豬ATP6V0A4基因啟動子核心區的篩選與分析
2021-07-01 08:07:40李忠秋唐曉東張海峰張冬杰
華北農學報 2021年3期
汪 亮,李忠秋,唐曉東,張海峰,劉 娣,張冬杰
(1.黑龍江省農業科學院,黑龍江 哈爾濱 150086;2.農業農村部種養結合重點實驗室,黑龍江 哈爾濱 150086)
V-ATP酶最早是在酵母細胞的液泡膜上發現的,其主要功能是利用水解ATP產生的能量實現氫離子的跨膜運輸,在水解ATP、調節pH值和產生電勢梯度等方面具有重要作用[1]。在進化上高度保守,主要存在于不同生物的細胞膜或細胞器膜上。V-ATP酶是一種復雜的異聚體蛋白,包括2個結構復合體V1和V0,V1存在于膜表面,屬于催化區域,負責ATP的水解;V0包埋在膜內,屬于質子傳導區域,負責質子的轉運。V1由8個亞基組成(A、B、C、D、E、F、G、H),V0由6個亞基組成(a、c、c′、c″、d、e)[2]。隨著研究的深入,人們發現V-ATP酶除了可以轉運H+,調節細胞器內的pH值外,還參與許多其他的生物學過程。比如,在植物中,可以調節細胞膨壓,促進細胞伸長以及幫助植物適應干旱、鹽堿、低溫等非生物逆境脅迫[3]。在昆蟲中,可調節煙草天蛾、東亞飛蝗等的蛻皮活動,參與柑橘小實蠅雄蟲的生殖過程,在家蠶絲腺細胞中建立電勢差,利于細胞的胞吐作用以及后續絲膠蛋白的分泌[4]。
ATP6V0A4編碼V-ATP酶的a4亞基,被認為是遺傳性遠端腎小管性酸中毒的致病基因,該疾病因遠端腎小管泌氫障礙,使得可滴定酸及尿NH4+排出減少,從而導致一系列的臨床表現[5]。ATP6V0A4基因缺失小鼠到斷奶時,表現出嚴重的代謝性酸中毒,低血鉀和早起腎鈣化,聽力嚴重受損。除非堿化,否則它們會快速死亡[6]。民豬資源研究與利用課題組在前期研究中發現,民豬在遭受冷應激后,其背部皮下脂肪內的ATP6V0A4基因的轉錄水平會顯著升高(P<0.05)。目前,還未見該基因與脂肪代謝或體溫調節相關的報道,但其作為編碼V-ATP酶亞基的一個基因,參與細胞內的跨膜轉運質子活動,這種活動對于pH值的穩定和膜電位的產生至關重要,而膜電位是細胞代謝的驅動力[7]。為了后續探討ATP6V0A4基因在冷應激過程中的生物學功能,本研究對其啟動子區的核心序列進行了篩選和分析,從分子水平探討其可能存在的轉錄調控機制。
1 材料和方法
1.1 試驗材料
1頭12月齡的雌性民豬基因組DNA由黑龍江省家畜育種重點實驗室-20 ℃保存,內切酶KpnⅠ、Hind Ⅲ和pMD18-T Vector購自TaKaRa,雙熒光素酶報告載體購自Promega。
1.2 試驗方法
1.2.1 引物設計與PCR擴增 根據NCBI已有的豬ATP6V0A4基因組序列設計引物(GenBank登錄號:NC_010460),擴增-1 504~-1 bp序列(將起始密碼子“A”的位置定義為1),引物序列見表1。
以豬的基因組DNA為模板進行PCR擴增,反應體系為模板DNA 1 μL(50 ng/μL);2×PCR Mix 10 μL;上下游引物(10 μmol/L)各0.5 μL;ddH2O補齊至20 μL。反應條件為95 ℃預變性5 min;95 ℃變性30 s,60 ℃退火30 s,72 ℃延伸2 min,30個循環;72 ℃終延伸5 min;4 ℃保存。
