口處分別接種20 μL 105個/mLB. cinerea懸浮液,晾干后覆上聚乙烯塑料薄膜(厚0.01 mm)保濕,貯藏于室溫(22±2)℃、相對濕度85%~90%的條件下。對照組和處理組分別固定30 個果實,用于統計病斑直徑和發病率,重復3 次。其余果實定期(0、6、12、24、48、72、120、168 h和216 h)取病斑周圍1~2 cm內健康果肉組織,用液氮迅速冷凍后磨成粉末,裝入錫箔紙袋中于-80 ℃保存,用于測定抗性相關代謝酶活力及活性物質含量。
1.3.2 指標測定
1.3.2.1 發病率和病斑直徑測定
病斑直徑利用游標卡尺進行十字交叉法測定,結果取平均值。蘋果病斑直徑不低于3.5 mm確定為發病,發病率按下式進行計算。

1.3.2.2 幾丁質酶、β-1,3-葡聚糖酶活力測定
幾丁質酶(chitinase,CHI)、β-1,3-葡聚糖酶(β-1,3-glucanase,GLU)活力分別參考袁仲玉[12]、曹建康[13]等的方法測定。稱取1 g果肉,加入5 mL pH 5.00.1 mol/L的乙酸-乙酸鈉緩沖液(含1 mmol/L乙二胺四乙酸二鈉和5 mmol/Lβ-巰基乙醇)冰浴研磨呈勻漿,離心(4 ℃、12000×g)后取上清液,即得到粗酶液。 以每分鐘每克鮮果肉中酶分解膠狀幾丁質產生1×10-9molN-乙酰葡萄糖胺為1 個CHI活力單位(U);以37 ℃下每克鮮果肉每分鐘催化底物昆布多糖產生1 nmol的葡萄糖為1 個GLU活力單位(U)。CHI和GLU活力單位均 為U/(min·g)。
1.3.2.3 苯丙氨酸解氨酶活力測定
苯丙氨酸解氨酶(phenylalanine amonialyase,PAL)活力測定參考Han Cong等[14]的方法。稱取1.0 g果肉,加入2 mL 0.1 mol/L pH 8.7的硼酸提取緩沖液(含40 g/L PVP、2 mmol/L乙二胺四乙酸二鈉、5 mmol/Lβ-巰基乙醇),冰浴條件下研磨成勻漿,4 ℃、12000×g離心30 min,取上清液,即得到粗酶提取液。以每小時每克鮮果肉組織酶促反應體系在290 nm波長處的吸光度增加0.01為1 個PAL活力單位(U),PAL活力單位 為U/(h·g)。
1.3.2.4 肉桂酸-4-羥化酶活力測定
肉桂酸-4-羥化酶(trans-cinnamic acid-4-hydroxylase,C4H)活力測定參考Liu Yaoyao等[15]的方法。稱取1.0 g果肉于研缽中,加入3 mL提取液(含50 mmol/L的Tris-HCl、15 mmol/Lβ-巰基乙醇、4 mmol/L MgCl2、5 mmol/L抗壞血酸、10 μmol/L亮抑酶肽、1 mmol/L PMSF、0.5 g/100 g PVP、體積分數10%丙三醇),研磨至勻漿,于4 ℃、12000×g離心20 min,得到上清液(粗酶液)。以每克鮮果肉組織酶促反應體系在340 nm波長處吸光度變化0.01為1 個C4H活力 單位(U),單位為U/(h·g)。
