引入豆科作物的輪作模式對設施蔬菜土壤微生物群落組成的影響
2021-07-01 08:07:38茹淑華孫世友張國印馬俊賢
華北農學報 2021年3期
劉 蕾,徐 夢,王 凌,茹淑華,孫世友,郜 靜,李 玭,張國印,馬俊賢
(1.河北省農林科學院 農業資源環境研究所,河北省肥料技術創新中心,河北 石家莊 050051;2.中國科學院 地理科學與資源研究所,生態網絡觀測與模擬重點實驗室,北京 100101;3.河北省市場監督管理局,河北 石家莊 050091)
由于我國耕地資源短缺,發展設施農業是推進農業結構調整、加快農業供給結構性改革的主要抓手和突破口,也是農民增收、農業增效、農村經濟和社會發展的重要推動力。然而,為追求高產,設施蔬菜通常種植指數高、輪作周期短,且水肥、農藥投入量高,加上常年處于高溫、高濕、高蒸發、無降水淋洗的特殊環境,極易導致土壤酸化、次生鹽漬化、土傳病害等問題,同時土壤中過量氮磷養分的淋洗會導致水環境污染。這些產區土壤質量退化和生態環境惡化可能進一步影響蔬菜生長過程,不僅導致減產減收,更會威脅食品安全和人類健康[1]。
以往研究中通常使用土壤理化性狀來表征或者評估設施栽培條件對土壤質量、蔬菜產量等的影響[2-4]。然而,越來越多的證據表明,傳統的理化指標逐漸難以滿足對土壤質量與土壤健康研究的需要,而土壤生物學性質則能敏感地反映出土壤環境和健康的變化[5]。土壤微生物是土壤中的關鍵組分,在促進土壤養分循環利用[6]、保持土壤健康和質量[7]以及污染物降解[8]等過程中發揮著重要作用,是評價土壤質量的重要指標[9]。土壤微生物對環境變化十分敏感,因此,通過土壤微生物性狀的變化評估設施土壤質量變化近年來成為研究設施土壤質量保持和功能恢復的熱點[10-11]。研究表明,設施農業種植方式顯著改變土壤理化性質,進而影響微生物群落,通常表現為多樣性隨種植年限顯著降低[12-13]或者群落組成改變(例如土壤細菌與真菌比值降低、有益菌群減少、病原微生物數量增加等)[10,14-17]。例如,相比于露地蔬菜種植,塑料大棚土壤微生物多樣性顯著降低,且大棚生產條件增加了微生物的環境脅迫進而導致群落退化[18]。Zhang等[10]在壽光連續11 a試驗發現,一年兩季番茄栽培使得土壤微生物組成朝著有益菌豐度降低、病原微生物積累的方向演變。大量研究表明,土壤微生物群落失衡是導致設施蔬菜土傳病害、連作障礙等加劇的主要原因[19-20]。因此,設施蔬菜種植過程中土壤微生物多樣性、群落組成和結構的變化可以作為土壤受損程度和質量變化的重要預警指標,理解土壤微生物及功能群的變化對揭示設施蔬菜栽培條件引起的土壤質量退化微生物學機制及保持設施土壤可持續性具有重要意義。然而,與農田相比,設施土壤微生物研究在我國起步較晚,且多集中在種植年限[10-11,13,15,17,21-22]和施肥[23-25]對設施土壤微生物區系的影響,對不同輪作模式下土壤微生物群落結構與功能變化的認識仍有很大的局限性。
1 材料和方法
1.1 試驗區概況
本試驗在河北衡水國家農業科技園區饒陽縣大尹村鎮南北巖村(115°50′82″E,38°16′14″N)進行,饒陽縣設施蔬菜經過30多年發展,種植規模達千公頃,居全省第一。研究區地屬冀中平原黑龍港流域,地貌類型為滹沱河洪積平原,南北巖村設施大棚建于廢棄河床之上,土壤類型為潮土,質地為沙壤,地下水埋深3~7 m。年均溫12.2 ℃,降雨量552.6 mm,屬于溫帶大陸季風氣候。該區典型種植模式為西紅柿-甜瓜輪作,秋冬茬為番茄(8-12月),平均產量為67 500~90 000 kg/hm2;冬春茬為甜瓜(1-7月),平均產量約75 000 kg/hm2。
