高溫和強光復合脅迫對鐵皮石斛光系統I和II活性的影響*
2021-06-25 00:59:40黃偉譚順玲孫虎
西部林業科學 2021年3期
關鍵詞:影響
黃偉,譚順玲,孫虎
(1.中國科學院昆明植物研究所 資源植物與生物技術重點實驗室,云南 昆明 650201;2.中國科學院昆明植物研究所 植物醫生研發中心,云南 昆明 650201)
植物利用光能進行光合作用,通過光反應產生的ATP(腺嘌呤核苷三磷酸)和NADPH(還原型煙酰胺腺嘌呤二核苷酸磷酸)在暗反應中被用于固定二氧化碳(CO2)并轉化為合成有機物。光反應中的光合電子傳遞分為線性電子傳遞、圍繞光系統I(PSI)的環式電子傳遞、假環式電子傳遞等[1]。光合電子傳遞的運轉形成跨類囊體膜質子驅動勢,驅動ATP合成酶合成ATP[2]。線性電子傳遞產生的ATP/NADPH比值大約為1.29,暗反應所需的ATP/NADPH比值大概為1.6,所以植物需要其他的替代電子傳遞途徑來補充ATP的合成[3]。環式電子傳遞產生的跨類囊體膜質子梯度不僅補充ATP的合成,并且通過調節光合電子傳遞鏈的氧化還原態保護PSI和光系統II(PSII)免受過剩光能的損傷[4]。非生物脅迫對PSII活性的影響主要集中在單因子脅迫(高溫、低溫、強光、干旱等)[5-8],但是目前關于植物對高溫和強光復合脅迫的光合生理響應還了解較少。
中等程度高溫會對PSII的放氧復合體活性造成損傷[9],進而引起PSII的光抑制[10]。與此同時,高溫導致氣孔關閉,抑制卡爾文循環,增加葉綠體內活性氧自由基的產生[11]。活性氧自由基會抑制D1蛋白的合成,進而影響PSII活性的快速修復[12-13]。所以,高溫脅迫不僅會提高損傷速率,還會降低修復速率[14]。在強光條件下,過剩光能大部分通過非光化學淬滅以無害的形式耗散(熱耗散),避免PSII的修復受到抑制[7,15-16]。熱耗散的激發依賴于跨類囊體膜質子梯度(ΔpH)的建立,而ΔpH的建立又依賴線性電子傳遞和環式電子傳遞[17],光合電子傳遞的運轉受PSI和PSII活性的控制[18-20]。所以,在高溫和強光復合脅迫下,PSII的損傷有可能抑制線性電子傳遞并影響跨類囊體膜質子梯度的建立,從而加劇光抑制的發生。
高溫脅迫也會對PSI的活性造成影響,研究表明42 ℃的中等程度高溫和300 μmol/(m2·s1)光照處理10 h就會對小麥(Triticumaestivum)的PSI活性造成嚴重損傷,且損傷程度要高于PSII[5]。這主要是高溫加劇了類囊體膜的質子滲漏,降低了ΔpH[21],進而造成PSI的電子載體處于過度還原態,引發PSI內活性氧自由基的產生,造成PSI受損。此外,42 ℃的中等程度高溫會加劇波動光強下煙草(Nicotianatabacum)幼葉的PSI損傷程度[22]。但是,中等程度高溫和持續恒定強光對PSI活性的影響還缺乏研究。