1.3.2.5 4-香豆酰輔酶A連接酶活力測定
4-香豆酰輔酶A連接酶(4-coumarate-CoA ligase,4CL)活力測定參照Takshak等[16]的方法并略有改動。稱取1.0 g果肉于研缽中,加入2 mL提取液(含0.2 mol/L Tris-HCl、8 mmol/L MgCl2、2 g/100 mL PVP、5 mmol/L DTT、體積分數0.1% Triton X-100、1 mmol/L PMSF)研磨至勻漿,于4 ℃、12000×g離心20 min,得到上清液(粗酶液)。以每分鐘每克鮮果肉組織于333 nm波長處吸光度變化0.01為1 個4CL活力單位(U),單位 為U/(min·g)。
1.3.2.6 總酚和類黃酮含量測定
總酚和類黃酮含量分別參考Toor[17]和González-Aguilar[18]等的方法測定。稱取1.0 g果肉,加入4 mL提取液(體積分數為1% HCl-甲醇,4 ℃預冷),研磨至勻漿,轉入離心管中,在4 ℃下提取15 min,于4 ℃、12000×g離心30 min。取上清液分別在280 nm和325 nm波長處測定OD值,并以OD280nm和OD325nm分別表征總酚和類黃酮含量。
1.3.2.7 木質素含量測定
按照周會玲等[19]的方法測定木質素含量,以每克鮮果肉在280 nm波長處的OD值表征木質素含量。
1.4 數據處理與分析
所有指標均平行測定3 次。以SPSS 17.0軟件進行數據分析,體外抑菌實驗數據采用單因素分析進行最小顯著性差異(least significance difference,LSD)分析,活體實驗數據采用t檢驗進行差異顯著性分析。用Excel 2007軟件作圖。
2 結果與分析
2.1 綠原酸對灰葡萄孢的體外抑制作用
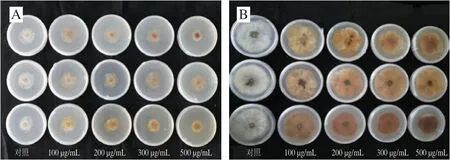
圖 1 綠原酸處理對體外接種B. cinerea菌落的影響Fig. 1 Effect of chlorogenic acid treatment on B. cinerea colonies in vitro
菌落是微生物在固體培養基上以母細胞為中心生長形成的肉眼可見子細胞集團,其直徑是反映微生物生長狀況的最直觀指標。如圖1所示,以含有0、100、200、300、500 μg/mL綠原酸的PDA培養基接種B. cinerea,接種后72 h,各處理組病斑直徑均小于對照組,菌落的顏色變為棕黃色,且隨著綠原酸質量濃度增加,菌落顏色加深。接種后120 h各處理組病斑直徑也均小于對照組,并且顏色由72 h時的棕黃色變成黃褐色,但不同質量濃度的綠原酸處理均未能完全抑制灰霉菌的生長。可見,綠原酸雖然不能完全抑制B. cinerea的生長,但與對照相比有抑制效果,并且能改變菌落的顏色,這可能是由于綠原酸處理影響了灰葡萄孢的正常生長。