1.2 試驗設計
定位試驗開始于2018年6月,在典型西紅柿-甜瓜輪作中通過引入養分、水分需求量更低的豆角,調整種植制度,共設置3個處理:西紅柿-甜瓜、豆角-甜瓜、西紅柿-豆角,養分和水分投入量如表1所示,每個處理3次重復,小區面積21.5 m2。試驗開始前土壤基礎理化性狀:容重1.01 g/cm3,黏粒9.6%,砂粒56.6%,有機質45.9 g/kg,NO3--N 147.1 mg/kg,Olsen-P 274.0 mg/kg。
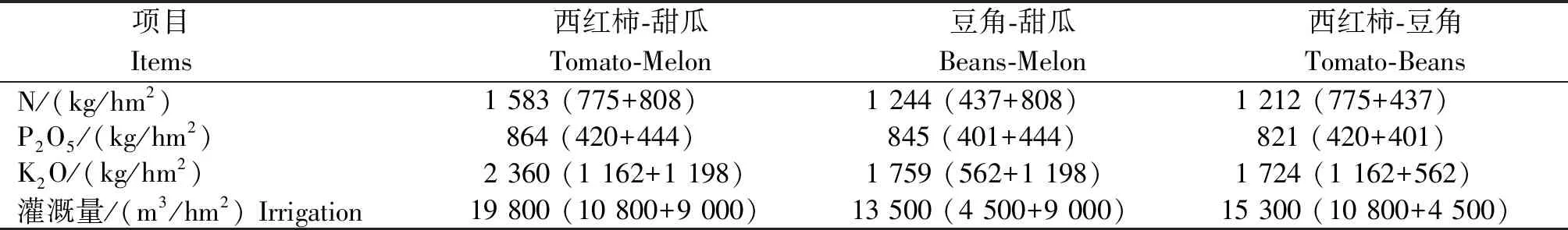
表1 不同輪作模式下周年養分及水分投入量Tab.1 Annual input of fertilizer and water under different rotation systems
1.3 樣品采集
土壤樣品采集于2020年7月28日西紅柿、豆角拉秧并清理完全后進行,每個小區選取3個樣方(1 m×1 m),樣方間距3 m,每個樣方中隨機用土鉆采集5個表層土壤(0~20 cm)并混合,即每個小區15鉆混合為一個樣品重復,一共采集9個土壤樣品。去除石塊、落葉等雜質后充分混勻,分成2份,過2 mm篩,一份用于微生物測定,保存于-20 ℃,一份用于土壤化學性質測定,保存于4 ℃冰箱。
1.4 土壤化學性質測定
土壤有機質用油浴加熱重鉻酸鉀氧化-容量法測定,土壤全鉀用堿熔-火焰光度法測定,土壤總氮用凱氏蒸餾法,土壤硝態氮用紫外分光光度法測定,土壤pH值用1 mol/L KCl浸提(水土比1.0∶2.5)電位法測定,水溶性鹽采用電導法測定,土壤總磷用氫氧化鈉熔融-鉬銻抗比色法,土壤Olsen-P 0.5 mol/L NaHCO3(pH值8.5)溶液浸提(水土比20∶1)鉬銻抗比色法測定[26]。
1.5 土壤微生物測定
使用OMEGA M5635-02 Soil DNA Kit試劑盒提取土壤DNA,使用NanoDrop ND-2000分光光度計(Thermo Scientific,Wilmington,DE,美國)測定DNA 的質量,送至上海派森諾公司進行Illumina MiSeq高通量測序,使用引物338F(5′-ACTCCTACGGGAGGC
AGCA-3′)和806R(5′-GGACTACHVGGGTWTCTAA
T-3′)對細菌16S rDNA V3V4基因進行擴增,引物ITS5F(5′-GGAAGTAAAAGTCGTAACAAGG-3′)和ITS1R(5′-GCTGCGTTCTTCATCGATGC-3′)對真菌ITS基因進行擴增。測序得到的原始序列經過QⅡME2、DADA2進行質控拼接,去重后得到擴增子序列變異體(Amplicon Sequence Variants,ASV)。采用QⅡME2的classify-sklearn算法對于每個ASV的特征序使用預先訓練好的Naive Bayes分類器進行物種注釋,得出物種分類水平,對比數據庫為Greengenes(Release 13.8,http://greengenes.secondgenome.com/)和UNIT (Release 8.0,https://unite.ut.ee/)。最后對采用QⅡME2的qiime feature-table rarefy功能進行抽平。采用QⅡME2的qiime diversity alpha-rarefaction命令,設置最小抽平深度為10、全體樣本中最低測序深度樣本序列量的95%,再在這一深度與最小深度之間均勻選取10個深度值,每個深度值抽平10次,計算所選的多樣性指數。選取最大抽平深度時的得分平均值作為α-多樣性指數。以Chao指數表征豐富度,以Shannon指數表征多樣性,以Pielou指數表征均勻度。
