無菌豬和普通豬早期脂肪發育及脂肪組織基因轉錄表達的差異
2021-06-23 07:55:32邱小宇劉作華齊仁立
生物技術通報 2021年5期
關鍵詞:差異
邱小宇 劉作華,2 齊仁立,3
(1.重慶市畜牧科學院,重慶 402460;2.農業農村部養豬科學重點實驗室,重慶 402460;3.重慶市養豬科學重點實驗室,重慶 402460)
動物的胃腸道內寄居著數量巨大、種類繁多的微生物,包括細菌、真菌、古菌、病毒等,其中90%以上為細菌,涵蓋了數千種不同的菌屬[1]。成年豬的腸道細菌數量高達1014CFU/g,厚壁菌門(Firmicutes)和擬桿菌門(Bacteroidete)細菌占全部腸道菌的95%以上[2]。腸道菌群按照一定比例共存,相互依賴、相互約束,最終在宿主體內形成一個復雜的微生態系統[3]。現有的研究發現,腸道微生物可以調控宿主體內多個組織或器官的發育與成熟,如腸道、肝臟、大腦、免疫系統等都會受到腸道微生物的直接或者間接影響[4-6]。而腸道微生物的失衡以及有害細菌的侵襲則會破壞宿主體內的生理穩態,甚至會導致宿主出現肥胖病[7]、糖尿病[8]、帕金森病[9]和抑郁癥[10]等多種疾病。
脂肪組織是動物機體重要的組織器官,功能包括:調節體溫、保護內臟器官,作為“燃料庫”為動物儲存和提供能量。脂肪還可以通過多種方式(吸收脂溶性維生素、供給必須脂肪酸、分泌多種細胞因子等)干預和調控體內其他組織器官的生理功能[11-12]。當前,大量的研究結果表明,腸道微生物主要通過兩種方式影響宿主脂肪組織的發育和功能:(1)直接影響宿主消化道內多糖等物質的發酵以及能量物質的產生與獲取[13];(2)經由細菌代謝物(如短鏈脂肪酸等)調控脂肪代謝相關基因的表達進而影響脂肪細胞分化和成熟[14]。然而,目前腸道微生物干預和調控脂肪發育的研究多集中在無菌小鼠、悉生小鼠等嚙齒類模型動物上,對豬、牛、羊等具有重要經濟價值的家畜上的相關研究還相對較少。
無菌動物是研究腸道微生物調節宿主生長發育的重要模型工具。在以往的研究中,無菌豬主要用于探索腸道微生物群對免疫系統發育和成熟的影響[15-17],而利用無菌豬來揭示腸道微生物群對豬脂肪組織發育的相關研究鮮有報道。此外,由于豬的腸道微生物組成和代謝模式與人有著較高的生理相似度[18],因此基于豬腸道微生物的研究對人的生理和病理相關研究也會具有一定指導意義。本研究選用無菌仔豬(germ free piglets,GF piglets)和普通帶菌仔豬(normal piglets)進行比對分析,比較兩種豬的脂肪組織形態、功能和基因表達的差異,揭示腸道菌定植對仔豬早期脂肪沉積和脂肪代謝的影響。研究的相關結果對于通過腸道微生態平衡改善動物生長和生產性能、維持動物生理健康能提供一定的理論依據。
