玉米轉錄因子基因ZmMYB308的克隆及表達分析
2021-06-22 00:46:30王新濤代資舉李保葉郝俊杰
西北植物學報 2021年5期
關鍵詞:分析
王新濤,楊 青,代資舉,李保葉,郝俊杰*
(1 河南省農業科學院 植物保護研究所,鄭州450002;2 河南省農業科學院 芝麻研究中心, 鄭州450002; 3 河南省作物分子育種研究院, 鄭州450002)
玉米是中國最重要的糧食作物之一,近年來玉米生產規模化程度提高,對于玉米品種適應全程機械化的要求也越來越迫切,而玉米莖稈倒伏是制約中國玉米機械化收獲發展的主要因素之一[1];同時隨著全球極端氣候發生頻率的不斷增多,對于玉米品種的穩產抗倒性要求也越來越高。研究表明,全世界每年因倒伏引起的玉米產量損失估計達到5%~20%,倒伏不僅嚴重影響玉米產量而且制約著玉米機械化收獲程度的提高[2-4]。木質素是構成次生細胞壁的主要成分之一,適當提高木質素含量可以增強玉米莖稈抗折力,從而降低倒伏引起的損失[5-6]。
MYB轉錄因子作為植物中重要的轉錄因子家族之一,廣泛地參與調節了植物生長發育的許多重要過程,如調控基因轉錄、維持細胞結構、信號通路傳導和生物及非生物脅迫的生理響應等[7]。MYB類蛋白的N端通常含有約51~52個氨基酸組成的高度保守的MYB結構域,C端為轉錄激活結構域;根據所含MYB結構域的數量和位置,MYB 轉錄因子可分為1R-MYB、R2R3-MYB、3R-MYB 和4R-MYB四個亞類:1R-MYB又被稱為MYB-related亞類,只包含一個R結構域,在維持細胞完整性和周期節律方面發揮著重要作用;R2R3-MYB是數量最多的亞類,廣泛參與植物的生長發育和代謝途徑調節等過程;3R-MYB亞類在植物中的數量相對較少,它們主要是參與細胞周期運行以及細胞分化;4R-MYB亞類在MYB轉錄因子基因家族中是最小的一類,對其功能的研究目前還很少[8]。
通過苯丙烷途徑合成的木質素是構成植物骨架的主要成分之一,多個MYB轉錄因子在木質素合成途徑中起著重要的調節作用[9]。擬南芥中次生細胞壁相關蛋白AtSND1通過調控下游的AtMYB83和AtMYB46,進而誘導AtMYB58、AtMYB63和AtMYB85的表達,最終引起相關木質素合成基因上調[10-11];桉樹MYB2蛋白通過直接與苯丙烷途徑中CCR和CAD基因的啟動子結合,從而影響木質素和次生壁的形成[12];水稻OsMYB46在擬南芥中過表達可以激活次生壁的合成途徑[13]。這些不同的MYB成員通過正向或負向作用機制有效調控木質素的合成,因此,加強對MYB家族基因的相關研究具有重要的現實意義。
玉米MYB轉錄因子在莖稈中參與木質素生物合成的研究相對較少,對ZmMYB308基因的研究還未見報道。本研究利用玉米自交系‘鄭58’不同發育階段的莖稈轉錄組數據,初步篩選出差異表達的MYB轉錄因子并進行分析,以‘鄭58’莖稈為材料,克隆了玉米ZmMYB308基因,對該基因進行了生物信息學特征分析,利用熒光定量PCR分析ZmMYB308在玉米不同部位與不同發育時期的表達模式,以期為玉米高莖稈強度基因篩選及分子育種應用提供依據。
1 材料和方法
1.1 材 料
以玉米優良自交系‘鄭58’為材料[14],種植行長2 m,行距0.6 m,株距0.25 m。2018年6月在河南省農業科學院原陽基地播種,出苗后正常田間管理,分別取6葉期、拔節期、大喇叭口期、吐絲期和灌漿期(吐絲后10 d)的玉米莖稈,在玉米吐絲期取莖、根、葉、花絲、雄穗和雌穗等不同組織為樣品,經液氮速凍后于-80 ℃超低溫冰箱保存備用,每個樣品3個生物學重復。