(1)給予兩組患者同樣的基礎用藥,指導患者合理使用克拉霉素片(廣東逸舒制藥股份有限公司;國藥準字H20000207;0.125g*24s)、阿莫西林分散片(石藥集團中諾藥業(石家莊)有限公司;國藥準字H10980075;0.25g*6s))、甲硝唑片(湖南漢森制藥股份有限公司;國藥準字H43020225;0.2g*21s);(2)對照組在基礎用藥基礎上加用奧美拉唑(廣東逸舒制藥有限公司;國藥準字H20040911;20mg*7S);實驗組在對照組基礎上加用雷貝拉唑(濟川藥業集團有限公司;國藥準字H20052317;10mg*7S)。
1.6 數據分析
使用單因素方差分析(ANOVA)比較3種種植制度下土壤化學性質、微生物α-多樣性、細菌及真菌各種群相對豐度的差異,并用Duncan′s multiple range test檢驗各處理間差異的顯著性(P<0.05水平下)。使用Spearman相關性分析土壤微生物多樣性指數及各種群相對豐度與土壤化學性質的相關性。方差分析和相關性分析使用IBM SPSS Statistics(v 18.0)進行。不同種植方式下共有/特有的ASV韋恩圖計算采用R語言VennDiagram進行。利用隨機森林模型(Random Forest)預測顯著影響3種輪作模式下土壤微生物群落結構差異的物種(屬水平)及其貢獻,使用R語言RandomForest進行運算分析,每個物種對預測模型貢獻的顯著性用軟件包rfPermute計算。微生物群落結構及其影響因子使用典型相關分析(Canonical Correlation Analysis,CCA),3種輪作模式下群落結構的差異用PERMANOVA進行檢驗,使用R語言Vegan包進行。
2 結果與分析
2.1 土壤基礎理化性質
相比于傳統茄科-葫蘆科輪作,引入豆科作物的輪作模式每年降低N-P2O5-K2O投入21.38%~23.46%,2.27%~5.04%,25.45%~26.97%,降低水分投入22.73%~31.82%,經過2 a四季肥水管理,不同設施輪作模式主要對土壤硝態氮含量和pH值產生了顯著的影響(表2):土壤NO3-N含量在豆角-甜瓜模式下最高,而在西紅柿-豆角模式下最低,相比于傳統西紅柿-甜瓜,西紅柿-豆角土壤NO3-N含量降低了65.31%;西紅柿豆角模式下土壤pH值顯著高于其他2種輪作模式,提高了4.4%~6.8%。
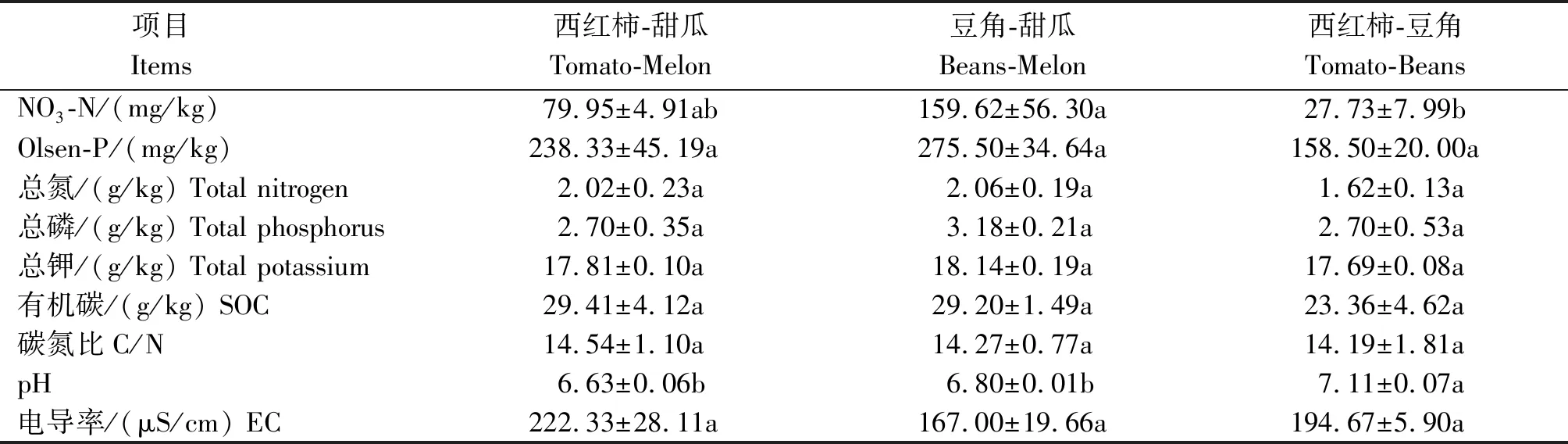
表2 不同輪作模式下土壤化學性質Tab.2 Soil general properties under different rotations
2.2 土壤細菌和真菌α-多樣性
高通量測序數據經過過濾、去燥、拼接、去除嵌合體及數量為1的序列后,共得到405 515個細菌序列和1 050 999個真菌有效序列。去重后土壤細菌得到9 638個ASVs,不同輪作模式下ASV數量差異不顯著(P>0.05)。土壤真菌獲得1 146個ASVs,其中西紅柿-豆角模式下土壤真菌ASV數量顯著低于另2種處理(P<0.05)。