1 材料與方法
1.1 材料
選擇無血緣關系,身體健康,外觀無異常,血清檢測豬瘟病毒、非洲豬瘟病毒、口蹄疫病毒、豬繁殖與呼吸綜合征病毒、乙型腦炎病毒、偽狂犬病病毒、豬圓環病毒2型、豬細小病毒、布魯氏菌、豬肺炎支原體、豬流感病毒等病原微生物均為陰性的妊娠母豬在預產期前3 d進行無菌剖腹產手術(無菌豬模型制備參照中國實驗動物學會團體標準TCALAS71-2019執行,3頭無菌豬由重慶市畜牧科學院無菌豬研究平臺提供)。將剖腹獲得的無菌仔豬飼養于無菌的隔離器內。無菌豬的飲用水和配方奶粉均經過60Co輻照且微生物檢測合格(飼料、飲用水微生物檢測參考孫靜等[19]的方法執行)。無血緣關系的3頭普通仔豬在標準化的飼養環境中出生和生長,正常母乳喂養,常規飼養管理。
1.2 方法
1.2.1 樣品采集 在25日齡時,3頭無菌仔豬和3頭普通仔豬分別通過異氟烷呼吸麻醉,放血處死,在潔凈環境內進行采樣。迅速切開仔豬頸部皮膚組織,使用游標卡尺測量頸部皮下組織的脂肪厚度。用無菌剪刀采集1.5 cm×1.5 cm×0.2 cm大小的頸部皮下脂肪組織,預冷的生理鹽水漂洗3次后,分別放入4%中性甲醛中固定和液氮速凍后-80℃冰箱保存。
1.2.2 石蠟切片和HE染色 將固定24 h后的脂肪組織從4%的甲醛溶液中取出,用超純水反復沖洗12 h后置于75%乙醇內,用70%、80%、90%、100%的梯度乙醇脫水,脫水后使用二甲苯透明、石蠟溶化后包埋,切成3 μm-5 μm厚的薄片后行進行HE染色,蒸餾水洗5 min,蘇木精染色6 min,自來水洗凈,分化液20 s,水中浸泡20 min返藍,1%伊紅質染1 min,自來水沖洗,脫水、透明、封片,置于顯微鏡下觀察脂肪組織細胞直徑大小和脂肪沉積差異。
1.2.3 蛋白提取和免疫印跡雜交 脂肪組織液氮研磨成粉末狀后,加入含有1 mmol/L PMSF的RIPA蛋白裂解液,4%搖床過夜。充分裂解后,12 000 r/min,4℃離心5 min,吸取上清液并檢測蛋白濃度。將所有蛋白樣品調至等濃度,加入Loading buffer充分混勻,98℃金屬浴10 min后進行SDS-PAGE電泳與轉膜。轉膜后用5%的脫脂奶粉室溫封閉1 h,加入目標蛋白的一抗,4℃孵育過夜。第2天用TBST室溫搖洗3次,每次5 min。加入HRP標記二抗,室溫孵育1 h,TBST搖洗3次,每次5 min。使用ECL反應試劑盒,化學發光自動成像儀曝光拍照。Image J軟件對蛋白條帶進行定量分析。