鐵皮石斛(DendrobiumofficinaleKimura et Migo)是一種有重要藥用價值的蘭科(Orchidaceae)植物,在亞熱帶地區被廣泛種植。但是,目前關于逆境脅迫對鐵皮石斛光合機構及光合電子傳遞影響的研究還十分欠缺。由于高溫和強光復合脅迫在鐵皮石斛種植地區的夏季經常出現,因此本研究將以鐵皮石斛“昆植1號”品種為研究對象,研究高溫和強光復合脅迫對其PSI和PSII的影響及其光合生理機制,為鐵皮石斛的種植策略提供參考。
1 材料與方法
1.1 研究材料
供試材料為種植在中國科學院昆明植物研究所溫室內的鐵皮石斛品種“昆植1號”。鐵皮石斛“昆植1號”植株被栽培在直徑為20 cm的透水塑料盆中,以樹皮為基質。生長季晝夜溫度為20/30 ℃,空氣濕度維持在60%左右,光照條件為全光照的40%。
1.2 實驗設計
將鐵皮石斛植株在常溫下暗適應30 min后,放入42 ℃溫室中繼續暗適應5 min,然后利用Dual-PAM 100雙通道葉綠素熒光儀(德國Walz公司)測定葉綠素熒光快速動力學曲線(300 ms)和Pm值,分析處理前葉片的PSII和PSI活性。隨后暗適應5 min,然后測定強光〔1 809 μmol/(m2· s)〕處理過程中PSI和PSII參數的變化。處理60 min后,再暗適應30 min測定葉綠素熒光快速動力學曲線和Pm值,分析處理后葉片的PSII和PSI活性。通過比較處理前后PSII和PSI活性的差異,判斷高溫強光處理對PSII和PSI活性的損傷程度。
1.3 指標測定
PSI和PSII參數的測定都利用Dual-PAM 100雙通道葉綠素熒光儀(德國Walz公司)完成。葉綠素熒光快速動力學曲線的測定使用20 000 μmol/(m2· s)的強光脈沖(300 ms),葉綠素相對熒光強度的快速動力學曲線利用最大熒光強度進行標準化。PSII的參數計算如下[23]:
Y(II)=(Fm′ -Fs)/Fm′;NPQ=(Fm-Fm′)/Fm′;Y(NO)=Fs/Fm。
式中:Fm代表暗適應后的最大葉綠素熒光強度;Fm′代表光適應后的最大葉綠素熒光強度;Fs代表光適應后的穩態熒光強度;Y(II)表示PSII的有效光化學量子產額;NPQ表示PSII的非光化學淬滅能力;Y(NO)表示PSII的非調節性熱耗散量子產額。PSI的參數計算如下[22]:
Y(I)=(Pm′-P)/Pm;Y(ND)=P/Pm;Y(NA)=(Pm-Pm′)/Pm。