不同輪作模式顯著影響土壤微生物α-多樣性,但是細菌和真菌的響應特征有區別(表3)。輪作模式顯著影響了土壤細菌的均勻度(P<0.05),其Pielou指數在西紅柿-甜瓜模式下最高,在豆角-甜瓜模式下最低。而土壤真菌的豐富度對輪作模式有顯著響應,其Chao指數在西紅柿-豆角模式下顯著低于其他2種模式。
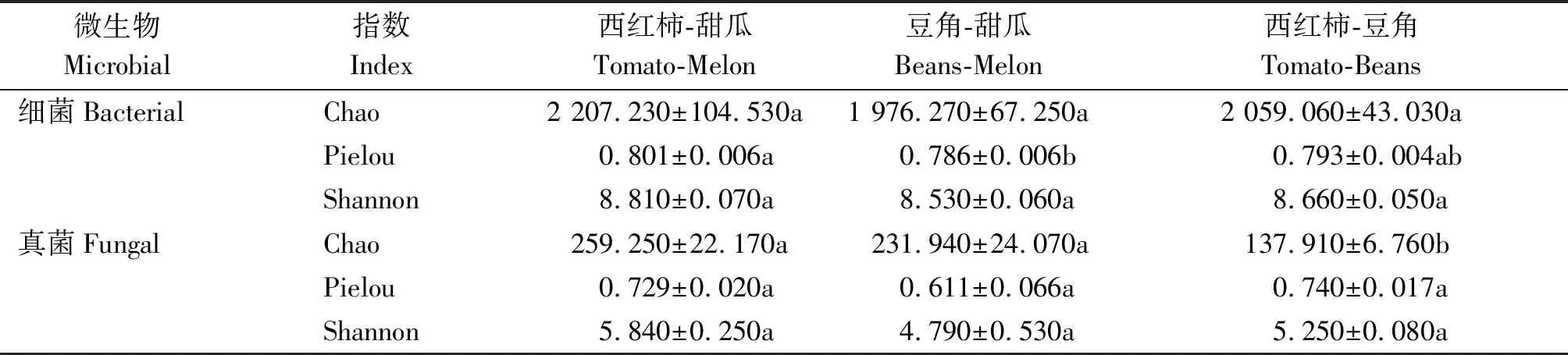
表3 不同輪作模式下土壤微生物群落α-多樣性指數Tab.3 Diversity indices of soil microbial community under different rotations
2.3 微生物群落組成和結構差異
高通量測序獲得的土壤細菌ASVs分屬于31門100綱231目372科617屬,且群落中優勢種群(相對豐度大于1%)在不同輪作模式土壤中的分布具有明顯差異(圖1)。在門水平上,己科河菌門(Rokubacteria)的平均相對豐度在西紅柿-豆角模式下((3.15±0.05)%)顯著高出西紅柿-甜瓜((1.36±0.51)%)和豆角-甜瓜((1.37±0.29)%)約2.3倍。在科水平上,西紅柿-豆角土壤中具有顯著(P<0.05)較高的亞硝化單胞菌科(Nitrosomonadaceae)((6.20±0.59)%)、Rokubacteriales((3.15±0.29)%)和S085((1.73±0.02)%);西紅柿-甜瓜種植制度顯著提高了Roseiflexaceae((6.21±0.07)%)和生絲單胞菌科(Hyphomonadaceae)((1.41±0.14)%)的相對豐度;豆角-甜瓜模式則顯著提高了甾體桿菌科(Steroidobacteraceae)的相對豐度((1.41±0.27)%)。
土壤真菌ASVs分屬于9門28綱55目117科174屬,其中子囊菌門(Ascomycota)在3種輪作模式下均是土壤中的優勢種群,其平均相對豐度能達到54.37%~69.50%。在門水平下,擔子菌門(Basidiomycota)的平均相對豐度在西紅柿-豆角種植制度下((13.71±2.50)%)顯著最高,分別是西紅柿-甜瓜((2.87±0.44)%)和豆角-甜瓜((3.35±0.96)%)的4.8,4.1倍(圖2)。土壤真菌優勢科中,小不整球殼科(Plectosphaerellaceae)的相對豐度在西紅柿-豆角種植制度下顯著最高((4.96±0.17)%),是西紅柿-甜瓜((1.62±0.93)%)和豆角-甜瓜((1.56±0.51)%)的約3倍。蟲草菌科(Cordycipitaceae)和叢赤殼科(Nectriaceae)則在西紅柿-豆角種植制度土壤富集,其相對豐度分別是(5.24±1.99)%,(1.34±0.25)%,顯著高于其余2種種植制度(分別是蟲草菌科(0.52±0.33)%,(0.37±0.19)%和叢赤殼科(0.71±0.36)%,(0.38±0.21)%)。
韋恩圖表明,土壤真菌群落在3種種植制度下的特異性更強(圖3)。真菌群落中,3種植制度下共有ASV(66個)占總真菌ASV的5.76%,而共有細菌ASV(1 016個)則占總細菌ASV的10.54%。并且,西紅柿-甜瓜、豆角-甜瓜2種種植制度下特有ASV占總ASV的比例均在真菌群落(35.08%,28.45%)大于細菌群落(23.60%,23.27%)。但是在西紅柿-豆角模式下真菌特有ASV占總ASV的比例(14.38%)則明顯低于細菌(27.43%)。對于2種模式下共有的ASV,無論是細菌還是真菌,均在當季作物相同情況下(即西紅柿-甜瓜和豆角-甜瓜,當季為甜瓜)數量最多(細菌652個,真菌107個)。