1.2.2 克隆測序 參照TransGen Biotech膠回收試劑盒(EG101-02)說明書操作。將回收后的PCR產物連入pMD18-T載體,構建pMD18-T-ATP6V0A4質粒并轉入DH5α感受態細胞。應用限制性內切酶KpnⅠ和Hind Ⅲ進行雙酶切鑒定。雙酶切體系為KpnⅠ和Hind Ⅲ各0.5 μL(10 U/μL),10×Buffer 1.0 μL,質粒 4.0 μL;ddH2O 4.0 μL。反應條件為37 ℃酶切15 min。1.0%瓊脂糖凝膠電泳鑒定后選取陽性克隆菌落送至北京華大基因科技有限公司測序。應用DNAMAN軟件對測序結果進行分析。
1.2.3 熒光素酶報告基因質粒構建 將測序正確的克隆載體與pGL3-Basic空載體分別使用KpnⅠ和Hind Ⅲ雙酶切處理,回收目的片段后,將帶有黏性末端的目的片段與線性化的pGL3-Basic載體相連接,轉化至大腸桿菌DH5α細胞中,篩選陽性重組質粒,經菌液PCR、雙酶切、測序驗證后,提取無內毒素質粒(提取方法嚴格按照天根無內毒素質粒提取試劑盒說明書操作),-20 ℃保存備用。
1.2.4 轉染與熒光素酶活性檢測 將報告基因重組質粒與pRL-TK質粒共轉染至豬PK15細胞,轉染前一天以4×105~8×105細胞/皿的密度鋪至24孔板中,使第2天轉染時的細胞達到80%~90%匯合,轉染過程嚴格按照LipofectamineTM2000 (Invitrogen)說明書操作。每孔加入總質量為0.5 μg的質粒和2 μL轉染試劑,轉染后4~6 h換液,24 h后進行熒光素酶活性檢測。檢測時按照Promega產品說明書操作。
1.2.5ATP6V0A4基因啟動子核心區鑒定 以克隆測序后的1 504序列為參考序列,設計一系列截短型上游引物,5′端引入KpnⅠ酶切位點,它們共用1條下游引物1504A(表1),組成8對引物。以pMD18-T-ATP6V0A4質粒為模板,分別進行PCR擴增,回收目的片段后分別連入pMD18-T載體,測序驗證后將正確的質粒連入雙熒光素酶報告基因載體,轉染入細胞,進行雙熒光素酶活性檢測。
1.2.6ATP6V0A4基因啟動子區調控元件鑒定 為了保證預測的準確性,在3個不同網站對ATP6V0A4基因的啟動子區進行調控元件的預測(http://www.gene-regulation.com/pub/programs/alibaba2/index.html、http://jaspar.genereg.net、http://alggen.lsi.upc.es/cgi-bin/promo_v3/promo/promoinit.cgi?dirDB=TF_8.3和http://www.ifti. org)。

表1 擴增截短型啟動子所用引物Tab.1 Primers for amplification of truncated promoter