式中:Y(I)代表光PSI的光化學量子產額,Y(ND)代表PSI反應中心P700的氧化態比例,Y(NA)代表PSI反應中心P700的還原態比例。光合電子傳遞速率的計算如下:
ETRI = PPFD×Y(I)×0.84×0.5;ETRII = PPFD×Y(II)×0.84×0.5。
式中:ETRI和ETRII分別代表通過PSI和PSII的電子傳遞速率,ETRI-ETRII代表環式電子傳遞速率[24],PPFD代表光合有效輻射強度。
1.4 數據處理
本研究的每組實驗測定包括5個重復,數據為平均值±標準誤。采用SPSS 16.0軟件對數據進行統計分析。處理前后的數據比較采用獨立樣本T檢驗進行。
2 結果與分析
2.1 高溫強光脅迫對葉綠素快速動力學曲線的影響
高溫強光脅迫對鐵皮石斛葉綠素影響見圖1。
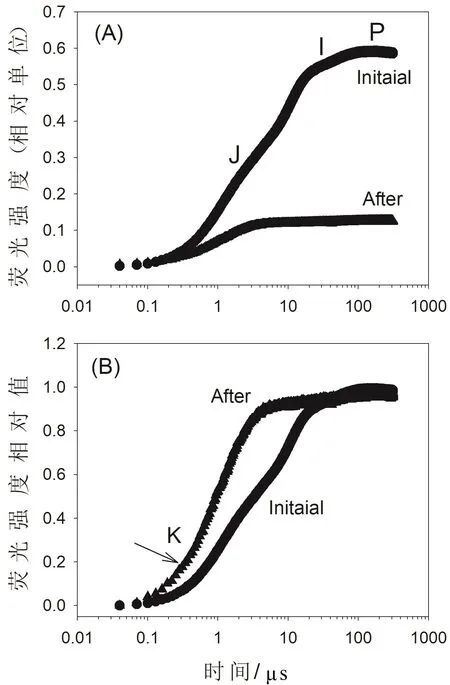
圖1 高溫強光處理對鐵皮石斛葉綠素熒光快速動力學曲線的影響
在42 ℃的中等高溫脅迫下對鐵皮石斛葉片進行強光〔1 809 μmol/(m2· s〕處理60 min后,測定其葉綠素熒光快速動力學曲線(OJIP曲線),發現J點、I點和P點的葉綠素熒光強度都發生大幅度下降(圖1A),表明PSII活性受到了嚴重損傷。此外,在300 μs時的相對葉綠素熒光強度(K點)發生了顯著上升(圖1B),表明PSII的電子供體即放氧復合體活性受到了明顯的損傷。
2.2 高溫強光脅迫對PSII光量子產額的影響
高溫強光脅迫對鐵皮石斛PSII光量子產額的影響見圖2。
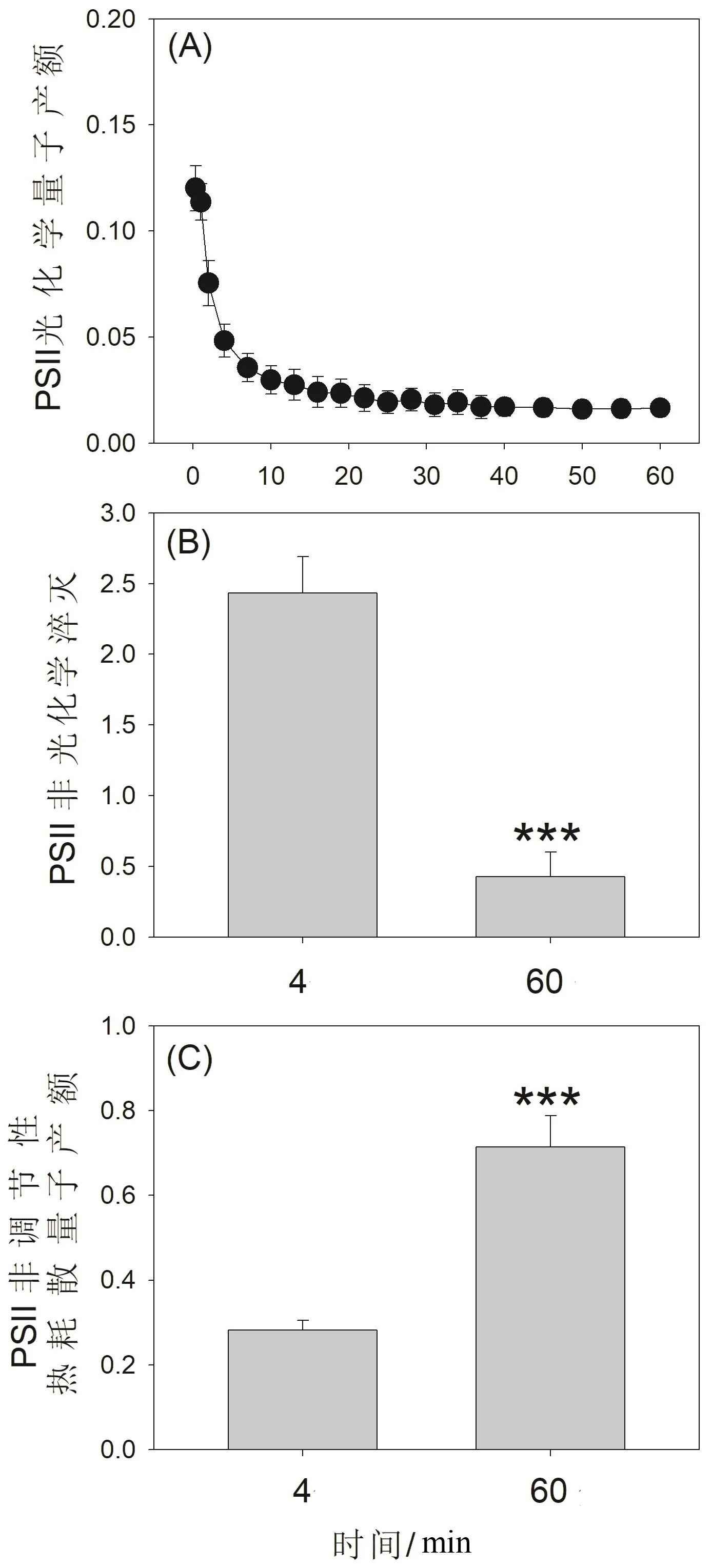
圖2 高溫強光處理對鐵皮石斛光系統II量子產額的影響
在暗適應葉片突然曝光的前1 min內,鐵皮石斛的PSII光化學量子產額[Y(II)]維持在很高水平(圖2A),這主要和水-水循環介導的PSII電子傳遞相關。在高溫和強光處理過程中,Y(II)在前6 min內大幅度下降(圖2A),說明鐵皮石斛的卡爾文循環受到了高溫脅迫的強烈抑制,導致線性電子傳遞產生的ATP和NADPH不能被及時消耗,NADP+/NADPH的比例下降,影響了電子從PSII經過PSI再傳遞給NADP+的過程。在高溫和強光處理60 min后,鐵皮石斛的NPQ從2.4下降到0.43,下降幅度達到了82%(圖2B)。NPQ的激發不僅依賴跨類囊體膜質子梯度,同時還受PSII復合體核心活性的影響,前者和后者對NPQ的貢獻分別大概為60%和40%。所以,鐵皮石斛經高溫和強光處理后NPQ的大幅度下降不僅和跨類囊體膜質子梯度的不足有關,同時還涉及PSII反應中心的受損。當NPQ能力不足時,PSII吸收的大部分光能就變成了過剩光能,引起Y(NO)的大幅度上升(圖2C)。Y(NO)的大幅度上升會引起活性氧自由基在PSII復合體處大量產生,進而影響D1蛋白的合成和PSII的修復過程。