隨機森林模型揭示了對預測3種種植制度下土壤微生物群落構建差異有顯著(P<0.05)貢獻的細菌和真菌種群。土壤細菌群落構建在3種種植制度下的差異主要受SWB02(貢獻量2.15%)、0319-6G20(1.95%)、Blastocatella(1.95%)、Dokdonella(1.92%)等種群的顯著影響,且這些種群大部分屬于Proteobacteria、Acidobacteria、Actinobacteria等主要細菌門。其中,Gemmatirosa(Gemmatimonadetes)、SWB02(Proteobacteria)等種群在西紅柿-甜瓜土壤中顯著富集;Microbispora(Actinobacteria)、Dokdonella(Proteobacteria)等種群在豆角-甜瓜土壤中顯著富集;MND1(Proteobacteria)、IMCC26256(Acidobacteria)等種群在西紅柿-豆角土壤中顯著富集(圖4)。對于土壤真菌,顯著影響不同種植制度下群落構建差異的屬主要為Verticillium(貢獻量3.20%)、Blastobotrys(3.17%)、Kernia(2.91%)、Dokmaia(2.30%)等8個屬,集中在2個門,并且不屬于群落中的優勢種,相對豐度均低于5%,其中Blastobotrys(Ascomycota)、Kernia(Ascomycota)顯著在西紅柿-甜瓜土壤中富集;Verticillium(Ascomycota)、Dokmaia(Ascomycota)、Ceriporia(Basidiomycota)顯著在西紅柿-豆角土壤中富集(圖5)。
2.4 不同輪作模式下土壤微生物群落差異的驅動因子
相關分析表明,土壤pH值與真菌多樣性(Chao指數)有極顯著負相關性(P<0.01),而細菌多樣性指數則與土壤理化性質沒有顯著相關性(表4)。細菌和真菌優勢種群的相對豐度也顯著受到土壤理化性質的影響:其中,相對豐度與pH值、NO3-N和土壤總氮的相關性最強,與Olsen-P、土壤總鉀和C/N的相關性次之(表4)。大部分細菌種群的相對豐度受到多個土壤性質的綜合影響,并且不同種群與土壤因子的相關性有明顯區別。例如,Rokubacteriales、OM190與NO3-N顯著負相關,與pH值顯著正相關;而Roseiflexaceae、鏈孢囊菌科(Streptosporangiaceae)則與NO3-N顯著正相關,與pH值顯著負相關。相比于細菌,大部分真菌種群僅受到1~2個土壤因子的影響,并且這些種群相對豐度與土壤因子的相關性基本一致:例如,小不整球殼科(Plectosphaerellaceae)與NO3-N極顯著負相關,蟲草菌科(Cordycipitaceae)等與NO3-N顯著負相關,而與pH值顯著正相關。
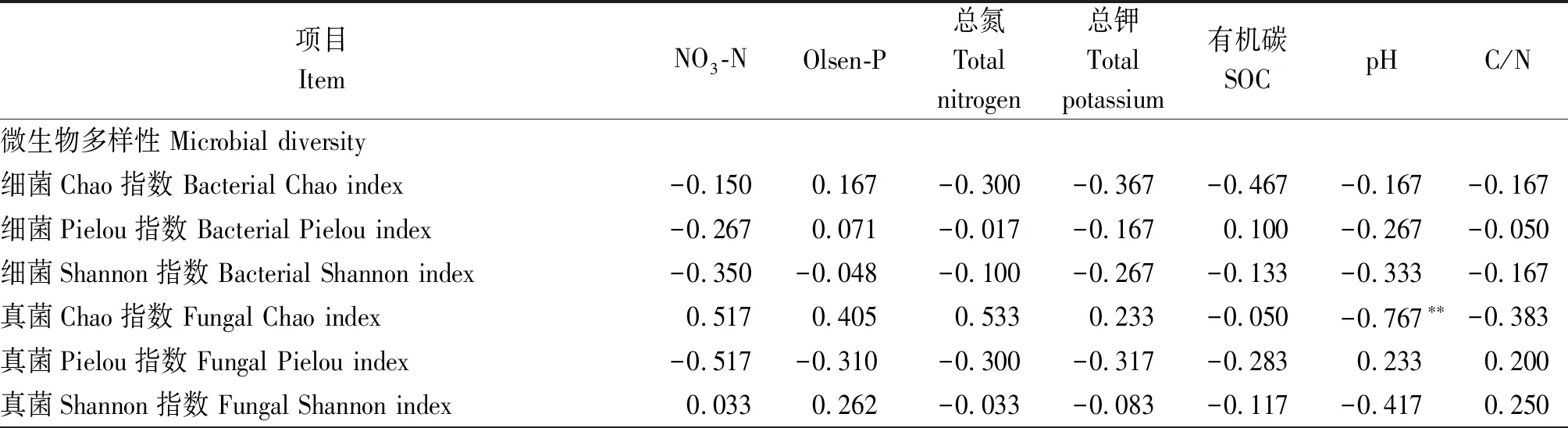
表4 微生物多樣性指數及相對豐度與土壤理化學性質的相關性Tab.4 Spearman correlation coefficients (r) of diversity indices and the relative abundance of microbial taxa with soil general properties