圖 2 綠原酸對體外接種B. cinerea菌落直徑的影響Fig. 2 Effect of chlorogenic acid treatment on the diameter of B. cinerea colonies in vitro
由圖2可知,在PDA培養基中,綠原酸質量濃度在0~300 μg/mL范圍內增加時,灰葡萄孢菌落直徑顯著降低, 但當綠原酸質量濃度繼續增加到500 μg/mL時,菌落直徑不發生顯著變化。因此,確定300 μg/mL為最佳抑菌質量濃度,并將其作為本實驗綠原酸的使用質量濃度。
2.2 綠原酸對蘋果灰霉病的抑制作用
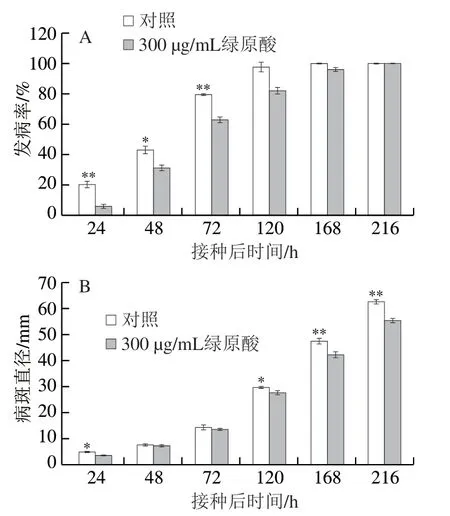
圖 3 綠原酸處理對蘋果灰霉病發病率(A)和病斑直徑(B)的影響Fig. 3 Effect of chlorogenic acid treatment on incidence (A) and lesion diameter (B) of gray mold on ‘Fuji’ apple fruit
發病率和病斑直徑是判斷發病情況的重要指標。從圖3A可以看出,接種后24 h,處理組和對照組果實均有發病,但對照組的發病率為20%,而處理組只有6%,差異極顯著(P<0.01)。此后對照組和處理組發病率均先迅速增加再緩慢增加,對照組和處理組分別在120、168 h時的發病率接近100%。由圖3B可以看出,對照組和處理組的病斑直徑在接種后72 h內均比較低,兩組無顯著差異,120 h后兩組病斑直徑均迅速增加,但處理組病斑直徑顯著低于對照組(P<0.05、P<0.01)。圖4為蘋果灰霉病發病的外觀照片。綜上,綠原酸處理能有效抑制灰霉病病斑擴展,從而減緩蘋果灰霉病的發生。

圖 4 綠原酸處理對蘋果灰霉病發病情況的影響Fig. 4 Effect of chlorogenic acid treatment on gray mold incidence on ‘Fuji’ apple fruit
2.3 綠原酸對蘋果果實CHI和GLU活力的影響
作為果實中重要的病程相關蛋白,CHI和GLU是受到病原菌侵染、機械損傷或環境脅迫等條件誘導寄主 產生防御反應的重要物質。由圖5A可知,接種灰葡萄孢后對照組和處理組CHI活力迅速增加,于48 h達到峰值,但此時處理組CHI活力比對照組高12.7%(P<0.01)。隨后對照組和處理組CHI活力有所降低,但整個貯藏期間處理組的CHI活力均高于對照組。