1.2 方 法
1.2.1 莖稈穿刺強度測定在玉米吐絲期和灌漿期進行莖稈穿刺強度測定,利用YYD-1型數顯植物莖稈強度測定儀(浙江托普儀器有限公司)選擇地上部第3節間中部,對玉米莖稈進行穿刺,瞬間讀取并記錄數據(N/mm2),每個時期連續選取5株進行測定。
1.2.2 轉錄組測序及差異表達基因篩選根據‘鄭58’莖稈強度變化,選擇其具有顯著差異時期(抽雄期和灌漿期)的莖稈(地上部第3節間)進行采樣,采集的樣品(3次生物學重復)送至北京諾禾致源科技股份有限公司進行轉錄組測序和分析。
分別提取各樣品總RNA并檢測合格后,依次進行Oligo(dT)磁珠富集、mRNA反轉錄、cDNA文庫構建和Illumina平臺測序,參考玉米B73(RefGen_v4)基因組序列,進行序列組裝、比對和功能注釋等相關分析。基因表達量采用FPKM (fragments per kilobase of exon model per million mapped reads)計算[15],利用DEseq分析差異基因,以差異倍數|log2(Fold change)|>1且錯誤檢出率(FDR)<0.05為標準篩選吐絲期和灌漿期莖稈中差異表達的MYB轉錄因子基因。
1.2.3 玉米RNA提取及cDNA第1鏈合成各樣品經液氮研磨后利用TRNzol法提取總RNA,1%瓊脂糖凝膠電泳檢測RNA完整性。以不同玉米樣品總RNA為模板,使用PrimeScript RT-PCR Kit(TaKaRa)試劑盒,反轉錄獲得玉米cDNA第一鏈。
1.2.4 玉米ZmMYB308基因的克隆根據轉錄組篩選的差異表達候選基因及在NCBI上比對篩選,在GenBank中獲得一條具有完整開放閱讀框(open reading frame, ORF)的玉米MYB308序列,利用Primer Premier 5.0軟件設計用于擴增ZmMYB308特異引物ZmMYB308-F(TCTTCCGTTTCCGTCTCCC)和ZmMYB308-R(CGACG-ACGCCTGCCTTAT)。以‘鄭58’莖稈cDNA為模板進行全長擴增。
1.2.5 生物信息學分析利用NCBI的ORF finder (https://www.ncbi.nlm.nih.gov/orffinder/)工具預測基因ORF區、CCD (https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)工具進行蛋白序列保守結構域分析;利用cNLS Mapper(http://nls-mapper.iab.keio.ac.jp/)分析基因核定位信號肽;使用ExPAsy的Compute pI/MW(http://www.expasy.org/tools/pi_tool.html)工具計算蛋白序列的分子量及等電點;利用DNAMAN9軟件進行蛋白翻譯和序列比對。
1.2.6 熒光定量PCR表達分析根據轉錄組篩選獲得的差異候選基因序列設計熒光定量引物,玉米肌動蛋白基因(β-Actin)為內參基因(表1)。以玉米不同部位和不同時期的莖稈cDNA為模板,反應體系為15 μL,其中cDNA模板1 μL,上下游引物各0.6 μL、SYBR Premix ExTaq(TaKaRa) 7.5 μL、ddH2O 5.3 μL。在熒光定量PCR儀(Bio-Rad Miniopticon Real-Time PCR System)上進行擴增,定量PCR程序:95 ℃預變性2 min,95 ℃變性10 s,58 ℃退火30 s,72 ℃延伸25 s,并收集熒光信號,40個循環。基因的相對表達量釆用2-ΔΔCT算法進行分析。
