廣西家蠶核型多角體病毒gp64基因遺傳多態(tài)性及進化分析
2021-06-30 02:53:44陳慧珠龍羽燕屈達才徐開遵朱方容陳朝蓉梁湘
南方農業(yè)學報 2021年2期
陳慧珠 龍羽燕 屈達才 徐開遵 朱方容 陳朝蓉 梁湘


摘要:【目的】了解廣西家蠶核型多角體病毒(Bombyx mori nucleopolyhedrovirus,BmNPV)gp64基因的遺傳多態(tài)性及進化特點,掌握BmNPV在廣西蠶區(qū)的流行和傳播情況,揭示BmNPV種群維持遺傳多樣性的模式與機制。【方法】對20株廣西BmNPV毒株的gp64基因進行測序分析,根據gp64基因構建遺傳進化樹及繪制毒株流行分布圖,并比對不同毒株的致病力。【結果】廣西BmNPV毒株gp64基因開放閱讀框(ORF)長度存在3種情況(1590、1593和1599 bp),分別編碼含529、530和532個氨基酸殘基的GP64蛋白。20株廣西BmNPV毒株與標準參考T3株的gp64基因核苷酸序列同源性在97.6%~99.2%,其推導氨基酸序列同源性在96.4%~99.6%。在廣西BmNPV毒株gp64基因近N端分別出現GCG缺失和GCGCCG/GTGCCG插入突變,發(fā)生核苷酸替換突變的位點數目在13~28個,但大部分為同義替換,對編碼蛋白的三聚體空間構象無明顯影響。廣西BmNPV毒株gp64基因編碼蛋白的N-糖基化位點為3~4個;除GXZS株外,所有毒株的O-糖基化位點均為2個,且預測位點一致。基于gp64基因構建的遺傳進化樹顯示,幾乎所有的廣西BmNPV毒株聚類于Clade I分群,其又被分為2個主要亞群(Sub-clade I和Sub-clade II);而幾乎所有的國外參考毒株聚類于Clade II分群。廣西蠶區(qū)的BmNPV流行分布呈集中性與分散性并存;GXUA株對四齡和五齡起蠶的半數致死量(LD50)分別為3.3和3.1,而GXZZ株對四齡和五齡起蠶的LD50分別為5.5和5.3,說明GP64蛋白糖基化位點較少的BmNPV毒株表現出較弱的致病力。【結論】廣西BmNPV毒株gp64基因在進化過程中其信號肽區(qū)出現明顯變異,發(fā)生同義突變的頻率較高,形成較獨立的進化分群,毒株間的致病力差異可能與GP64蛋白糖基化位點不同有關,說明廣西BmNPV毒株具有不同的基因型和表型,在一定程度上維持了BmNPV野生群體的遺傳多樣性。
關鍵詞: 家蠶核型多角體病毒;gp64基因;遺傳多態(tài)性;進化分析;致病力
中圖分類號: S884.51? ? ? ? ? ? ? ? ? ? ? ?文獻標志碼: A 文章編號:2095-1191(2021)02-0448-09
Abstract:【Objective】This research was aimed to understand the genetic polymorphism and evolving characteristic of the gp64 genes derived from Bombyx mori nucleopolyhedrovirus(BmNPV) in Guangxi, study the condition of Bm-NPV epidemic and spread in Guangxi sericultural regions, andreveal the patterns and mechanisms of maintaining genetic diversity in BmNPV population. 【Method】The gp64 gene of 20 Guangxi BmNPV strains were sequenced and analyzed. The phylogenetic tree was constructed according to gp64 gene and the epidemic distribution map was drawn. The pathogenicity of different strains was also compared. 【Result】The open reading frame(ORF) length of gp64 genes of these Guangxi BmNPV strains showed three conditions(1590,1593 and 1599 bp), which encoded GP64 proteins of 529, 530 and 532 amino acid residues, respectively. The homologies of nucleotide sequence in gp64 genes of 20 Guangxi BmNPV strains was 97.6%-99.2% and the homologies of deduced amino acid sequence was 96.4%-99.6% compared with standard reference T3 strain. The mutations of GCG deletion and GC/TGCCG insertion were found near the N terminal of gp64 gene of Guangxi BmNPV strains. The site mutation numbers of nucleotide substitution were between 13 to 28, but most of them were synonymous substitutions which had little effect on the trimeric spatial conformation of the coded protein. There were three to four N-glycosylation sites in decoded amino acids of gp64 genes of Guangxi BmNPV strains. There were two O-glycosylation sites in all strains except for GXZSand the predicted sites were all the same. Almost all the Guangxi BmNPV strains were clustered as CladeⅠwhich was further separated into two major groups(Sub-clade I and Sub-clade II) in the phylogenetic tree constructed based on gp64 gene, while almost all the reference strains from abroad were clustered as Clade Ⅱ. The BmNPV epidemic distribution showed coexistence of concentration and scatteration. The median lethal dose(LD50) of GXUA strains for 4thand 5th instar silkworm were 3.3 and 3.1, while LD50 of GXUA strains for 4thand 5th instar silkworm were 5.5 and 5.3. It indicated that BmNPV strain with less glycosylation site in GP64 protein glycosylation site showed weak pathogenicity. 【Conclusion】A major mutation in signal peptide of gp64 genes of Guangxi BmNPV strains occurs,? frequency of synonymous mutation is high and a relatively independent cluster is formed during the evolution. The diverse pathogenicity of BmNPV strains may be related to the differentiation of GP64 protein glycosyla-tion sites. It shows that there are diverse genotypes and phenotypes among the Guangxi BmNPV strains. These diversities sustain the genetic diversity in BmNPV wild population to a certain extent.