1.2.4 酶聯免疫檢測 脂肪組織加入適量預冷的生理鹽水研磨粉碎,1 000 r/min離心后取上清液,置于-20℃保存待測。使用ELISA檢測試劑盒測定脂肪細胞因子-脂聯素(adiponectin)和瘦素(leptin)的含量(上海酶聯生物有限公司),操作步驟嚴格按照說明書進行。
1.2.5 轉錄組測序與生物信息學分析 使用試劑盒提取脂肪組織總RNA,Nanodrop 2000檢測RNA濃度和純度,瓊脂糖凝膠電泳檢測完整性,Agilent2100測定RIN值。利用帶有Oligo(dT)的磁珠分離mRNA,通過Illumina HiSeq平臺對其序列進行測序。測序完成后,對所獲得的測序數據進行質量控制,去除含有接頭的reads、全部都是A堿基的reads、含N比例大于10%的reads和低質量的reads(質量值Q≤20的堿基數占整條reads的50%以上)。
使用比對軟件Tophat2(2.1.1)將得到的序列對比到豬的參考基因組上。利用FPKM(fragments per kilobase of transcript per million mapped reads)計算法,統計轉錄本的表達量及其分布情況,并根據FPKM值分析樣本兩兩之間的相關性。差異表達基因篩選采用DEGseq2軟件并以RPKM(reads per kilobase transcriptome per million mapped reads)計算方法進行,篩選條件為P<0.001,FC>2,至少一組中的RPKM>1(FC為3個樣品表達量的平均值)。對篩選出的差異基因進行KEGG(kyoto encyclopedia of genes and genomes)通路富集分析,應用超幾何檢驗尋找顯著性富集的通路,確定差異表達基因參與的主要生化代謝途徑和信號轉導通路。
1.2.6 qRT-PCR驗證轉錄組測序結果 從轉錄組測序的結果中挑選具有明顯表達變化的脂肪代謝相關調控基因,使用qRT-PCR的方法進行驗證。將測序所用的total RNA樣品使用PrimeScriptTMRT reagent Kit(TaKaRa,日本)試劑盒反轉錄為cDNA。使用Q6熒光定量PCR儀(AB 公司,美國)進行PCR反應,反應條件:95℃預變性30 s;(95℃ 5 s,60℃35 s)40 個循環。U6 為參照基因,2-△△CT法計算基因的相對表達量。
1.2.7 數據統計 實驗數據以Mean±S.E.形式表示,用Graphpad Prism 7.0軟件進行Student t檢驗和作圖。*P<0.05為顯著,**P<0.01為極顯著。