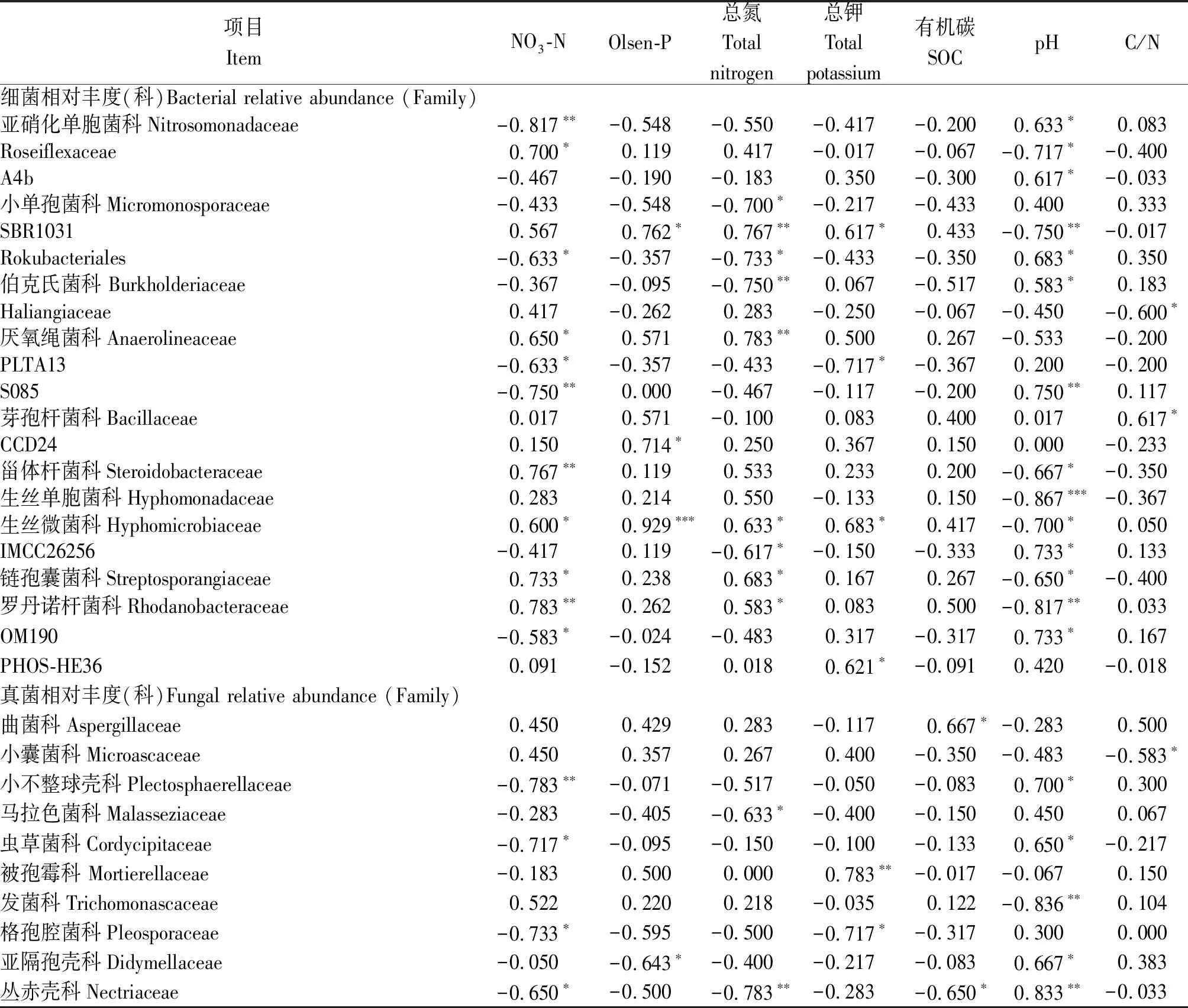
表4(續)
典范對應分析(CCA)進一步揭示了影響土壤微生物群落結構的關鍵環境因子(圖6)。CCA結果表明,土壤因子對細菌和真菌群落變化的解釋度分別為33.61%,32.12%。在所有環境因子里,土壤NO3-N和pH值是顯著影響細菌和真菌群落結構的關鍵因子(P<0.05)。此外,當季作物類型也對土壤微生物群落結構有顯著影響:PERMANOVA檢驗結果表明,當季作物為甜瓜的種植制度(西紅柿-甜瓜+豆角-甜瓜)下土壤細菌和真菌的群落結構與當季作物為豆角的模式(西紅柿-豆角)產生顯著差異(細菌F=2.35,P=0.01;真菌F=1.88,P=0.01)。
3 討論與結論
設施蔬菜高集約化生產方式及不合理的水肥管理極易造成土壤酸化、次生鹽漬化等土壤質量退化,而土壤微生物對環境變化十分敏感,因此,通過土壤微生物性狀的變化評估設施土壤質量變化近年來成為研究設施土壤質量保持和功能恢復的熱點[10-11]。然而,與農田相比,設施土壤微生物研究在我國起步較晚,且多集中在種植年限[10,11,13,15,17,22]和施肥[23-25]對設施土壤微生物區系的影響,對不同輪作模式下土壤微生物群落結構與功能的變化認識仍有很大的局限性。特別是引入豆科作物的輪作模式,其增產[28-29]和環保[30-33]效果雖得到普遍認可,然而該體系對土壤細菌和真菌群落的綜合影響及其驅動因子卻鮮有報道。本研究結果發現,設施蔬菜種植中,當季作物種類(豆角/甜瓜)顯著影響了土壤微生物多樣性、群落組成和結構。Ridl等[34]研究氮磷鉀肥料和種植不同作物(辣根、茄子、煙草)對土壤微生物群落的影響時同樣指出,作物種類對細菌群落結構產生顯著影響,其作用效果甚至大于施肥。而這種差異的主要原因在于,植物與土壤微生物在相互作用過程中會形成植物-微生物協同調控機制:即植物通過分泌碳源物質調控根際土壤微生物的種類和結構;同時,微生物也會通過產生激素類物質、抑制病原菌生長、誘導植物對生物及非生物脅迫產生抗性等方式促進植物的生長[35-36]。引入豆科作物的輪組模式顯著改變了碳源底物、并且水肥管理差異也改變了土壤理化性狀,從而調控微生物組成和活性。在所有土壤因子中,pH值和NO3-N對土壤微生物群落結構變化的影響最為顯著。由于設施環境極端性及復雜性,目前,報道的設施土壤微生物群落組成及結構的驅動因子各不相同,甚至相互矛盾。例如,Song等[17]指出土壤pH值和粘粒含量是影響設施土壤細菌群落的主要因素,而影響真菌群落的主要因素則為pH值和磷含量。Yao等[18]在對比設施及露地蔬菜土壤微生物群落結構時指出設施土壤微生物群落隨種植年份發生顯著改變,然而,Tian等[11]在調查環渤海區設施土壤微生物后卻發現微生物群落與栽培年份關系并不顯著,而是與土壤鉀含量密切相關。這些明顯不同甚至相悖的研究結果表明,對土壤微生物結構與功能的認識還十分有限,特別是顯著差別與農田生態系統的設施土壤。