Key words: Bombyx mori nucleopolyhedrovirus(BmNPV); gp64 gene; genetic polymorphism; evolution analysis; pathogenicity
Foundation item: Guangxi Natural Science Foundation(2017GXNSFBA198158)
0 引言
【研究意義】家蠶血液型膿病是繭絲生產上最常見且危害最嚴重的一種病毒性傳染病(王霞等,2020),為農業(yè)農村部公布的三類動物疫病(中華人民共和國農業(yè)部公告 第1125號)。家蠶血液型膿病傳染性強、病死率高,目前尚無有效的治療方法,生產上主要依靠化學消毒劑降低環(huán)境中的病原含量進行防控,但消毒劑的性狀及穩(wěn)定性通常會影響其消毒效果,因此一旦暴發(fā)流行家蠶血液型膿病即對桑蠶生產造成巨大經濟損失(魯興萌,2012)。家蠶血液型膿病由家蠶核型多角體病毒(Bombyx mori nucleopolyhedrovirus,BmNPV)感染引起,BmNPV組裝形成的多角體(Polyhedral inclusion body,PIB)蛋白晶體結構耐腐爛、干燥、紫外線照射和極端氣候等不良環(huán)境因素,病毒粒子能在多角體的保護下長期保持感染活力(陳朝蓉等,2017)。病毒多角體在自然環(huán)境中富集,極易對桑葉和蠶作環(huán)境造成污染,尤其在交通、貿易等人為因素及風雨、掠食動物等自然因素的作用下,還能導致病毒遠距離傳播(Fuxa,2004),極大制約了蠶桑產業(yè)的可持續(xù)發(fā)展。此外,當前國內的蠶區(qū)呈現建設區(qū)域化,生產規(guī)模集約化,生產季節(jié)集中,蠶種流通及蠶繭貿易頻繁,為BmNPV的傳播和流行創(chuàng)造了有利條件。gp64基因是BmNPV的重要結構蛋白基因,在病毒感染早期和晚期均能轉錄表達(夏定國等,2007;陳朝蓉等,2017),因此,加強對BmNPV流行毒株gp64基因進行遺傳多態(tài)性及進化分析,不僅能為其分子致病機理的研究提供參考,還有利于揭示同一宿主桿狀病毒種群致病力差異的分子機理。【前人研究進展】BmNPV是桿狀病毒科(Baculoviridae)α桿狀病毒屬(α-NPV)的重要成員,是除苜蓿銀紋夜蛾核型多角體病毒(Auto-grapha californica multiple nucleopolyhedrovirus,AcMNPV)外,研究昆蟲桿狀病毒的最常用模型(柳林,2018)。囊膜糖蛋白(GP64)是桿狀病毒出芽型(BV)病毒粒子特有且占比較高的囊膜成分,位于病毒粒子一端,以同源三聚體的形式構成表面嵌突。GP64蛋白是桿狀病毒從宿主細胞質膜出芽并最終形成成熟病毒粒子及病毒在組織細胞間傳播必不可少的要素(Monsma et al.,1996),其介導病毒與細胞受體的識別和吸附,以及低pH條件下病毒囊膜與細胞內吞體膜的融合(Zhou and Blissard,2008)。GP64蛋白還參與決定桿狀病毒的宿主域。Katou等(2006)研究發(fā)現,當BmNPV的gp64基因被AcMNPV的gp64基因替代后,即能順利進入非受納細胞Sf9的細胞核,并產生具有感染活性的子代病毒。此外,GP64蛋白與病毒的致病性密切相關,是病毒重要的毒力因子。AcMNPV的GP64蛋白位于胞外區(qū)靠近跨膜域,有4個氨基酸殘基對病毒的出芽和感染力起重要作用(Li and Blissard,2009),而將GP64蛋白的合成時間延遲能有效降低AcMNPV經口感染棉鈴蟲的毒力(Washburn et al.,2003)。因此,以gp64基因為靶標研發(fā)BmNPV防治藥物具有極大潛能。已有研究證實,以gp64基因為RNA干擾靶標,能有效抑制BmNPV在細胞中的基因表達和復制增殖,而降低家蠶死亡率(夏定國等,2007;Jiang et al.,2013;Zheng et al.,2019)。Li等(2011)研究表明,以表面重組表達BmNPV GP64蛋白的枯草桿菌芽孢免疫家蠶,家蠶機體能對BmNPV入侵產生一定抵抗力。Kato等(2012)通過構建展示表達人腎素受體的重組BmNPV,并證實重組BmNPV對腎素的結合具有親和性,可用于分析受體和配體間的相互作用。Deo等(2014)利用BmNPV構建展示表達腫瘤靶標單鏈可變片段的病毒樣顆粒,為腫瘤的特異性免疫治療提供了新思路。【本研究切入點】宿主昆蟲對病原基因型的選擇是野外桿狀病毒種群保持遺傳多樣性的一種機制(Hitchman et al.,2007)。