2 結果
2.1 無菌豬和普通豬的脂肪組織發育比較
圖1-A顯示了普通仔豬和無菌仔豬脂肪細胞的形態差異。與具備完整腸道微生物的普通仔豬相比,相同日齡無菌仔豬的脂肪沉積量明顯較少。無菌仔豬的頸部皮下脂肪厚度、脂肪細胞直徑顯著低于普通仔豬(P<0.001;圖1-B,圖1-C)。脂肪酸結合蛋白4(fatty acid binding protein 4,FABP4)、過氧化物酶體增殖體激活受體γ(peroxisome proliferators activated recepor γ,PPARγ)、乙酰輔酶A羧化酶(acetyl CoA carboxylase,ACC)、脂肪酸合成酶(fatty acid synthase,FAS)是參與脂肪合成與沉積重要調控因子。Western blot分析表明,無菌仔豬脂肪組織中FABP4、PPARγ、ACC、FAS的蛋白表達量都顯著或極顯著低于普通仔豬(圖2-A),提示無菌仔豬在脂肪合成代謝方面顯著弱于普通仔豬。
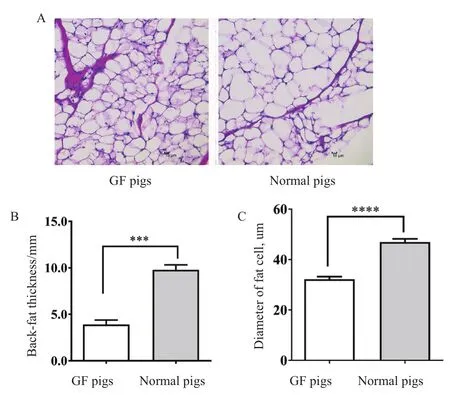
圖1 無菌仔豬和普通仔豬脂肪沉積的差異Fig.1 Difference of fat deposition between GF pigs and normal pigs
脂聯素(adiponection)和瘦素(leptin)是脂肪組織分泌產生的兩種最為重要的細胞因子。試驗結果表明,相比于正常仔豬,無菌豬的脂肪組織中脂聯素和瘦素的含量均不同程度降低(P=0.100,圖2-B;P=0.095,圖2-C)。這個結果一定程度上反應了無菌仔豬脂肪組織生理功能的減弱。
2.2 轉錄組(RNA-seq)測序結果
經RNA-seq分析,在無菌豬和普通豬上平均得到53 472 016條和59 866 864條原始測序序列(Raw reads)。去除低質量的序列后,普通豬上得到干凈序列(Clean reads)59 285 399條(均值,下同),占總序列數的99.02%;無菌豬上得到干凈序列52 961 306條,占總序列數的99.04%。將Clean reads與豬的參考基因組基因序列比對,平均有48 033 697條序列比對豬參考基因組上(比對率>85%)。其中,無菌豬和普通仔豬上的唯一比對序列分別占總比對序列的77.98%和79.43%(表1)。
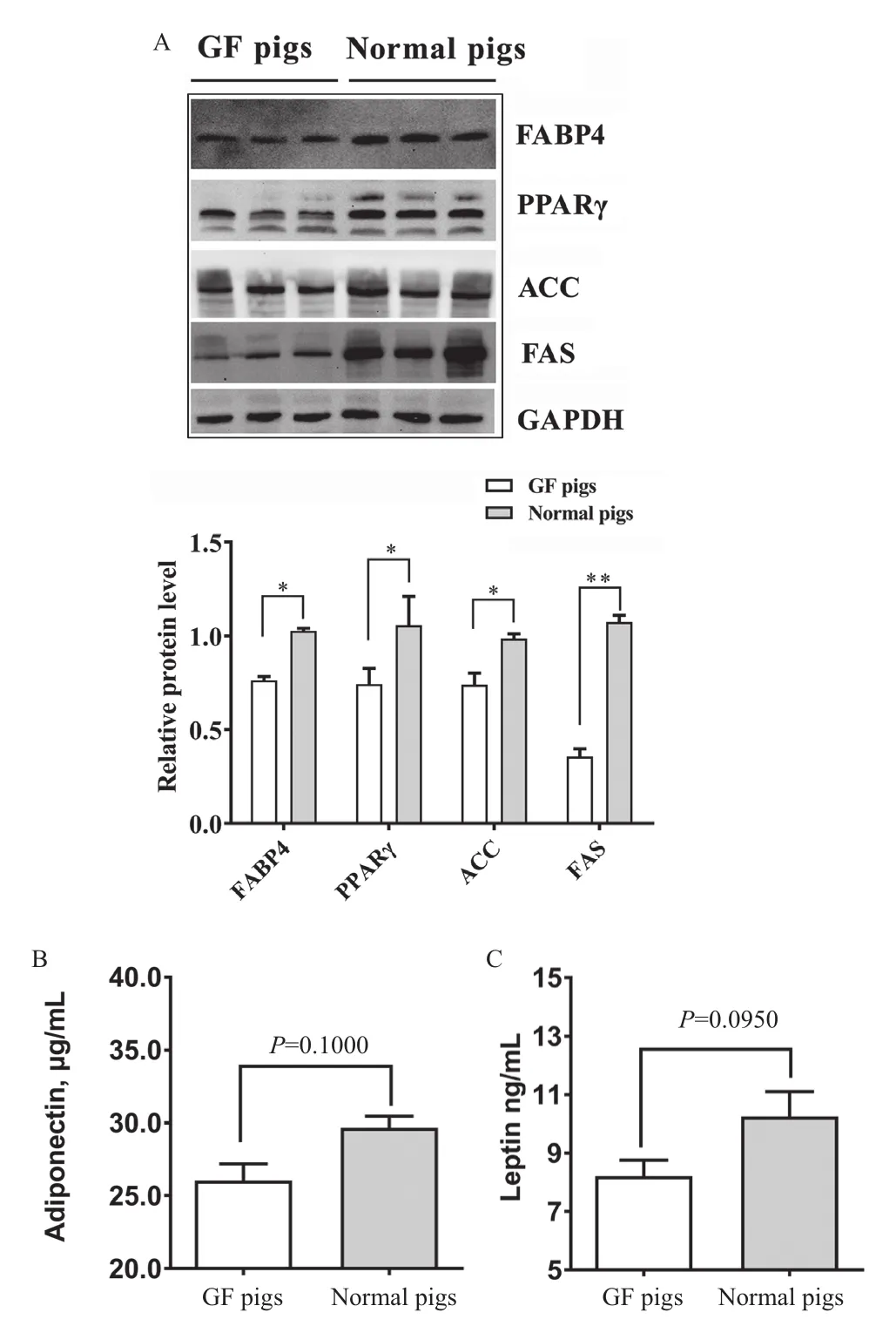
圖2 無菌豬和普通豬的脂肪合成調控分子表達和脂肪細胞因子含量的差異Fig.2 Differences in fat synthesis regulatory factors and adipocytokines between GF pigs and normal pigs