2.3 高溫強光脅迫對PSI量子產額的影響
高溫強光脅迫對鐵皮石斛PSI光量子產額的影響見圖3。
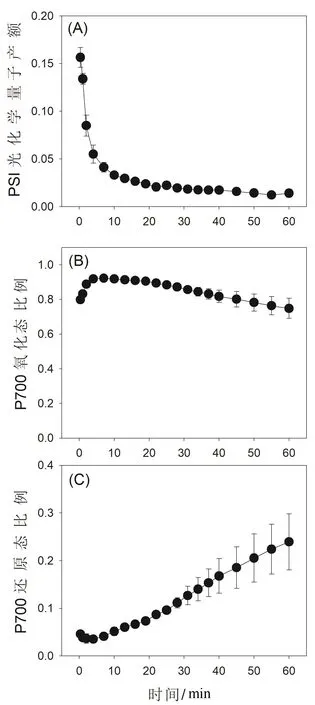
圖3 高溫強光處理對鐵皮石斛光系統I量子產額的影響
與Y(II)類似,在暗適應葉片突然曝光的前1 min內,鐵皮石斛的PSI光化學量子產額[Y(I)]維持在很高水平(圖3A),這主要和水-水循環及環式電子傳遞的高度激發有關。在高溫和強光處理過程中,Y(I)在前6 min內大幅度下降,隨后緩慢下降(圖3A)。PSI的反應中心在暗適應突然曝光后的20 s就高度氧化(圖3B),說明水-水循環在高溫處理的初期耗散了積累在PSI的過剩活化電子,避免其出現過度還原態(圖3C)。但是,在隨后的高溫強光處理過程中,PSI的氧化態比例逐漸下降(圖3B),過度還原態比例逐漸上升(圖3C),這主要和PSI的逐漸受損相關。
2.4 高溫強光脅迫對光合電子傳遞的影響
高溫強光脅迫對鐵皮石斛光合電子傳遞的影響見圖4。
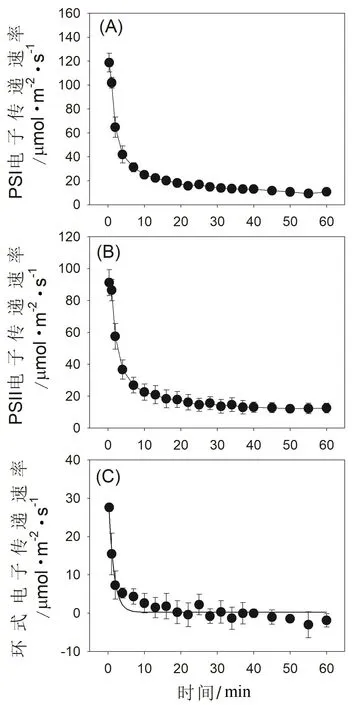
圖4 高溫強光脅迫對鐵皮石斛光合電子傳遞速率的影響
在暗適應葉片突然曝光的前1 min內,PSI電子傳遞速率(ETRI)維持在很高水平(圖4A),這主要是因為水-水循環和環式電子傳遞都高度激發。在隨后的處理過程中,水-水循環介導的PSII電子傳遞大幅度下降(圖4B),并且環式電子傳遞的激發受到抑制(圖4C),導致ETRI快速下降。值得注意的是,在高溫強光處理5 min以后,環式電子傳遞的活性基本喪失(圖4C),這會嚴重影響跨類囊體膜質子梯度的構建,進而抑制NPQ的激發。
2.5 高溫強光脅迫對PSI和PSII活性的影響
經過60 min高溫和強光處理后,最大葉綠素熒光強度(Fm)下降了78%,表明PSII活性受到嚴重損傷。與此同時,最大可氧化的P700值(Pm)下降了31%,說明PSI活性也出現了中等程度損傷。相比較PSII活性而言,PSI活性的受損程度較低,這說明PSII是高溫強光脅迫對鐵皮石斛造成光損傷的主要靶點。
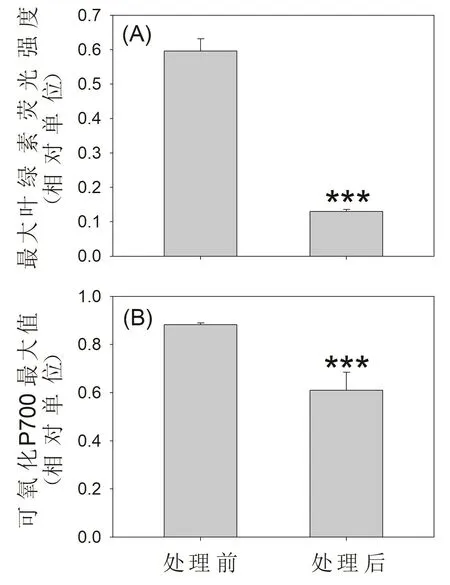
圖5 高溫強光脅迫對鐵皮石斛光系統I和II活性的影響
3 討論與結論
3.1 討論