家蠶在生長和繁殖過程中受人工干預的程度遠高于自然界其他昆蟲種類,且抗家蠶血液型膿病品種的選育和推廣養(yǎng)殖、凈化蠶作環(huán)境病原使用的消毒劑及小蠶共育+大蠶分戶的飼養(yǎng)模式等均會對BmNPV的進化和傳播產生影響,也意味著BmNPV種群所面對的選擇壓力較其他桿狀病毒種群更大,而有可能導致更快的進化速度及衍生特殊的進化模式。GP64蛋白與BmNPV的致病性密切相關,但至今鮮見廣西家蠶核型多角體病毒gp64基因遺傳多態(tài)性及進化分析的研究報道。【擬解決的關鍵問題】對20株廣西BmNPV毒株的gp64基因進行測序分析,根據gp64基因構建遺傳進化樹及繪制毒株流行分布圖,并比對不同毒株的致病力,了解gp64基因遺傳多態(tài)性及其進化特點,旨在掌握BmNPV在廣西蠶區(qū)的流行和傳播情況,以揭示BmNPV種群維持遺傳多樣性的模式與機制。
1 材料與方法
1. 1 試驗材料
供試家蠶品種為兩廣二號的四齡和五齡起蠶。20株BmNPV毒株中有18株是從廣西不同桑蠶產區(qū)采集獲得,采集地點及gp64基因GenBank登錄號分別為GXCW(梧州蒼梧,MT165666)、GXHJ(河池環(huán)江,MT165668)、GXJX(百色靖西,MT165670)、GXLeY(百色樂業(yè),MT165671)、GXLuoC2(河池羅城,MT165672)、GXMS2(梧州蒙山,MT165673)、GXNN(南寧,MT165674)、GXTY(百色田陽,MT165675)、GXWX(來賓武宣,MT165677)、GXXD1(欽州小董,MT165678)、GXYZ2(河池宜州,MT165682)、GXYF(桂林永福,MT165680)、GXYJ(百色右江,MT165681)、GXZP(賀州昭平,MT165683)、GXZS(賀州鐘山,MT165684)、GXXZ(來賓象州,MT165679)、GXHP(北海合浦,MT165669)和GXFM(玉林福綿,MT165 667);GXZZ(MT165685)和GXUA(MT165676)株則分別由廣西蠶業(yè)技術推廣總站和亞熱帶農業(yè)生物資源保護與利用國家重點實驗室保存提供。
1. 2 BmNPV多角體提純及鏡檢
將采集的血液型膿病病蠶樣本分別置于密封瓶子中常溫腐熟,加入超純水稀釋后,以3層紗布過濾去除不腐爛的蠶體組織。濾液進行400和4000 r/min各10 min的反復差速離心,待多角體沉淀呈灰白色時以純水重懸;然后將病毒多角體懸液置于20%~60%連續(xù)密度梯度的蔗糖溶液上,4000 r/min離心15 min,收集含多角體條帶的蔗糖溶液再離心,重懸多角體沉淀后加入等量丙酮溶液,離心收集沉淀以除去附著于多角體表面的脂類雜質。多角體沉淀經37 ℃烘干成粉狀,以備掃描電鏡觀察使用。
1. 3 引物設計與合成
根據BmNPV標準參考株(日本T3株)基因序列信息,采用Primer 5.0設計gp64基因的特異性引物,gp64-F:5'-GCCAAATAGCGGTCGGGTAT-3'(101450~101469 nt),gp64-R:5'-CGCTTGTGGTAT GAGAAACGAAC-3'(99679~99701 nt),預計擴增片段長度1791 bp,包含gp64基因的完整開放閱讀框(ORF)。特異性擴增引物委托深圳華大基因股份有限公司合成。
1. 4 gp64基因克隆及測序
取BmNPV多角體懸液,加入堿性溶液(0.20 mol/L Na2CO3+0.32 mol/L NaCl)進行裂解,以釋放內部的病毒粒子,10000 r/min離心1 min后取上清液,用1 mol/L Tris-HCl調整pH至7.0。加入10% SDS和蛋白酶K,55 ℃作用1 h,使用等體積的苯酚—氯仿—異戊醇混合液(25∶24∶1)抽提病毒DNA。PCR反應體系及擴增程序參照陳朝蓉等(2017)的方法,以58 ℃為最佳退火溫度。將清晰、無非特異性擴增且與預期相符的目的條帶切下,使用DNA純化回收試劑盒進行回收純化。將目的基因連接至pMD18-T載體,陽性克隆送至深圳華大基因股份有限公司測序,并使用MegAlign和SeqMan進行序列拼接。
1. 5 遺傳多態(tài)性及進化分析
以BmNPV日本T3株的gp64基因為標準參考,不同地域來源的BmNPV毒株為同一種群參考,親緣關系相近的同屬異種核型多角體病毒(NPV)為種群外參考,包括AcMNPV、薄荷灰夜蛾NPV(RoMNPV)和野桑蠶NPV(BomaNPV)。使用MegAlign中的ClustalW進行序列比對,并以MEGA 5.0的Kimura 2-parameter模式構建系統發(fā)育進化樹,Bootstrap重復設為1000。分別通過NetNGlyc 1.0 Server和NetOGlyc 4.0 Server預測蛋白編碼序列可能存在的N-糖基化位點和O-糖基化位點;以SWISS-MODEL在線建模預測編碼蛋白三級結構;根據系統發(fā)育進化樹的分群,對廣西BmNPV毒株的流行分布地點進行標識。