2.3 無菌仔豬和普通仔豬脂肪組織基因表達情況
如圖3所示,6個RNA文庫樣本測序數據對基因覆蓋度好,無兩端傾向性,反應了測序數據完整性及可信度高。對得到的轉錄本信息進行功能注釋,在無菌仔豬和普通仔豬共確認了15 711個已知基因。將所有基因的表達水平進行歸一化處理后,分析并繪制無菌豬和普通豬的基因整體表達水平分布圖和樣本間基因相關性熱圖。結果顯示兩組的全部基因表達量整體差異不明顯,樣本間基因表達相關性好,反應實驗和樣本選擇的可靠,可應用于后續的差異基因篩選與分析(圖4)。

表1 測序數據基因組比對統計Table 1 Statistical list of mapping to genome
圖3 樣本測序基因覆蓋度Fig.3 Coverage of sequencing
2.4 差異表達基因及qRT-PCR驗證
韋恩分析顯示13 747個基因在兩種豬上共表達,764個基因僅在普通仔豬中表達,1 200個基因僅在無菌仔豬中表達(圖5-A)。去除了低表達基因(FPKM<1),然后根據變化倍數(fold change)>2和P<0.001的標準篩選差異表達基因(differently expressed genes,DEGs)。結果顯示,無菌仔豬和普通仔豬的脂肪組織有695個DEGs,其中表達上調基因338個,表達下調基因357個(圖5-B 和5-C)。表2顯示了無菌仔豬脂肪中差異最顯著的10個上調基因和10個下調基因。其中,與脂肪轉運、合成以及代謝相關的基因:LIPG、SucCβ、PIK3C2G、CYP1A1;與炎癥反應相關的基因:PRG4;與營養能量代謝相關的基因:MAPKAPK;與氨基酸合成相關的基因:NRIP3、C4BPA。
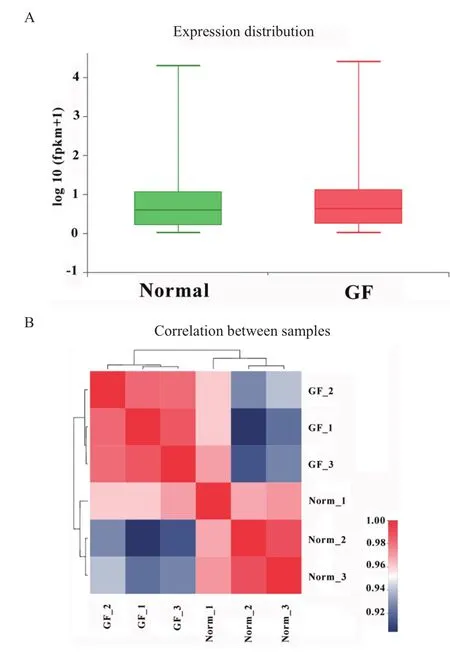
圖4 樣本基因表達分布和相關性Fig.4 Distribution and correlation of genes expression patterns in different samples
通過qRT-PCR方法對測序中得到一些脂肪代謝調控關鍵基因的表達變化進行驗證,PCR結果與轉錄組測序的結果基本一致(圖6)。與普通豬相比,無菌豬體內一些調控脂肪分解代謝的基因如ATGL,UCP3表達顯著上調,此外,負責脂質吸收、轉運的基因如CD36和FATP1的表達水平也被上調,而涉及脂肪合成代謝的調控基因(PPARG和CEBPA)的表達則不同程度下調。