由圖5B可知,對照組和處理組在貯藏過程中GLU活力總體上都表現出先增加再減小然后又增加的趨勢,但處理組GLU活力整體上高于對照組。接種灰葡萄孢后,處理組GLU活力迅速增加,6 h時即達到較高水平,而對照組GLU活力前期增加不明顯。兩組GLU活力都在72 h達到峰值,但此時處理組GLU活力極顯著高于對照組(P<0.01)。由此可見,綠原酸處理可顯著提高CHI和GLU活力,從而增強對病原菌的抑制作用,提高果實的抗病力。
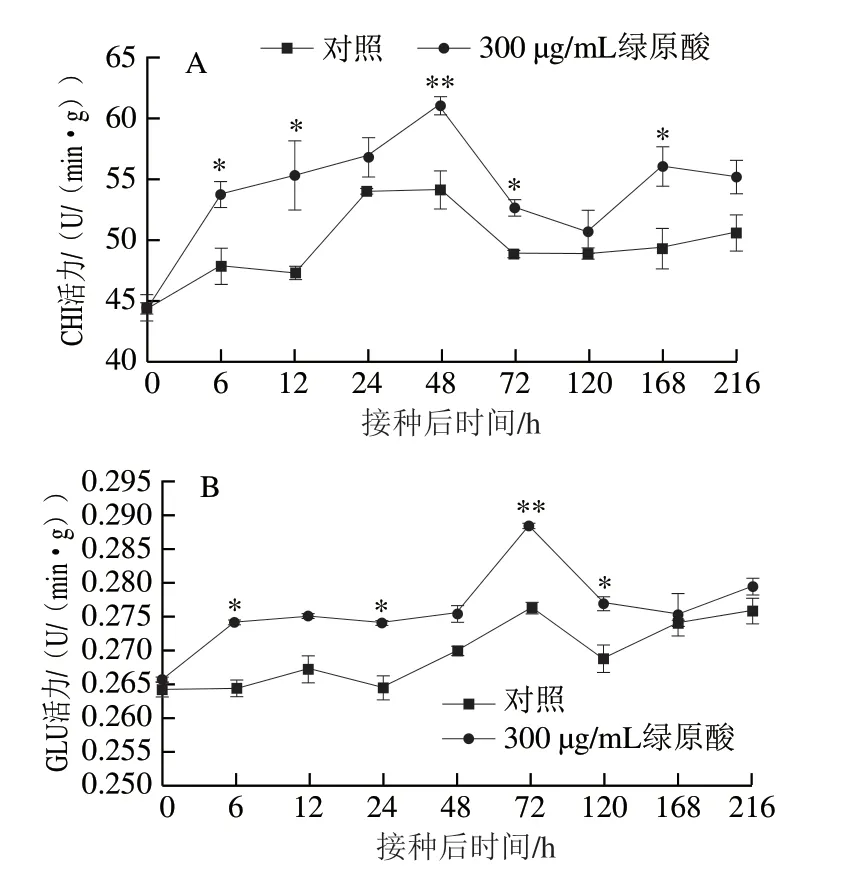
圖 5 綠原酸對接種灰葡萄孢后蘋果CHI(A)和GLU(B)活力的影響Fig. 5 Effect of chlorogenic acid on the activities of chitinase (A) and β-1,3-glucanase (B) in ‘Fuji’ apple fruit
2.4 綠原酸處理對苯丙烷代謝相關酶活力的影響
PAL與植物體內許多抗菌物質的生成密切相關,其活力是表征植物抗性的重要指標之一。如圖6A所示,整個貯藏過程中,處理組果實PAL活力均高于對照組,且兩組PAL活力變化總體上都表現為先升高后降低的趨勢。接種灰葡萄孢之后,處理組果實PAL活力迅速增加,在24 h即達到峰值,此時處理組PAL活力比對照組高6.7%(P<0.01)。對照組果實PAL活力增長較慢,48 h才達到活力峰值。在貯藏末期,對照組PAL活力仍有升高的趨勢,而處理組PAL活力開始降低,但處理組PAL活力高于對照組。
由圖6B可知,接種灰葡萄孢之后,對照組和處理組C4H活力總體上都表現出先升高后降低然后再升高的趨勢,且處理組C4H活力均高于對照組。兩組C4H的活力都在12 h達到峰值,但此時處理組C4H活力比對照組 高15.4%(P<0.01)。顯然,C4H在蘋果抵御灰霉病的抗性反應中產生了響應,且綠原酸處理對這一過程有促進作用。
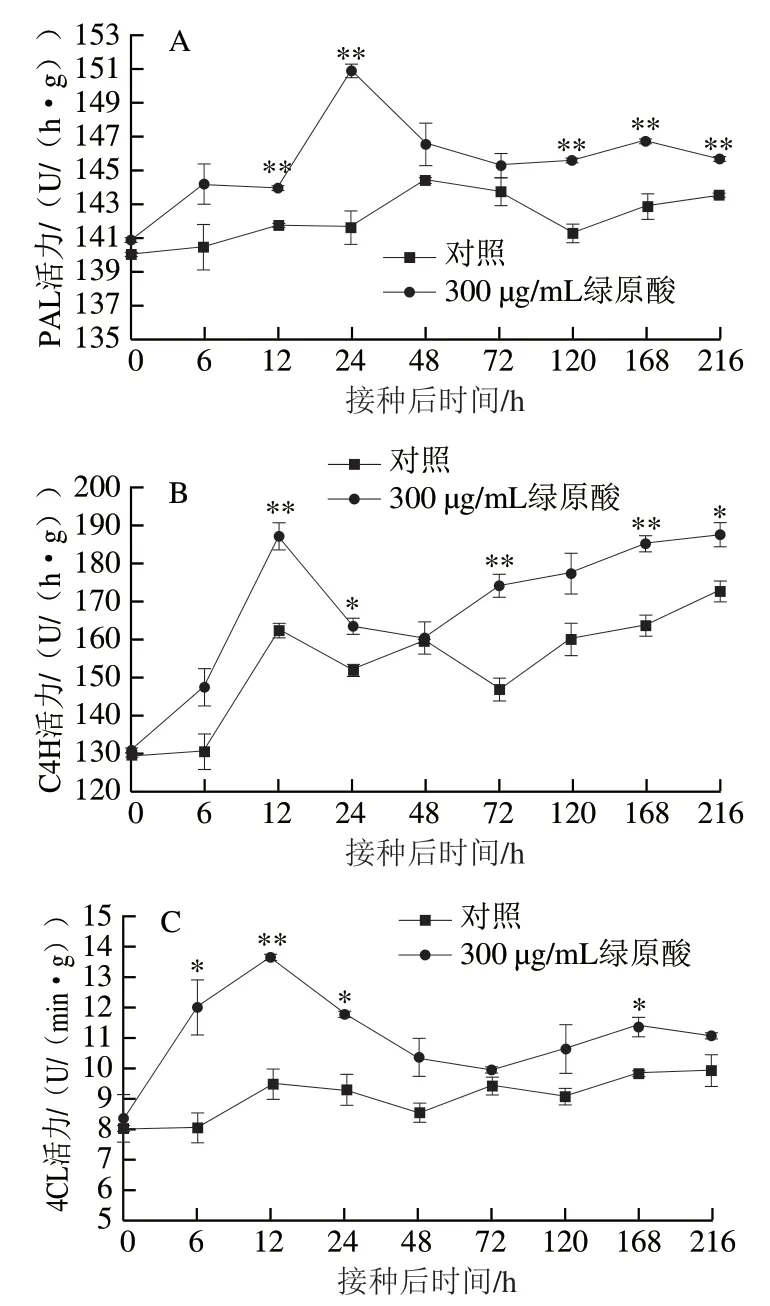
圖 6 綠原酸處理對接種灰葡萄孢后蘋果抗性相關酶活力的影響Fig. 6 Effect of chlorogenic acid treatment on the activities of gray mold disease resistance-related enzymes in ‘Fuji’ apple fruit
4CL是總酚、類黃酮、木質素等與抗性相關物質合成過程中一種關鍵性的限速酶,其活力常作為表征植物抗病性的一個重要指標。由圖6C可知,整個貯藏期間處理組果實4CL活力出現2 個峰值,而對照組4CL活力則表現出波動的變化趨勢,且無明顯的活力峰,并且處理組4CL活力始終高于對照組。處理組果實在接種灰葡萄孢后12 h時的4CL活力與對照組差異最大,比對照組高43.2%(P<0.01)。48 h之后兩組4CL活力之間的差異雖然不顯著,但處理組4CL活力仍高于對照組。由此可見,外源綠原酸處理可以提高4CL活力,且影響作用在接種后初期更為明顯。