自然條件下,高溫和強光脅迫是植物在夏季經常遭遇的非生物逆境脅迫,但這種復合脅迫對植物葉片PSI和PSII的影響還有待揭示。有研究表明,42 ℃和中等光強復合脅迫對小麥PSI活性的影響大于其對PSII活性的影響。本研究發現,在42 ℃下強光脅迫處理60 min后鐵皮石斛的最大葉綠素熒光值(Fm)下降了78%,表明其PSII活性受到嚴重損傷[20]。與此同時,其最大可氧化的P700值(Pm)下降了31%,表明PSI活性也有顯著的下降[22,25-26]。所以,高溫和強光脅迫對鐵皮石斛PSII活性的影響大于其對PSI活性的影響。此外,通過PSI和PSII的電子傳遞速率分別下降91%和86%,說明PSI和PSII的損傷對整個光合電子傳遞鏈的功能造成了嚴重影響。
PSII的損傷程度由損傷速率和修復速率共同決定,當損傷速率大于修復速率時,PSII活性發生損傷[27]。反之,當修復速率大于損傷速率時,PSII活性得到修復[28]。在自然條件下,強光導致的PSII損傷會在弱光條件下修復[29]。PSII的損傷速率主要和光照強度相關,但修復速率會受到其他環境因素的影響,例如鹽脅迫、溫度脅迫等[10]。本研究發現,高溫和強光復合脅迫處理對鐵皮石斛的放氧復合體活性造成顯著性損傷,這和PSII損傷的“兩步模型”相吻合。在“兩步模型”中,放氧復合體是PSII損傷的最初靶點,放氧復合體的受損隨后引起PSII的損傷[30]。在逆境條件下,葉片的光能利用效率下降,導致活性氧自由基的含量上升。活性氧自由基會抑制PSII復合體關鍵亞基D1蛋白的重新合成,進而影響D1蛋白和受損PSII復合體的組裝,限制PSII活性的修復。所以,環境脅迫對PSII修復速率的影響主要和活性氧自由基的產生密切相關[10]。當暗適應葉片在42 ℃下突然曝光后,其PSII電子傳遞速率(ETR II)先保持在較高水平,然后急速下降,這主要和水-水循環電子傳遞的短暫激發相關,此時水-水循環電子傳遞的運轉避免了PSI處于過度還原態,也消耗了PSII的部分過剩光能。但是,在隨后的高溫強光處理過程中,ETR II逐漸下降并維持在較低的水平,說明鐵皮石斛的暗反應受到限制。高溫容易引起卡爾文循環的受限,導致NADPH的消耗速率變慢,引起NADP+的缺失,進而造成電子從PSI到NADP+的傳遞受阻,最終導致ETRII受限,引發活性氧自由基的大量產生,從而抑制PSII活性的修復。此外,非光化學淬滅(NPQ)作為PSII耗散過剩光能的主要保護機制,在高溫強光脅迫處理60 min后大幅度下降,這也加劇了PSII內部活性氧自由基的產生。所以,在42 ℃和強光復合脅迫條件下,較高的PSII損傷速率和較低的PSII修復速率協同作用,導致鐵皮石斛的PSII活性嚴重受損。