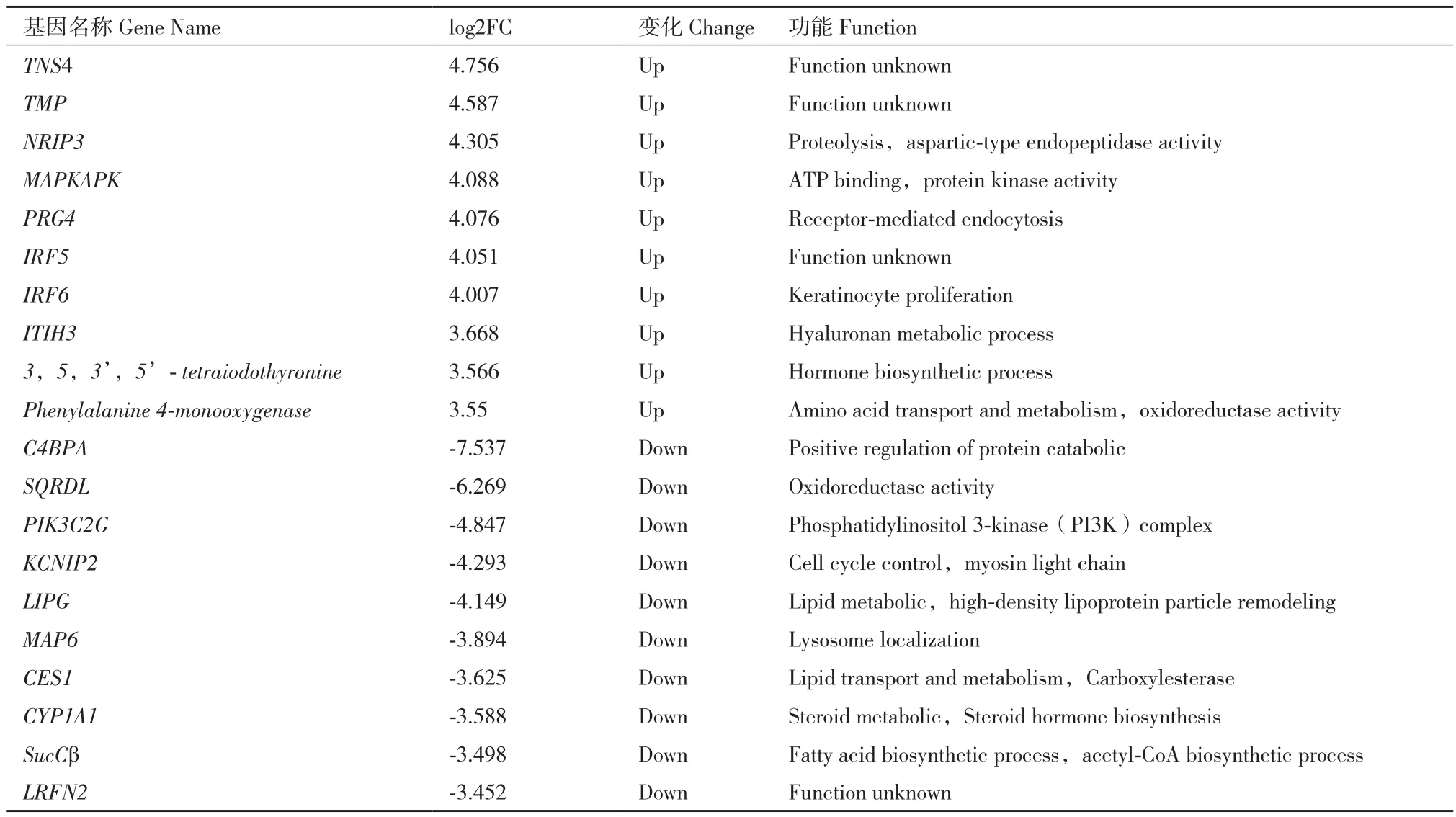
表2 十個表達差異最顯著的上調和下調基因Table 2 Top 10 up or down expressed genes
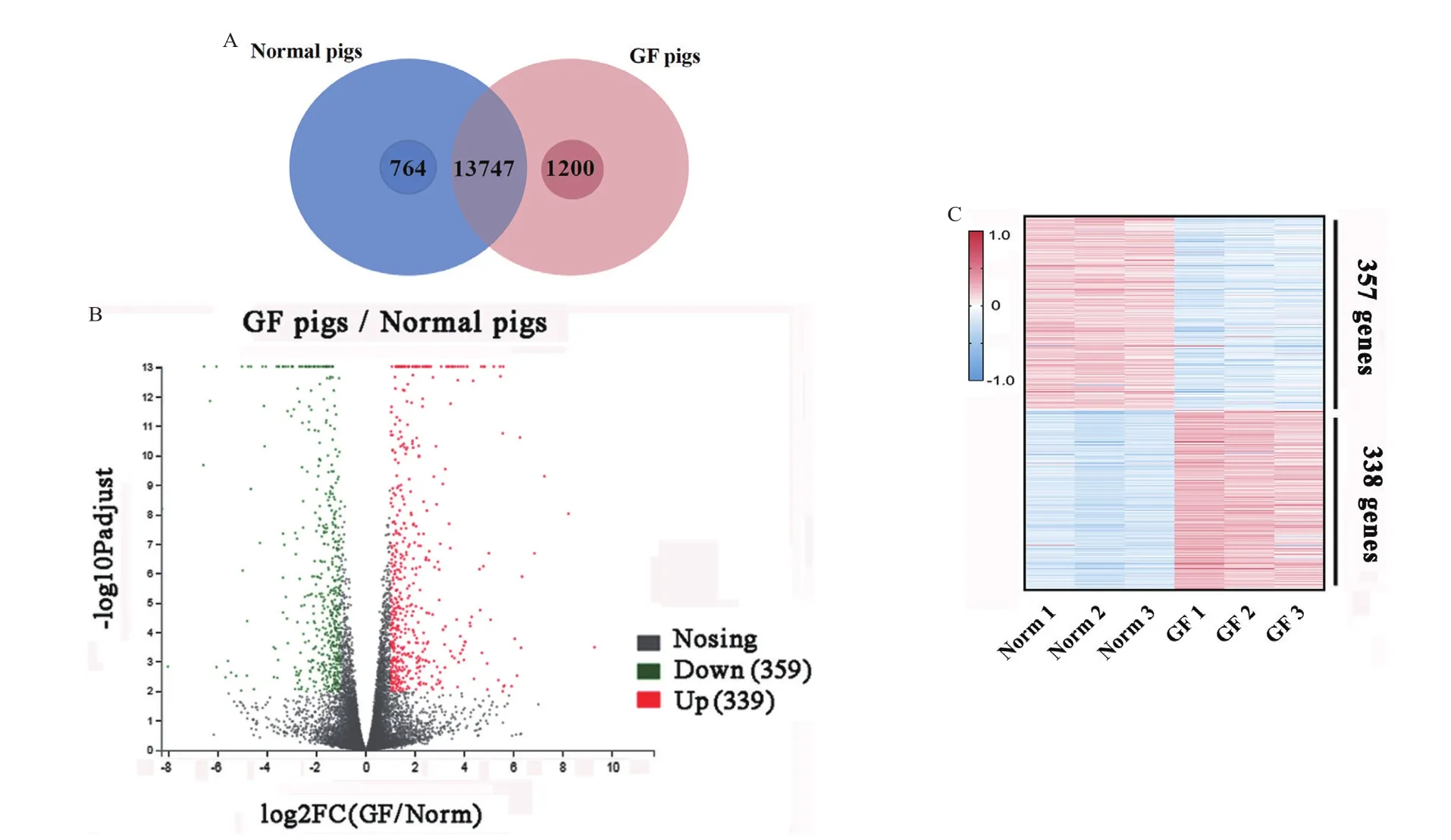
圖5 差異表達基因Fig.5 Differentially expressed genes
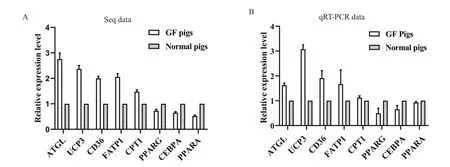
圖6 差異表達基因的qRT-PCR驗證Fig.6 Verification of the differentially expressed genes by qRT-PCR
2.5 差異表達基因的KEGG富集分析
將測序得到的所有差異表達基因進行KEGG通路富集分析,進一步揭示帶菌豬和無菌豬脂肪代謝的差異分子網絡。圖7分別顯示了無菌豬脂肪組織中表達上調和表達下調基因富集的KEGG通路(富集顯著性前20位)。表達上調基因主要富集于脂肪分解、脂質代謝(sphingolipid,glycerolipid,ether lipid等)、PPAR信號通路;而表達下調基因主要與免疫功能、細胞生長、物質代謝(糖代謝、脂類合成、氨基酸合成)和能量穩態有關,主要包括細胞周期、糖酵解、PI3K-Akt信號通路、AMPK信號通路、MAPK信號通路等。這些結果說明缺少腸道菌的存在,仔豬體內的物質分解代謝和能量消耗大于能量蓄積和合成代謝,這些代謝的差異引起脂肪生長和發育遲緩。
3 討論
脂肪是動物體內存儲能量的重要組織,脂質合成與沉積同動物體內營養物質的消化、吸收、代謝和能量周轉有著直接的關系[20]。在家畜中,脂肪組織的生長發育對于動物產品品質(肉、蛋、奶)也有重要的影響。早在2004年,腸道微生物首次被證實對宿主的脂肪組織發育具有重要的調節作用[21]。而隨著研究的深入,研究者們發現腸道微生物與動物的脂肪沉積存在直接的聯系[22]。如Turnbaugh等[23]給體況相近的成年無菌小鼠接種肥胖或瘦的小鼠腸道微生物,飼養14 d后發現接種肥胖小鼠腸道微生物的無菌小鼠體脂含量分別比接種瘦小鼠腸道微生物的無菌鼠和未接種的無菌鼠體脂含量增加了20%和47%。然而目前大多數針對腸道微生物調控宿主脂肪發育的研究都基于嚙齒類動物模型。豬作為一種自然界肥育度最高的動物,是研究腸道微生物調控脂肪發育和脂質代謝的天然有利模型。研究腸道微生物對于豬脂肪組織的影響不僅能補充我們對腸道微生物生理調控功能的認識,也會為下一步通過微生物手段來改善豬肉品質奠定新的理論基礎。本研究中,我們通過直接比較無菌仔豬和普通仔豬,發現無菌豬的體脂沉積量、脂肪細胞尺寸、脂肪細胞分泌功能均弱于普通仔豬,說明了腸道菌群的缺失導致宿主脂肪發育不良。