1. 6 BmNPV毒株毒力測定
將GXZZ和GXUA株分別接種至五齡起蠶進行復壯增殖,提純病毒多角體并以血球計數板計數,然后配制成濃度約109 PIBs/mL的病毒原液,取2 mL置于18 mL純水中進行10倍梯度稀釋,共配制10個梯度。各稀釋樣品4000 r/min離心10 min,棄上清液,僅留少量重懸多角體沉淀。將稀釋樣品分別均勻涂抹在2張適熟偏嫩的桑葉表面(10 cm×10 cm),稍晾干后給四齡和五齡起蠶餉食,每個稀釋梯度添食30頭家蠶,設2個組重復,待食盡再添新的桑葉,確保病毒被完全攝食,對照組桑葉涂抹純水。統計發(fā)生血液型膿病的病死家蠶及死籠繭數目,采用Reed-Muench法計算半數致死量(LD50)。
2 結果與分析
2. 1 廣西BmNPV毒株多角體的形態(tài)結構
在普通光學顯微鏡下觀察,廣西BmNPV毒株多角體外觀呈多角形,大小不等,表面有較強的折光性,呈暗藍色(圖1-A);在掃描電子顯微鏡下觀察發(fā)現多角體呈近似球形或方形的多面體,個體差異顯著,大型多角體的體積是小型多角體的2~3倍,且多面體的構成面存在數量和形狀上的差異(圖1-B),進一步說明野生BmNPV多角體結構存在不均質性。
2. 2 廣西BmNPV毒株gp64基因遺傳多態(tài)性
廣西BmNPV毒株gp64基因ORF長度存在3種情況(1590、1593和1599 bp),分別編碼含529、530和532個氨基酸殘基的GP64蛋白。其中,GXHJ和GXYZ2株為1593 bp,與標準參考T3株相同;GXNN、GXJX、GXYF、GXZP、GXZS、GXXZ、GXYJ、GXLeY、GXCW、GXTY、GXZZ和GXUA等12株毒株為1590 bp,均缺失第94~96位編碼丙氨酸的GCG密碼子;GXFM、GXHP、GXLuoC2、GXXD1、GXMS2和GXWX等6株毒株為1599 bp,則在第97~102位出現6個核苷酸插入,GXLuoC2和GXWX株為GCGCCG插入,編碼丙氨酸和脯氨酸,其余4株毒株為GTGCCG插入(表1),編碼纈氨酸和脯氨酸,故推測此處為1個高變位點,允許突變的程度較大。
20株廣西BmNPV毒株與標準參考T3株的gp64基因核苷酸序列同源性在97.6%~99.2%,其推導氨基酸序列同源性在96.4%~99.6%(表1)。其中,GXJX株與GXZZ株的gp64基因核苷酸序列完全一致;而GXYF、GXJX、GXNN和GXZZ等4株毒株的gp64基因推導氨基酸序列完全一致,GXFM、GXHP和GXMS2等3株毒株的gp64基因推導氨基酸序列也完全一致。廣西BmNPV毒株發(fā)生核苷酸替換突變的位點數目為13~28個,除GXTY株外,各毒株的同義替換位點數目為非同義替換位點的2~7倍,說明廣西BmNPV野生群體gp64基因發(fā)生點突變頻率較高,但大部分變異并未改變氨基酸編碼。其中,有20個位點的單核苷酸突變在廣西BmNPV毒株gp64基因中較常見,但在廣西BmNPV毒株中出現的數目和分布情況各不相同。根據gp64基因遺傳多態(tài)性分析結果可知,不同廣西BmNPV毒株gp64基因的編碼長度和突變程度存在明顯差異,說明從廣西不同蠶區(qū)采集的BmNPV毒株可能來源于同一祖先病毒株,但在不同地理環(huán)境及經不等傳代后,表現出進化的程度各不相同。
采用NetNGlyc 1.0 Server和NetOGlyc 4.0 Server對20株廣西BmNPV毒株及標準參考T3株的gp64基因推導氨基酸序列進行N-糖基化位點和O-糖基化位點預測,結果顯示:標準參考T3株有4個N-糖基化位點和2個O-糖基化位點,廣西BmNPV毒株的N-糖基化位點為3~4個(表1),部分毒株的預測位點也存在差異,如GXTY株為第5、175和213位,而GXUA株為第5、175和442位;除GXZS株外,其他廣西BmNPV毒株的O-糖基化位點均為2個,且預測位點一致。以SWISS-MODEL對廣西BmNPV毒株GP64蛋白序列進行同源建模,發(fā)現GP64蛋白三級結構皆為空間構型相似的同源三聚體。可見,gp64基因N端多核苷酸的插入或缺失突變,以及其他位點的單核苷酸突變尚未對編碼蛋白的空間構象產生明顯影響。
2. 3 廣西BmNPV毒株gp64基因遺傳進化樹
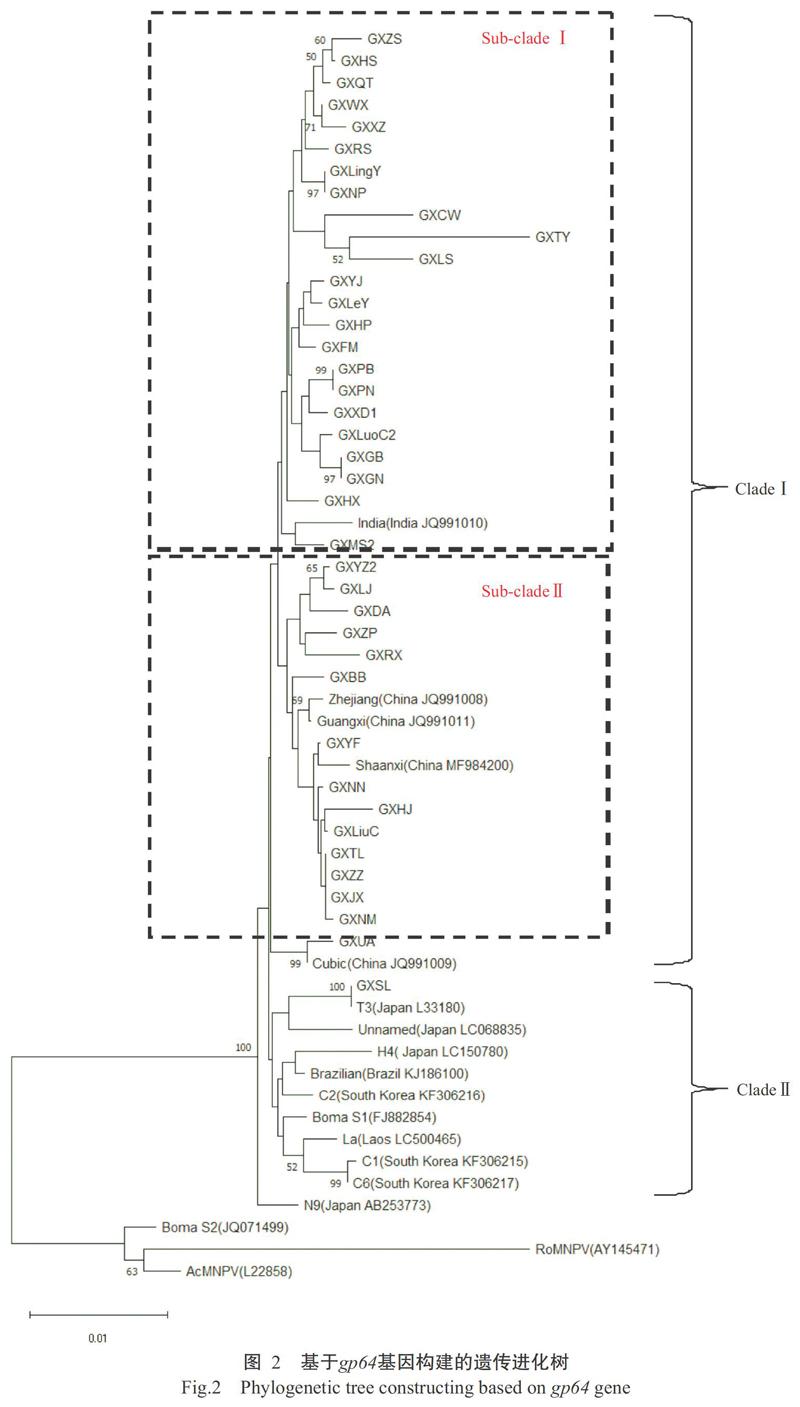