2.5 綠原酸處理對苯丙烷代謝途徑次生物含量的影響
總酚和類黃酮是植物體內的直接抑菌物質,其含量與植物抗病能力密切相關。由圖7A可知,整個貯藏過程中,處理組蘋果果實總酚含量高于對照組。接種灰葡萄孢后對照組和處理組總酚含量均迅速增加,但貯藏初期兩組總酚含量差異并不顯著。隨著病害的發展,兩組 果實中總酚含量差異逐漸增大,到24 h時達到極顯著水平(P<0.01),此時處理組總酚含量比對照組高40%。接種24 h后,兩組總酚含量都開始緩慢減少,但216 h時但處理組總酚含量仍顯著高于對照組(P<0.05)。

圖 7 綠原酸處理對蘋果灰霉病抗性相關物質含量的影響Fig. 7 Effect of chlorogenic acid treatment on the contents of gray mold resistance-related substances in ‘Fuji’ apple fruit
由圖7B可知,整個貯藏過程中,處理組類黃酮含量均高于對照組。接種灰葡萄孢之后處理組類黃酮含量迅速增加,在12 h達到峰值,此時比對照組類黃酮含量高25%(P<0.01)。12 h后處理組類黃酮含量出現一個短暫的降低過程,但隨后又開始升高,并且直到貯藏末期,處理組類黃酮含量仍極顯著高于對照組 (P<0.01)。可見,與對照組相比,綠原酸處理可以明顯提高蘋果對灰霉病的抗性反應過程中類黃酮的含量。
木質素可以強固細胞壁和增加抗性,是植物抗性系統的重要組成部分。由圖7C可知,接種灰葡萄孢后,處理組木質素含量迅速增加,于72 h達到最高,此后有所降低,而對照組的木質素含量上升比較緩慢,120 h達到最高,之后處于相對穩定狀態。但整個過程處理組木質素含量明顯高于對照組。可見,與對照組相比,綠原酸處理可以使蘋果在抵抗灰霉病的抗性反應過程中積累更多的木質素。
3 討 論
綠原酸是植物體內普遍存在的一種與抗性密切相關的酚酸類物質[20]。研究表明,抗病性強的蘋果品種含有的綠原酸較多,且在受到病原菌侵染后,其綠原酸增加量顯著高于抗性低的蘋果品種[21]。李珊珊等[22]研究表明,綠原酸對西瓜枯萎病菌有較強的離體抑制作用,且隨著濃度的升高,其抑菌活性呈現出增強的趨勢。本研究發現,綠原酸處理能明顯降低蘋果灰霉病發病率,抑制病斑直徑,且在接種前期表現較為突出。可能是由于前期用綠原酸浸泡處理導致部分綠原酸滲入果實內部,從而對灰葡萄菌產生直接抑制作用,因此在接種后24 h,處理組的發病率與對照組有極顯著差異 (P<0.01),此時處理組病斑直徑也顯著低于對照組(P<0.05)。隨著綠原酸在果實內部被代謝,其濃度逐漸降低,處理組發病率與對照組差異逐漸縮小。發病末期,處理組病斑直徑顯著低于對照組,可能是由于外源綠原酸處理提高了果實自身抗病能力,從而抑制了病斑擴散情況。