近年來,基于無菌動物模型、菌群移植技術、宏基因組等多種研究技術,腸道微生物與宿主脂肪組織發育之間的聯系正在被逐漸揭示。腸道微生物影響脂肪組織發育可能涉及多種不同的機制。(1)腸道微生物可以調控胰島素等激素的分泌影響脂肪的生長發育[24]。(2)多數情況下,腸道微生物是通過產生和釋放代謝產物如短鏈脂肪酸、生物胺、細菌素等物質進入循環系統,到達并作用于脂肪細胞和脂肪組織,影響脂肪組織代謝[25]。(3)腸道微生物還能通過“微生物-腸-腦軸”調控大腦的營養感知、激素分泌、神經信號傳導,進而改變宿主的食欲、采食量、生長節律、物質代謝和能量周轉平衡,影響脂肪組織的發育[26-28]。(4)在一些特殊病理或者炎癥情況下,腸道菌群能突破腸道屏障,通過“腸漏”逃逸到脂肪組織,直接刺激和影響脂肪細胞[29]。其中,短鏈脂肪酸是目前研究較多的腸道菌重要代謝產物,除了作為能源底物直接被腸道吸收,它們也可以作為信號分子激活靶器官的G蛋白偶聯受體GPCR41和GPCR43,促進瘦素等激素的分泌,調控宿主的糖脂代謝和脂肪組織胰島素敏感性[30-31]。本研究中,我們在無菌豬的脂肪組織上也觀察到瘦素和脂聯素分泌量的減少和G蛋白偶聯受體家族基因(GPCR)的表達下調,可以推測無菌狀態下,腸道內產生的短鏈脂肪酸大量減少進而導致脂肪組織分泌功能的降低。
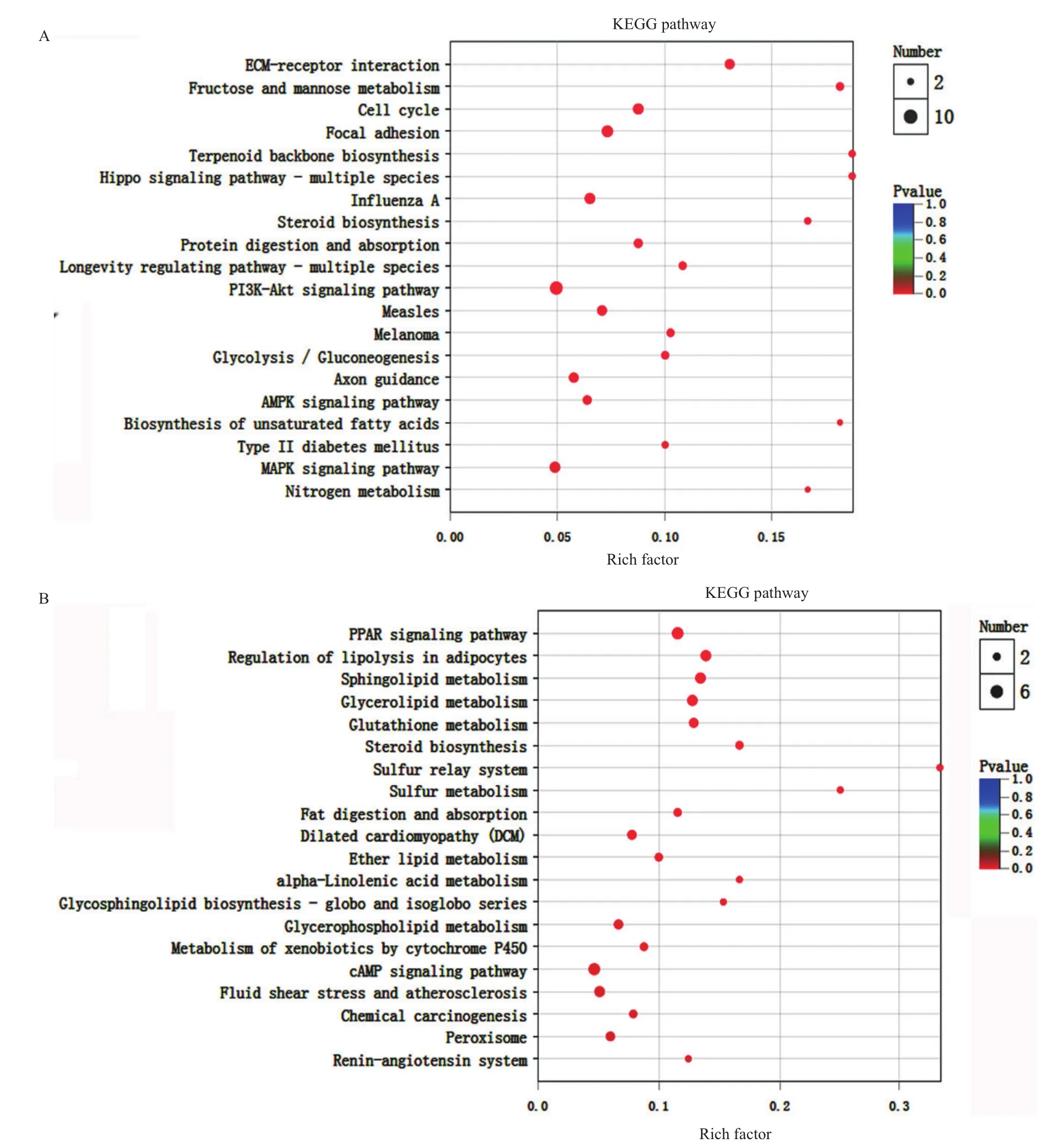
圖7 差異表達基因的KEGG功能富集Fig.7 KEGG enrichment analysis of the differentially expressed genes
伴隨著脂肪組織形態和功能變化的是分子層面的巨大差異。我們通過高通量測序解析了無菌豬和普通豬的脂肪組織基因表達譜的差異,這為進一步探索腸道微生物調節豬脂肪組織發育的分子機制奠定了基礎。本研究結果顯示無菌豬的脂肪中,表達變化最顯著的差異基因主要與免疫、炎癥、脂肪和能量代謝直接相關。如已有文獻報道能催化ATP水解和具有抑制腸上皮細胞屏障功能的MAPK激酶蛋白酶(MAPK-Activated Protein Kinases,MAPKAPK)[32-34]、能在炎癥條件下被急劇上調來穩定細胞外基質的α-胰蛋白酶抑制劑重鏈3(Interα-trypsin inhibitor heavy chain 3,Itih 3)[35-36]以及與氨基酸轉運代謝和氧化還原酶活性正相關的苯丙氨酸4-單加氧酶(phenylalanine 