以GenBank已公布的14株國內外BmNPV毒株為群內參考,以AcMNPV、RoMNPV及BomaNPV(S1和S2)為群外參考,將本研究測序的20株廣西Bm-NPV毒株與本課題組前期測序的18株廣西BmNPV毒株(陳朝蓉等,2017),基于gp64基因構建遺傳進化樹。結果(圖2)顯示,所有BmNPV毒株位于同一大群中,而群外參考毒株中除BomaS1外,其余毒株均位于另一小群,說明gp64基因在種系發(fā)育上存在較大的進化距離。在BmNPV進化群中,除參考毒株N9外,其他BmNPV毒株被分成兩大群類(Clade I和Clade II)。幾乎所有的廣西BmNPV毒株聚類于Clade I,且又被分為2個主要亞群(Sub-clade I和Sub-clade II),其中GXUA株與參考毒株Cubic的遺傳關系最近,被單獨聚為一支。除參考毒株India外,Sub-clade I中全部為廣西BmNPV毒株,其他廣西BmNPV毒株和國內參考毒株則位于Sub-clade II。幾乎所有的國外參考毒株聚類于Clade II,廣西BmNPV毒株中僅GXSL株被歸類在該群類,是由于其gp64基因核苷酸序列與標準參考T3株完全一致(陳朝蓉等,2017)。可見,廣西BmNPV毒株的進化程度與國外參考毒株明顯不同,與國內參考毒株也存在一定差異。
2. 4 廣西BmNPV毒株的流行分布情況
根據gp64基因的遺傳進化樹Sub-clade I和Sub-clade II分群情況,繪制廣西BmNPV毒株的流行分布圖。結果(圖3)顯示,Sub-clade I毒株主要分布在廣西中南部和西部地區(qū),Sub-clade II毒株則主要分布在廣西中北部地區(qū),流行區(qū)域既呈集中連片,也有分散個體。在廣西河池、柳州和來賓地區(qū)2個亞群毒株的流行呈交織分布,而在廣西貴港和百色地區(qū)主要流行Sub-clade I毒株。
2. 5 廣西BmNPV毒株的致病力比較
以GXZZ株和GXUA株分別接種四齡和五齡起蠶,計算血液型膿病的病死率及毒株對各齡蠶的LD50。結果(圖4)發(fā)現,在相同的病毒多角體稀釋梯度下,GXUA株的平均死亡率均低于GXZZ株,且出現死亡的最高稀釋倍數小于GXZZ株。GXUA株對四齡和五齡起蠶的LD50分別為3.3和3.1,而GXZZ株對四齡和五齡起蠶的LD50分別為5.5和5.3,前者較后者低2個數量級,二者對四齡蠶的LD50均高于五齡起蠶。說明GXUA株對家蠶的致病力較GXZZ株弱,但二者對四齡和五齡起蠶的致病力相差不明顯。
3 討論
BmNPV標準參考T3株的gp64基因ORF編碼530個氨基酸殘基,起止位點為99864~101436 nt,與AcMNPV的gp64基因核苷酸序列同源性在95.0%以上(Gomi et al.,1999)。在BmNPV和AcMNPV的gp64基因ORF中潛在2個能成為翻譯起始位點的框架,且這2個起始密碼子相距54 nt(Rahman and Gopinathan,2003)。廣西BmNPV毒株gp64基因近N端的GCG缺失及GCGCCG/GTGCCG插入突變,均位于第2個翻譯起始框架內,對應AcMNPV的gp64基因信號肽區(qū),且突變氨基酸呈疏水性,堿基突變對編碼蛋白的三聚體空間構象未產生顯著影響,故推測這些突變發(fā)生在BmNPV的gp64基因信號肽區(qū),即與GP64蛋白基因其他功能區(qū)相比,gp64基因信號肽區(qū)在BmNPV進化過程中的保守性較低。
與AcMNPV不同,BmNPV的GP64蛋白信號肽長38 aa,在BmN和Bm5細胞中完整的GP64信號肽可將報告基因(eGFP)定位于細胞質膜,而不完整的GP64信號肽均無此項功能;在Sf9細胞中完整的GP64信號肽也將eGFP基因定位于細胞質膜,但部分缺失的BmNPV GP64信號肽也具有此功能,說明GP64信號肽具有宿主特異性(柳林,2018)。另有研究結果顯示,在BmNPV和AcMNPV的GP64蛋白序列中分別存在4和3個膽固醇識別位點,其區(qū)別在于BmNPV的GP64信號肽內含有1個膽固醇識別位點(1~13 aa),將該位點中保守的酪氨酸突變?yōu)楸彼岷螅瑢χ亟MBmNPV的感染性和致病力均無影響,但報告基因的表達有所增強(南文斌,2018)。BmNPV的GP64信號肽在引導成熟蛋白跨膜定位后未被切割,可能是其信號肽內的膽固醇識別位點與宿主膽固醇發(fā)生相互作用,且BmNPV的GP64蛋白加工分泌及介導入侵細胞途徑可能與AcMNPV的GP64蛋白有所不同(梁飛,2016;柳林,2018)。本研究結果表明,所有廣西BmNPV毒株及其他參考毒株的GP64信號肽膽固醇識別位點均非常保守,但GP64信號肽末端疏水性氨基酸的缺失或插入是否對信號肽酶切割產生影響尚有待進一步探究。桿狀病毒具有嚴格的宿主域,病毒關鍵蛋白的信號肽在進化過程中可能已發(fā)生種屬特異性變異,BmNPV的宿主域遠比AcMNPV窄,是否與攜帶膽固醇識別位點的GP64信號肽發(fā)揮限制作用有關也需進一步驗證。