2 結果與分析
2.1 差異表達MYB轉錄因子的篩選及分析
‘鄭58’在吐絲期的莖稈穿刺強度為25.87 N/mm2,在灌漿期的莖稈穿刺強度為30.23 N/mm2。對吐絲期與灌漿期的莖稈MYB轉錄因子基因數目進行統計分析(表2),獲得14個差異表達MYB轉錄因子基因,分別位于玉米的1、2、3、5、6、7、8和10染色體,其中10個表達量下降,4個表達量上升。熒光定量PCR分析結果(圖1,部分結果)顯示,MYB39/MYB59(下調)和MYB4/MYB126(上調)在吐絲期和灌漿期莖稈中的表達趨勢與轉錄組數據分析結果趨勢相符,驗證了轉錄組基因表達數據的可靠性。
根據SANT保守結構域的數目進行分類,這些差異表達的MYB轉錄因子包含2個1R-MYB和12個R2R3-MYB亞類,沒有發現3R-MYB和4R-MYB。對蛋白序列分析發現,莖稈差異表達的MYB蛋白家族分子量在27.01~50.1kD之間,等電點為4.58~9.17。這些結果說明隨著玉米莖稈的生長發育,各MYB轉錄因子的表達出現差異化。

表1 熒光定量PCR分析的候選基因及其引物序列

表2 差異表達MYB轉錄因子基因信息

不同小寫字母代表0.05水平差異顯著圖1 玉米吐絲期和灌漿期MYB部分基因的相對表達Different normal letters represent significant differences at 0.05 levelFig.1 Relative expression of some MYB genes in the silking stage and grain filling stage
2.2 玉米ZmMYB308基因cDNA的克隆
根據以上轉錄組差異基因分析數據,再結合其他植物MYB308類基因在莖稈木質素合成途徑中的負調控作用模式[16-17],選取在吐絲期和灌漿期顯著下調表達的玉米MYB308基因進行后續試驗分析。以玉米自交系‘鄭58’的莖稈cDNA為模板,經PCR擴增獲得玉米ZmMYB308的cDNA序列(圖2)。其cDNA全長907 bp,包含一個747 bp的ORF,編碼248個氨基酸。通過MaizeGDB數據庫Blast序列分析發現,ZmMYB308基因位于玉米第8染色體的bin8.08區域,包含2個外顯子和1個內含子。

M. DL5000;1.cDNA圖2 ZmMYB308基因cDNA擴增Fig.2 PCR product of ZmMYB308 cDNA
2.3 玉米ZmMYB308基因的生物信息學分析
經ExpasyProtParam預測ZmMYB308蛋白分子量為27.01 kD,等電點為9.17,分子式為C1179H1874N354O349S13,不穩定系數為61.67,蛋白疏水性指數為-0.533,脂肪系數為 69.8,說明該蛋白為不穩定的親水性蛋白。利用cNLS Mapper預測在蛋白N端有一段核定位信號(圖3)。通過NCBI分析CDD功能結構域發現ZmMYB308蛋白N端具有2個保守的 DNA 結合結構域(圖4),屬于典型的R2R3-MYB亞類轉錄因子。

SANT 結構域,Myb DNA-binding結構域,REB1超家族圖4 ZmMYB308蛋白功能性位點分析SANT domain, Myb DNA-binding domain, REB1 superfamilyFig.4 Conserved domain analysis of ZmMYB308 protein