4-monooxygenase)都顯著表達上升[37],而能激活糖原合成酶,促進肝糖原的合成,增加甘油三酯的生成的磷脂酰肌醇3激酶復合物(phosphatidylinositol 4-phosphate 3-kinase C2 domain-containing subunit gamma,PIK3C2G)[38]、有助于高密度脂蛋白/低密度脂蛋白/極低密度蛋白通過內吞進入細胞內產生分解反應并為細胞提供脂質合成必需的脂質前體以及具有一定抗炎癥作用的內皮脂肪酶(endothelial lipase,LIPG)[39]、有著膽固醇水解活性和甘油三酯水解酶活性的羧酯酶1(carboxylesterase 1,CES1)[40-41]以及能抑制 LPS 誘導的炎癥細胞因子腫瘤壞死因子-α和白細胞介素-6表達的細胞色素P450 1A1(cytochrome P450 1A1,CYP1A1)均顯著下調[42]。
在嚙齒類動物類動物模型上,Backhed等[43]的研究發現無菌小鼠的能量利用和消耗高于普通小鼠。同樣地,通過轉錄組分析我們發現無菌豬的能量代謝強于普通仔豬,但脂肪發育上卻弱于普通仔豬。如無菌豬中上調表達的差異基因顯著富集于脂肪分解、糖代謝和能量利用,如脂肪細胞脂解調控信號通路(regulation of lipolysis in adipocytes)和糖異生相關的cAMP信號通路(cAMP signaling pathway)等,而下調表達的基因則更多的與免疫、炎癥、細胞生長和物質合成代謝有關,如具有調節器官發育、組織穩態以及脂代謝功能的關鍵信號通路Hippo 信號通路(hippo signaling pathway)、與細胞生長、發育、葡萄糖穩態相關的PI3K-Akt信號通路以及與不飽和脂肪酸合成相關的不飽和脂肪酸合成信號通 路(biosynthesis of unsaturated fatty acids signaling pathway)。這些研究結果再次強調了正常的腸道菌群對于維持和促進宿主動物的脂肪代謝和脂肪組織發育是非常必要的。
4 結論
本研究基于無菌豬和普通豬脂肪細胞形態和脂肪基因表達譜的比較,證實了腸道微生物對于維持和促進豬的脂肪組織發育具有重要影響。腸道微生物的缺失減少了豬的體脂沉積,削弱了脂肪細胞的生理功能,導致脂肪組織中數百個基因發生了顯著的變化,主要涉及細胞周期、免疫功能、物質(糖、脂、氨基酸等)代謝和能量穩態調控。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