桿狀病毒進化與宿主昆蟲進化并行,遵循宿主依賴性進化原則,宿主的抗病毒防御反應可能是其主要驅動因素之一(Nagamine and Sako,2016)。桿狀病毒的核心基因在進化過程中非常保守,基于核心基因構建的桿狀病毒系統發(fā)育進化樹顯示其聚類分群與宿主昆蟲的分類基本一致(Herniou and Jehle,2007;覃呂高等,2013)。在同一桿狀病毒種群中,核心基因的進化也相當保守,對廣西BmNPV毒株種群核心基因polh序列的分析結果顯示大部分毒株的polh基因核苷酸序列同源性大于99.9%,發(fā)生突變的位點很少(Liang et al.,2013)。可見,核心基因可能更適用于不同桿狀病毒種群的宏觀進化分析,但在微觀進化過程中無法全面反映進化事件發(fā)生的具體信息,因此對非核心基因的進化分析顯得尤為重要。本研究結果表明,與標準參考T3株相比,發(fā)生變異的廣西BmNPV毒株gp64基因核苷酸序列同源性為97.6%~99.2%,在基于gp64基因構建的遺傳進化樹中,廣西BmNPV毒株主要被分為兩大群類(Clade I和Clade II)。而本課題組前期研究發(fā)現,廣西BmNPV毒株的微管相關蛋白p10基因核苷酸序列同源性在98.1%~99.5%,根據p10基因構建的遺傳進化樹將廣西BmNPV毒株分為三大群類(Liang et al.,2013;陳朝蓉等,2017)。盡管基于不同基因的遺傳進化分析結果存在差異,但從BmNPV的群類分布結果來看均能有效反映出集中分布的特點及其傳播規(guī)律,鑒于gp64基因是BmNPV非常重要的致病性相關基因,故推測根據gp64基因構建的遺傳進化樹更具代表性。
廣西BmNPV毒株的gp64基因突變較頻繁,但大部分變異并未改變氨基酸編碼,僅在信號肽序列上存在差異,而導致其進化與其他參考BmNPV毒株保持一定距離。究其原因除了與當地的氣候環(huán)境有關外,還可能與當地飼養(yǎng)的家蠶品種存在一定關聯。Xu等(2013)以不同來源的BmNPV、BomaNPV和AcMNPV為研究對象,基于基因組序列構建遺傳進化樹,結果顯示除廣西和浙江2個地理分離BmNPV毒株被歸為同一分支外,其他地理分離毒株均單獨聚類為一分支,表明地理分離毒株間的基因型差異可能與地理來源有關。本研究基于gp64基因構建遺傳進化樹,結果顯示,幾乎所有的廣西BmNPV毒株聚類于Clade I,而幾乎所有的國外參考毒株聚類于Clade II,進一步證實BmNPV的基因進化與地理來源相關,且與基于基因組構建遺傳進化樹的進化分群結果(Xu et al.,2013)基本一致。
BmNPV的GP64蛋白糖基化修飾被抑制,其誘導的膜融合活性降低,具有傳染性的出芽型病毒粒子產量減少(Rahman and Gopinathan,2003)。Jarvis等(1998)研究表明,當GP64蛋白第198、355、385和426位的N-糖基化位點單獨或共同發(fā)生突變時,AcMNPV產生感染性子代病毒的水平明顯低于野生型,且突變型出芽型病毒粒子的受體結合能力受損,感染擴散的速度變慢。在本研究中,GXUA株GP64蛋白的N-糖基化位點比GXZZ株少1個,其對家蠶的致病力也較GXZZ株弱,推測是由于糖基化位點改變而影響GP64蛋白的折疊及構象,進而影響病毒的復制增殖,導致其致病力減弱。廣西的蠶桑主產區(qū)多為老蠶區(qū),分布區(qū)域集中,病原種類繁多且經年積累。廣西蠶區(qū)的BmNPV流行分布呈集中性與分散性并存,反映了病毒傳播的廣泛性和復雜性,給血液型膿病的防控帶來極大難度。在缺乏有效藥物防治的情況下,及時預警、精準防控及群防群控等措施有助于阻止BmNPV的流行和傳播。只有從根本上減少蠶區(qū)BmNPV多角體累積,才能有效降低家蠶血液型膿病的發(fā)生概率,實現廣西蠶桑產業(yè)的持續(xù)健康發(fā)展。
4 結論
廣西BmNPV毒株gp64基因在進化過程中其信號肽區(qū)出現明顯變異,發(fā)生同義突變的頻率較高,形成較獨立的進化分群,毒株間的致病力差異可能與GP64蛋白糖基化位點不同有關,說明廣西BmNPV毒株具有不同的基因型和表型,在一定程度上維持了BmNPV野生群體的遺傳多樣性。
參考文獻:
陳朝蓉,屈達才,朱方容,李俊,閉立輝,梁湘. 2017. 廣西家蠶核型多角體病毒株gp64基因克隆與序列分析[J]. 南方農業(yè)學報,48(2):328-335. doi:10.3969/j:issn.2095-1191. 2017.02.328. [Chen C R,Qu D C,Zhu F R,Li J,Bi L H,Liang X. 2017. Cloning and sequence analysis of gp64 gene in Bombyx mori nucleopolyhedrovirus Guangxi isolates[J]. Journal of Southern Agriculture,48(2):328-335.]
梁飛. 2016. BmNPV膜融合蛋白GP64作用機制初步研究[D]. 鎮(zhèn)江:江蘇科技大學. [Liang F. 2016. A preliminary study on mechanism of BmNPV envelope fusion protein GP64[D]. Zhenjiang:Jiangsu University of Science and Technology.]
柳林. 2018. 量子點標記BmNPV芽生病毒粒子研究及GP64信號肽分析[D]. 鎮(zhèn)江:江蘇科技大學. [Liu L. 2018. Study on quantum dots labeling of BmNPV BV and ana-lysis GP64 signal peptide[D]. Zhenjiang:Jiangsu University of Science and Technology.]
魯興萌. 2012. 家蠶對血液型膿病的抗性與防治策略[J]. 中國蠶業(yè),33(3):4-7. doi:10.16839/j.cnki.zgcy.2012.03. 002. [Lu X M. 2012. Resistance and prevention strategy of Bombyx mori to nuclear polyhedrosis[J]. China Sericulture,33(3):4-7.]
南文斌. 2018. BmNPV膜融合蛋白GP64中膽固醇結合位點活性分析[D]. 鎮(zhèn)江:江蘇科技大學. [Nan W B. 2018. Analysis of cholesterol-binding site activity in GP64 of BmNPV membrance fusion protein[D]. Zhenjiang:Jiangsu University of Science and Technology.]
覃呂高,屈達才,李俊,徐周徐. 2013. 家蠶抗核型多角體病毒基因與蛋白的研究進展[J]. 南方農業(yè)學報,44(5):854-858. doi:10.3969/j:issn.2095-1191.2013.5.854. [Qin L G,Qu D C,Li J,Xu Z X. 2013. Research progress in silkworm gene and protein against Bombyx mori nuclear polyhedrosis virus[J]. Journal of Southern Agriculture,44(5):854-858.]
王霞,黃旭華,蔣滿貴,唐亮,董桂清,黃深惠,石美寧,潘志新. 2020. 廣西家蠶血液型膿病發(fā)病因子及其病原分子系統發(fā)育分析[J]. 南方農業(yè)學報,51(3):669-676. doi:10.3969/j.issn.2095-1191.2020.03.025. [Wang X,Huang X H,Jiang M G,Tang L,Dong G Q,Huang S H,Shi M N,Pan Z X. 2020. Epidemic factors of Bombyx mori hemolymph-type septic disease in Guangxi and its molecular phylogenetic analysis[J]. Journal of Southern Agriculture,51(3):669-676.]
夏定國,張國政,王文兵,趙巧玲,唐順明. 2007. gp64基因相應dsRNA對家蠶核型多角體病毒(BmNPV)增殖的抑制[J]. 中國農業(yè)科學,40(12):2882-2887. [Xia D G,Zhang G Z,Wang W B,Zhao Q L,Tang S M. 2007. Inhibitory effects of the corresponding dsRNA of gp64 gene on BmNPV multiplication[J]. Scientia Agricultura Sinica,40(12):2882-2887.]