下劃線為核定位信號圖3 ZmMYB308基因開放閱讀框及預測氨基酸序列The nuclear localization signal is underlinedFig.3 ORF of ZmMYB308 gene and predicted amino acid sequence
2.4 ZmMYB308同源及進化分析
利用NCBI數據庫的BlastP在線檢索篩選其他植物的 MYB308蛋白序列,選取了桉樹(XP_010090332.1)、白楊(XP_009376958.1)和二穗短柄草(XP_003564745.1)等共14個同源序列,通過DNAMAN9.0將玉米ZmMYB308與其他植物的MYB308蛋白序列對比,建立了一個無根分子系統進化樹(圖5)。比對顯示MYB308 類蛋白序列在N端高度保守,而C端的序列保守性較低,系統進化分析將15個不同植物的MYB308蛋白劃分為2組,其中ZmMYB308與谷子、高梁和二穗短柄草等MYB308蛋白聚為一類,且與谷子的相似性最高,達到94%。
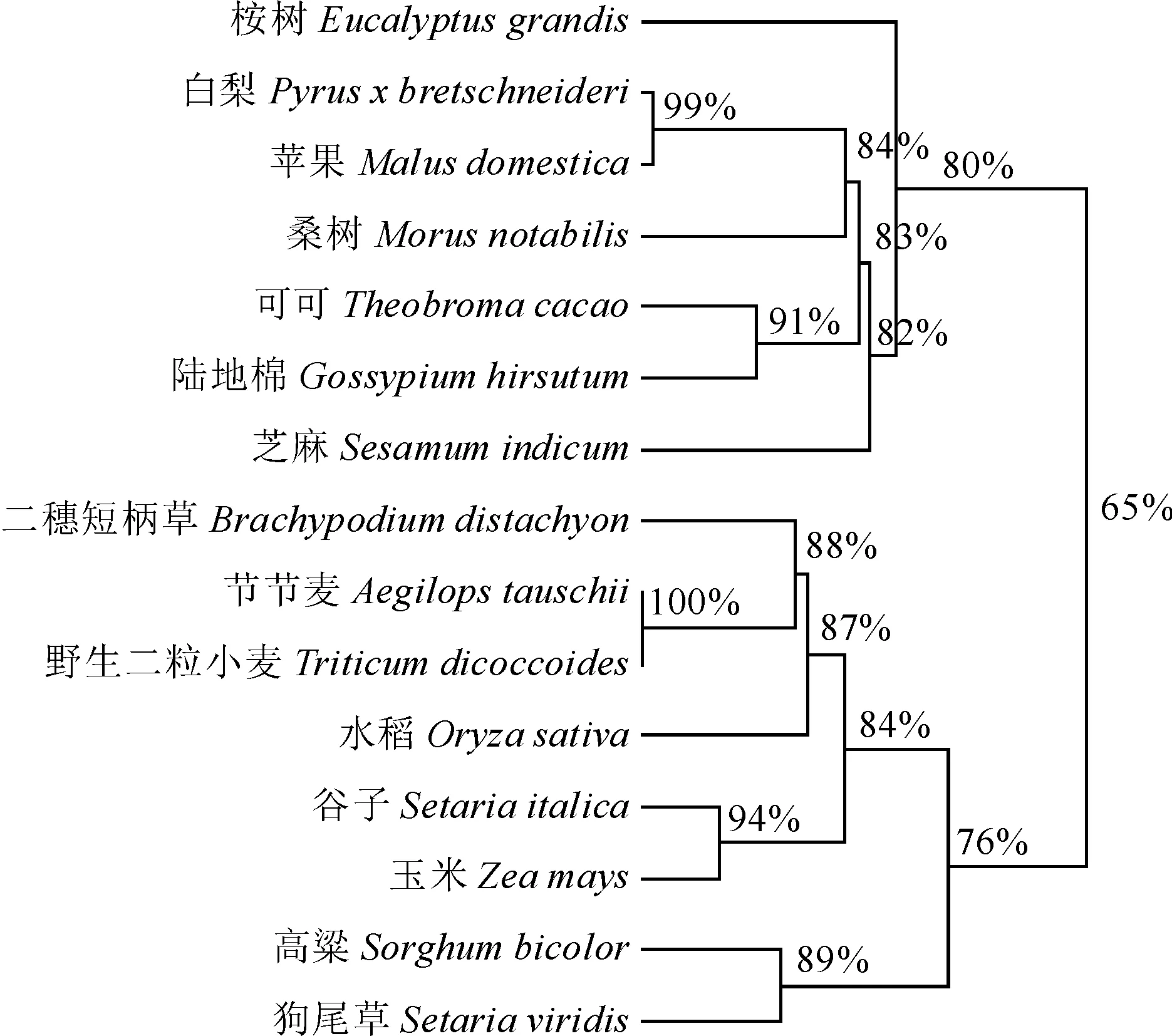
圖5 不同物種間MYB308蛋白的進化樹Fig.5 Phylogenetic tree of MYB308 protein homologues from different species
2.5 玉米ZmMYB308基因的表達分析
為揭示玉米不同發育時期在莖稈中ZmMYB308基因的表達特性,分別對‘鄭58’不同發育時期莖稈中該基因的表達進行分析,結果(圖6)表明,ZmMYB308隨著玉米發育的推進呈先升高后降低的趨勢,在吐絲期表達量最高,隨后在灌漿期下降,其變化趨勢與莖稈轉錄組測序結果趨勢一致。
為了揭示ZmMYB308在玉米不同組織中的表達情況,在玉米吐絲期對不同組織進行實時熒光定量分析,結果(圖7)表明ZmMYB308在玉米不同組織中均有表達,在玉米莖稈中的表達量最高,根、雌穗、雄穗、葉片中的表達量次之,花絲中的表達量最低,其莖稈中的表達量約為花絲中的7.5倍。
3 討 論
隨著轉錄組測序技術的不斷進步,在基因水平上對整體轉錄活動進行實時檢測,從而完整分析不同的代謝通路和驗證基因功能變得越來越高效快捷[18-19]。近年來,人們在擬南芥、水稻和玉米等多種植物的不同發育過程進行了大量的轉錄組分析,從不同角度解析了目的基因及其調控基因的調控網絡[20-22]。本研究通過轉錄組測序和分析,在不同發育時期的玉米莖稈中鑒定出14個差異表達MYB轉錄因子,在此基礎上,采用RT-PCR技術克隆了玉米R2R3-MYB類轉錄因子ZmMYB308基因,并對其生物信息學與時空表達特性進行了分析。
2R-MYB亞類作為MYB轉錄因子的主體,可以通過激活木質素合成途徑的相關基因來實現對木質素合成的正向調控:在擬南芥中,MYB58能夠直接激活木質素生物合成基因(PAL1、C4H和4CL1等)和次生壁相關漆酶基因(LAC4)的表達進而參與了木質素的生物合成途徑[10];在楊樹中,過量表達PtrMYB2、PtrMYB3、PtrMYB20和PtrMYB21基因可以引起次生壁的異位沉積,導致次生壁增厚[23]。