植物體內的CHI和GLU在抵御病原菌侵染的防衛反應中主要起細胞壁水解酶的作用[23]。Schlumbaum等[24]研究發現,菜豆積累的CHI經過提純后,在體外能夠明顯抑制綠色木霉(Trichoderm viride)菌絲的生長。正常情況下,CHI和GLU主要分布于植物細胞液泡內,細胞間隙中相對較少[12],且活性很低,當病原菌侵染植物后會對細胞膜造成損害,從而誘導CHI和GLU活性升高[25]。本實驗中,接種灰葡萄孢后,處理組CHI和GLU活力迅速增加,且在整個貯藏期間均大于對照組,有效提高了蘋果的灰葡萄孢抗性,與袁仲玉[12]、周曉婉[26]等利用1-甲基環丙烯、蘆薈粗提物誘導蘋果灰霉病抗性的結果一致。
苯丙烷代謝是植物抗病反應中重要的代謝途徑之一。PAL是苯丙烷代謝途徑中第一個催化產生酚類的酶,可催化苯丙氨酸生成肉桂酸,對酚類、植物抗毒素和木質素的生物合成具有調節作用[27]。C4H可以催化肉桂酸生成4-香豆酸,是苯丙烷代謝途徑中重要的節點酶之一[28]。4CL催化對香豆酸形成的對香豆酸輔酶A是多個重要代謝分支的前體物質,可進一步合成綠原酸等酚酸類,也可在查耳酮合成酶的催化下形成查耳酮和根皮苷等類黃酮類[29]。PAL、C4H和4CL作為苯丙烷代謝途徑上游的幾個關鍵節點酶,負責催化多種物質的合成[30], 為整個代謝途徑的眾多下游分支提供前體物質,其活性與植物的抗性密切相關。經外源綠原酸處理的果實在接種灰葡萄孢后,PAL、C4H、4CL活力迅速升高,在很短時間內達到較高水平,可以更快地積累與抗病性相關的次生代謝產物,從而提高果實的抗病性。
作為苯丙烷途徑的次生代謝產物,總酚和類黃酮具有殺菌、抗病毒、清除自由基等多種與植物抗病性相關的生物活性[31]。Deng Jianjun[32]和Li Guangjin[33]等研究發現,苯丙烷代謝途徑關鍵酶活性的增加可以促進總酚、類黃酮的積累,從而增強桃、甜瓜對病原菌的抗性。本研究發現,PAL、C4H、4CL活力升高的同時,總酚、類黃酮含量也增加,綠原酸處理組果實中總酚和類黃酮含量明顯高于對照組,這與Terry等[34]的研究結果一致。本實驗中,接種灰葡萄孢后,綠原酸處理組和對照組木質素含量都明顯增加,但處理組木質素含量增加更多。木質素可以與細胞壁多糖、細胞壁蛋白結合,增厚細胞壁形成機械屏障,阻止病原菌侵入及其在植物體內的擴展[35]。
綜上,與對照相比,外源綠原酸處理可以明顯提高果實中CHI和GLU活性,抑制灰葡萄孢的生長,同時提高苯丙烷代謝途徑中PAL、C4H、4CL等關鍵酶活性,促進次生代謝產物總酚、類黃酮和木質素的積累,從而提高果實的抗病性,有效改善接種果實發病情況。