PSII活性雖然容易受損,但是在合適條件下只需要幾個小時就能夠基本修復[31]。而PSI活性則不同,一旦受損就需要一周甚至更長時間才能完全修復[18,32]。PSI的損傷主要是由于其反應中心附近的電子載體處于過度還原態,造成PSI復合體內部產生大量的活性氧自由基,進而引發氧化損傷[33]。在正常生長條件下,葉片產生較高的跨類囊體膜質子梯度(ΔpH),在Cyt b6f復合體處調節PSII到PSI的電子傳遞,保證PSI的電子載體在強光條件下都處于高度氧化態,進而避免過度還原引起的光損傷[34-35]。本研究發現,鐵皮石斛的PSI電子載體在高溫強光脅迫處理的初期并沒有發生過度還原,但是在處理后期PSI電子載體出現過度還原并伴隨著光損傷。ΔpH在調控PSI氧化還原態過程中發揮重要作用,例如在擬南芥(Arabidopsisthaliana)環式電子傳遞突變體pgr5中,強光條件下ΔpH的建立不足導致其PSI發生嚴重損傷,甚至造成植株死亡[36-39]。所以,環式電子傳遞是植物在強光條件下建立充分的ΔpH并保護PSI活性的重要替代電子傳遞途徑[1,40]。本研究還發現,在高溫強光脅迫處理的初期,鐵皮石斛的環式電子傳遞被高度激發。但是在處理后期,環式電子傳遞的激發程度很低,這就導致ΔpH的建立不足。同時,由于NPQ的激發也依賴ΔpH的建立,因此高溫強光處理后NPQ的下降也證實了ΔpH的建立受限。所以,長期的高溫強光脅迫會導致環式電子傳遞失活,引起ΔpH的建立不足,進而引起PSI活性受損。
3.2 結論
高溫脅迫對光系統II活性及光量子產額的影響是導致其光合作用機構受損的重要方面[41-44]。本研究發現,在高溫強光復合脅迫處理60 min后,鐵皮石斛的PSI和PSII活性都顯著下降,且PSII的下降幅度更大,表明PSII活性的受損是鐵皮石斛難以適應長期高溫強光復合脅迫的重要原因。提高光能利用效率是維持植物生長和提高產量的基礎,而負責光反應的PSI和PSII活性是決定光能利用效率的重要因素。鑒于高溫強光脅迫會對鐵皮石斛的PSI和PSII活性造成嚴重損傷,在鐵皮石斛種植條件的選擇和控制上,應盡量避免高溫和強光復合脅迫。本研究的設計思路和實驗結果對于其他農作物和經濟作物種植策略的選擇同樣具有參考和借鑒意義。
猜你喜歡
英語文摘(2022年4期)2022-06-05 07:45:18
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
家庭影院技術(2020年10期)2020-12-14 07:54:18
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
知識經濟·中國直銷(2016年3期)2016-02-27 16:15:49
現代檢驗醫學雜志(2014年6期)2014-02-02 03:02:04