本研究3種輪作模式中,豆角-甜瓜和西紅柿-豆角相比于傳統西紅柿-甜瓜輪作模式分別減少N-P2O5-K2O投入21.38%~23.46%,2.27%~5.04%,25.45%~26.97%,降低水分投入22.73%~31.82%,經過2 a四季肥水管理,西紅柿-豆角輪作模式相對于傳統西紅柿-甜瓜,NO3-N含量顯著降低了65.31%。并且由于氮肥減投緩解了土壤酸化,使得pH值相對于傳統模式提高了6.8%。本研究結果也表明,正是pH值和NO3-N含量的差異驅動了不同輪作模式下細菌和真菌群落組成和結構的差異。其余土壤養分差異不顯著,可能由于試驗年份較短。然而,豆角-甜瓜輪作模式下土壤養分含量卻與西紅柿-甜瓜沒有顯著差異,這可能與甜瓜季養分投入量遠大于西紅柿季有關,且甜瓜季溫度高于西紅柿季,土壤養分礦化加劇,因此,甜瓜季高投入、高礦化使得土壤養分含量激增,掩蓋了上一季豆角減投的效果,具體機制還需進一步研究。
對于土壤細菌,結果表明變形菌門(Proteobacteria)在所有輪作模式是中所占比例最大,與以往研究一致[11,17,37],主要原因在于該類型細菌屬于典型富養型生物,其生存的r-策略使得它在碳源底物有效性高的設施環境中快速生長,成為優勢種群。然而,不同微生物對輪作模式改變及其導致的環境因子變化的響應不同,這取決于不同微生物的生理、生態特性差異。例如,發現亞硝化單胞菌科在西紅柿-豆角土壤中顯著增加,且與NO3-N含量有極顯著負相關(r=0.817,P<0.01)。以往研究也發現,減量施肥顯著提高亞硝化單胞菌科(屬)的相對豐度[38-39]。另外,亞硝化單胞菌科的相對豐度與pH值具有顯著正相關(r=0.667,P<0.05),這與普遍認為的低pH值會限制硝化細菌生長[40]結論一致。在3種輪作模式下,當季豆角種植(西紅柿-豆角)的水肥投入量明顯低于當季甜瓜種植模式,導致土壤pH值升高、NO3-N下降,因此,有助于該菌群生長。西紅柿-豆角輪作模式還顯著提高了己科河菌門的相對豐度。作為一種新型土壤細菌,該菌群具有多樣的生物次級代謝物基因,可能與抗藥性密切相關[41]。本研究發現,土壤NO3-N、總氮和pH值顯著影響該類微生物的相對豐度,表明此類細菌對土壤無機氮敏感性很高,并且有研究表明該菌群可以進行反硝化作用[42]。另外,相比于引入豆角的輪作模式(豆角-甜瓜和西紅柿-豆角),西紅柿-甜瓜土壤中Roseiflexaceae和生絲單胞菌科(Hyphomonadaceae)的相對豐度顯著增加。這兩類細菌多被發現于高溫、高鹽分環境中[43]及各種海洋環境中[44],而西紅柿-甜瓜土壤具有相對較高的EC值,可能是導致該種群富集的原因。
對于土壤真菌,子囊菌門(Ascomycota)在3種輪作模式中所占比例均最高,達到54.37%~69.50%,與Tian等[16]在環渤海區設施土壤調查結果一致[11]。研究指出設施土壤中子囊菌門豐度增加與其土壤磷含量高密切相關。在3種輪作模式中,擔子菌門(Basidiomycota)的相對豐度在西紅柿-豆角模式下顯著增加,其中銀耳綱(Tremellomycetes)和外擔菌綱(Exobasidiomycetes)的相對豐度增加程度較高。研究發現,這些真菌可能在抑制植株根部土傳病害方面發揮作用,是有效的生防菌[45]。然而,小不整球殼科、蟲草菌科和叢赤殼科(Nectriaceae)等具有致病的種群的相對豐度也顯著增加。 Bainard等[46]發現,豆科植物輪作頻數的增加會導致病原菌數量增加2~3倍,因此,提出應避免連續種植豆科作物,以防根系特異性病原菌累積。土壤中真菌優勢科的大部分屬于子囊菌門,在群落演替中處于演替前期,與接合菌、細菌等最先侵入枯枝落葉等基質形成優勢菌,分解代謝吸收大量糖和NPK等營養物質[47],因此,相對豐度與土壤NO3-N、Olsen-P、總氮、總鉀、有機碳、C/N等密切相關。以叢赤殼科為例,NO3-N、總氮、有機碳和pH值顯著影響其相對豐度,但具體影響機制仍需進一步研究。近年來,盡管高通量測序技術在各類型生態系統的廣泛應用為更好的理解土壤微生物多樣性及群落構建提供了豐碩的成果,但仍有相當一部分微生物在生態系統中功能并不十分清楚。