也有多個2R-MYB亞類轉錄因子是負向調控木質素合成的:在擬南芥中過表達玉米ZmMYB31、ZmMYB42基因可以抑制木質素合成途徑中的多個基因,從而降低組織中的木質素含量[24-25];銀合歡LiMYB1過表達抑制了苯丙烷途徑基因的表達水平,從而引起木質素含量的降低[26];金魚草中AmMYB308和AmMYB330共同作用可降低木質素的積累[16]。本研究通過轉錄組分析玉米吐絲期和灌漿期莖稈中MYB轉錄因子表達量變化,鑒定出差異表達的MYB轉錄因子中2R-MYB亞類占到85.71%,因此推測這些差異表達的MYB轉錄因子,特別是2R-MYB亞類轉錄因子正向或負向地參與了玉米莖稈木質素的生物合成,進一步克隆這些玉米的MYB類轉錄因子,將有助于全面了解這類基因的功能。

不同小寫字母代表0.05水平差異顯著圖6 ZmMYB308在玉米不同發育時期的表達Different normal letters represent significant differences at 0.05 levelFig.6 The expression pattern of ZmMYB308 in different development stages

不同小寫字母代表0.05水平差異顯著圖7 ZmMYB308在不同組織的表達Different normal letters represent significant differences at 0.05 levelFig.7 Relative abundance of ZmMYB308 transcripts in different tissues
作為MYB家族成員的MYB308及其同源基因在多種作物中被證實能夠調控影響木質素合成途徑的相關基因[16-17]。本研究從玉米中克隆得到ZmMYB308基因,生物信息學分析顯示ZmMYB308蛋白與其他已報道的MYB308類基因的蛋白保守結構域具有較高的相似性,在其N端具有家族典型的MYB保守結構域和核定位信號,玉米ZmMYB308轉錄因子可能在細胞核中與其他基因互作并調控其下游基因。ZmMYB308基因在不同部位是組成型表達的,在莖中表達量最高,并且ZmMYB308隨著莖稈發育在吐絲期達到峰值后下降,同PgMYB308基因在石榴中的表達趨勢類似[24]。結合‘鄭58’莖稈強度的表型變化,ZmMYB308基因可能和金魚草AmMYB308、石榴PgMYB308基因一樣在木質素合成過程中起著負調控作用。研究還發現ZmMYB31表達受低溫誘導,過表達ZmMYB31能增強植株對低溫脅迫的耐受性[27],蘋果MdMYB308基因在調控蘋果花青苷合成中起著重要作用[28],小桐子JcMYB308基因在根與莖中對低溫做出響應[29],紫斑白三葉TrMYB308參與類黃酮次生代謝生物合成調控[30],本研究中ZmMYB308在根、雌穗中的表達量高于葉片和花絲,暗示了玉米ZmMYB308基因可能還具有其他重要功能和作用機制,需要進一步深入研究。
本研究通過RNA-Seq測序和數據分析發現,在獲得的玉米莖稈轉錄組數據中除MYB轉錄因子外還存在其他差異表達基因,挖掘和解析這些重要基因的功能有助于進一步提高對玉米木質素合成機制的認識,也為利用分子生物學手段培育高莖稈強度玉米品種奠定了基礎。
作者貢獻:王新濤和郝俊杰是本研究的實驗設計和實驗研究的執行人;王新濤和李保葉完成數據分析和論文初稿的寫作;楊青和代資舉參與實驗設計與實驗所得數據分析;郝俊杰是本研究的的構思者及負責人,指導數據分析,論文寫作與修改。全體作者都閱讀并同意最終的文本。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