Deo V K,Yui M,Alam J,Yamazaki M,Kato T,Park E Y. 2014. A model for targeting colon carcinoma cells using single-chain variable fragments anchored on virus-like particles via glycosyl phosphatidylinositol anchor[J]. Pharmaceutical Research,31(8):2166-2177. doi:10.1007/s11095-014-1316-4.
Fuxa J R. 2004. Ecology of insect nucleopolyhedroviruses[J]. Agriculture,Ecosystems & Environment,103(1):27-43. doi:10.1016/j.agee.2003.10.013.
Gomi S,Majima K,Maeda S. 1999. Sequence analysis of the genome of Bombyx mori nucleopolyhedrovirus[J]. The Journal of General Virology,80(5):1323-1337. doi:10. 1099/0022-1317-80-5-1323.
Herniou E A,Jehle J A. 2007. Baculovirus phylogeny and evolution[J]. Current Drug Targets,8(10):1043-1050. doi:10. 2174/138945007782151306.
Hitchman R B,Hodgson D J,King L A,Hails R S,Cory J S,Possee R D. 2007. Host mediated selection of pathogen genotypes as a mechanism for the maintenance of baculovirus diversity in the field[J]. Journal of Invertebrate Pathology,94(3):153-162. doi:10.1016/j.jip.2006.10.002.
Jarvis D L,Wills L,Burow G,Bohlmeyer D A. 1998. Mutational analysis of the N-linked glycans on Autographa californica nucleopolyhedrovirus gp64[J]. Journal of Virology,72(12):9459-9469. doi:10.1128/JVI.72.12.9459-9469.1998.
Jiang L,Zhao P,Wang G H,Cheng T C,Yang Q,Jin S K,Lin P,Xiao Y,Sun Q,Xia Q Y. 2013. Comparison of factors that may affect the inhibitory efficacy of transgenic RNAi targeting of baculoviral genes in silkworm,Bombyx mori[J]. Antiviral Research,97(3):255-263. doi:10. 1016/j.antiviral.2012.12.020.
Kato T,Suzuki F,Park E Y. 2012. Display of the human (pro) renin receptor on Bombyx mori nucleopolyhedrovirus(BmNPV) particles using Bm cells[J]. Journal of Bioscience and Bioengineering,114(5):564-569. doi:10.1016/ j.jbiosc.2012.06.008.
Katou Y,Ikeda M,Kobayashi M. 2006. Abortive replication of Bombyx mori nucleopolyhedrovirus in Sf9 and High Five cells:Defective nuclear transport of the virions[J]. Virology,347(2):455-465. doi:10.1016/j.virol.2005.11.043.
Li G H,Tang Q,Chen H Q,Yao Q,Ning D G,Chen K P. 2011. Display of Bombyx mori nucleopolyhedrovirus GP64 on the Bacillus subtilis spore coat[J]. Current Microbio-logy,62(5):1368-1373. doi:10.1007/s00284-011-9867-7.
Li Z F,Blissard G W. 2009. The pre-transmembrane domain of the Autographa californica multicapsid nucleopolyhedrovirus GP64 protein is critical for membrane fusion and virus infectivity[J]. Journal of Virology,83(21):10993-11004. doi:10.1128/JVI.01085-09.
Liang X,Lu Z L,Wei B X,Feng J L,Qu D C,Luo T R. 2013. Phylogenetic analysis of Bombyx mori nucleopolyhedrovirus polyhedrin and p10 genes in wild isolates from Guangxi Zhuang Autonomous Region,China[J]. Virus Genes,46(1):140-151. doi:10.1007/s11262-012-0820-z.
Monsma S A,Oomens A G,Blissard G W. 1996. The GP64 envelope fusion protein is an essential baculovirus protein required for cell-to-cell transmission of infection[J]. Journal of Virology,70(7):4607-4616. doi:10.1128/JVI. 70.7.4607-4616.1996.
Nagamine T,Sako Y. 2016. A role for the anti-viral host defense mechanism in the phylogenetic divergence in baculovirus evolution[J]. PLoS One,11(5):e0156394. doi:10. 1371/journal.pone.0156394.
Rahman M M,Gopinathan K P. 2003. Characterization of the gene encoding the envelope fusion glycoprotein GP64 from Bombyx mori nucleopolyhedrovirus[J]. Virus Resea-rch,94(1):45-57. doi:10.1016/s0168-1702(03)00123-0.
Washburn J O,Chan E Y,Volkman L E,Aumiller J J,Jarvis D L. 2003. Early synthesis of budded virus envelope fusion protein GP64 enhances Autographa californica multicapsid nucleopolyhedrovirus virulence in orally infected Heliothis virescens[J]. Journal of Virology,77(1):280-290. doi:10.1128/jvi.77.1.280-290.2003.
Xu Y P,Cheng R L,Xi Y,Zhang C X. 2013. Genomic diversity of Bombyx mori nucleopolyhedrovirus strains[J]. Genomics,102(1):63-71. doi:10.1016/j.ygeno.2013.04.015.
Zheng H,Ren F F,Lu Q Y,Cao Z M,Song J C,Feng M,Liu J S,Sun J C. 2019. An efficient method for multigene co-interference by recombinant Bombyx mori nucleopolyhedrovirus[J]. Molecular Genetics and Genomics,294(1):111-120. doi:10.1007/s00438-018-1491-9.
Zhou J,Blissard G W. 2008. Identification of a GP64 subdomain involved in receptor binding by budded virions of the baculovirus Autographica californica multicapsid nucleopolyhedrovirus[J]. Journal of Virology,82(9):4449-4460. doi:10.1128/JVI.02490-07.
(責任編輯 蘭宗寶)