利用隨機森林模型,本研究預測出多種能夠顯著(P<0.05)影響3種輪作模式下群落構建差異的微生物種群。這些種群的相對豐度在3種輪作模式下有顯著差異,進而可能導致不同土壤中碳氮循環過程的改變。例如,紅微菌屬(Rhodomicrobium)是一種需微氧至厭氧的紫色光合細菌,具有極其多樣的碳源代謝能力,可代謝幾乎所有的發酵產物,包括CO2、小分子有機酸和CH4等,同時是一種自生固氮菌,具有固氮能力[48-49]。發現紅微菌屬的相對豐度在西紅柿-豆角土壤中顯著高于其余2種輪作模式,這暗示了共生固氮過程可能是豆角土壤中氮素供應的重要來源或補給。本研究還發現,氫孢菌屬(Hydrogenispora)的相對豐度在西紅柿-甜瓜土壤中顯著高于引入豆角的種植制度。該屬的細菌偏好利用糖類底物,如Hydrogenisporaethanolica能發酵葡萄糖、半乳糖、淀粉[50],這表明不同種植制度下土壤有機碳組分可能發生變化。另外,西紅柿-豆角土壤中輪枝孢屬(Verticillium)相對豐度顯著高于其余2種輪作模式,這類真菌會引起枯萎病,但是經過合理輪作,其相對豐度會顯著下降,可以看到上一季種植豆角的輪作模式豆角-甜瓜在這一季土壤中該類真菌相對豐度與西紅柿-甜瓜差異已經不顯著。
除了群落結構,本研究發現土壤真菌的豐富度對于輪作模式及其導致的土壤理化性質的改變更為敏感,而細菌多樣性的變化與土壤因子的相關性不顯著。3種輪作模式下土壤真菌Chao指數的變化與土壤pH值具有極顯著的負相關性(r=-0.767,P<0.01),這與以往研究結果一致[51-52]。目前,研究普遍承認pH值是影響真菌群落多樣性的主要化學性質[53-54],不過Bahram等[55]指出全球范圍內土壤真菌多樣性與pH值相關性較弱。這種差異很可能是由于生物及非生物因素改變,如植被變化、施肥變化等帶來的碳源底物、土壤理化性狀等環境因子改變綜合調控的結果,而非pH值的直接作用引起的變化。總體來說,本研究中輪作模式的改變僅影響到土壤微生物多樣性的某個方面(例如豐富度、均勻度),這可能是由于相應的施肥、灌水處理時間尚短。土壤微生物多樣性是其發揮生態系統功能的基礎[56],后續研究還需長期連續觀測,深入研究種植制度改變導致的生物及非生物因素變化對土壤微生物多樣性及群落特征的影響,從而揭示其影響設施土壤碳氮循環的機理。
設施農業高集約化生產方式導致土壤理化性質的改變,進而影響土壤微生物的多樣性、群落組成及功能。基于豆科作物養分減投的輪作制度調整技術相對于茄科-葫蘆科傳統輪作模式,經過2 a四季肥水管理顯著降低了土壤NO3-N含量并提高pH值。土壤環境的變化顯著改變了細菌和真菌的多樣性和群落組成,但不同微生物對輪作模式改變引起的環境因子變化的響應不同,主要取決于不同微生物的生理、生態特性差異。在所有土壤因子中,pH值和NO3-N對土壤微生物群落結構變化的影響最為顯著,并且當季作物種類(豆科/非豆科)對土壤微生物富集模式有顯著作用。此外,土壤細菌對輪作制度調整的響應比真菌敏感,未來應深入研究這種群落變化如何影響設施土壤碳氮循環關鍵過程。