依據不同片段熒光素酶活性的測定結果,同時結合軟件的預測結果應用重疊延伸PCR對部分元件進行缺失突變,缺失突變用引物序列見表2。PCR反應分2輪進行,第1輪反應利用含有缺失序列的引物與原有引物進行配對擴增,在相互重疊的PCR產物末端突變目的序列,第2輪反應以第1輪反應獲得的2個片段為模板,利用原有引物進行拼接擴增,將缺失突變引入產物內部。質粒的構建與熒光素酶活性的測定同上。
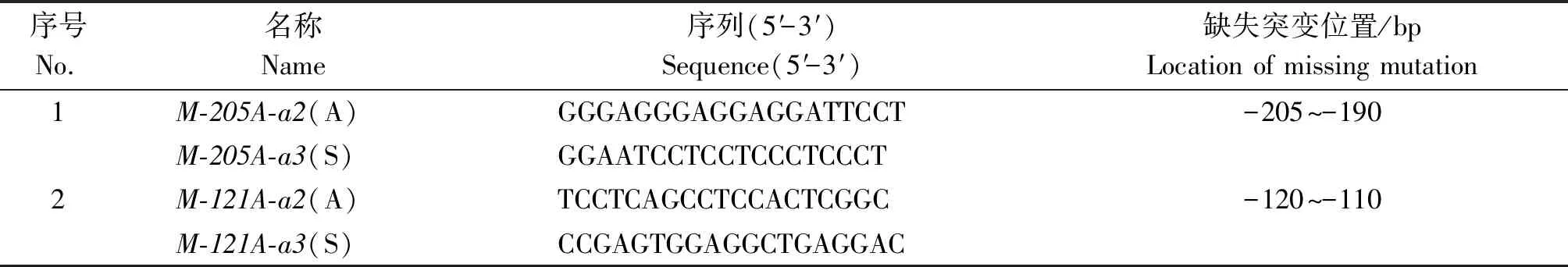
表2 突變調控元件結合位點所用引物信息Tab.2 Primer information for binding sites of mutation regulatory elements
2 結果與 分析
2.1 ATP6V0A4基因啟動子區的擴增與鑒定
利用引物1504S和1504A進行PCR擴增,瓊脂糖凝膠電泳檢測后在1 500 bp左右發現清晰明亮的條帶(圖1),經雙酶切鑒定及測序驗證后,證明所擴增的片段為豬的ATP6V0A4基因啟動子區。
2.2 熒光素酶報告基因質粒構建
利用限制性內切酶KpnⅠ和Hind Ⅲ對構建成功的克隆載體及pGL3-Basic空載體進行雙酶切處理,純化目的片段及線性化的載體。經連接、轉化后得到重組質粒。對重組質粒進行雙酶切、測序鑒定。結果表明,成功地將NoxⅠ基因啟動子區連入了pGL3-Basic載體。
2.3 熒光素酶活性檢測
將提取的pGL3-Basic-1504無內毒素質粒轉染至PK15細胞中,24 h后檢測熒光素酶活性。結果顯示-1504~-1 bp(圖2)片段含有啟動子活性。
2.4 ATP6V0A4基因啟動子區系列截短序列載體的構建
以1504片段為模板,利用8條截短序列的上游引物與1504A配對,共擴增出8條長度不同的ATP6V0A4啟動子區片段。將PCR擴增出的目的片段與pMD18-T載體相連,經轉化,挑取單克隆菌落擴大培養后提取質粒,經雙酶切(圖3)、測序驗證后,分別連入pGL3-Basic載體,構建8個報告基因重組載體,分別為pGL3-Basic-1124、pGL3-Basic-813、pGL3-Basic-581、pGL3-Basic-468、pGL3-Basic-368、pGL3-Basic-265、pGL3-Basic-185和pGL3-Basic-107(數字代表插入片段的長度)。
2.5 熒光素酶活性分析
第1輪熒光素酶活性分析主要是從1 504 bp片段至265 bp片段,區間間隔大約為300 bp長度。因此,共檢測了4個片段的熒光素酶活性,以空載體pGL3-Basic作為陰性對照,以pGL3-Basic-1504作為陽性對照,具體檢測結果見圖4。由圖可知,1504截短至1124、813、581后,啟動子活性無顯著變化,說明-1 504--581 bp區域內沒有重要的轉錄調控位點。將581位置截短至265后,啟動子活性顯著升高(P<0.05),說明-581--265 bp區域內存在負調控元件,-265--1 bp區域內存在正調控元件。
第2輪缺失以發生顯著變化的區間-581--1 bp為基礎,將此區間進一步分割,結果發現-265--185 bp區域內存在正調控元件(圖5)。
2.6 ATP6V0A4基因轉錄調控元件的預測與鑒定
2.6.1 生物信息學預測結果 根據前期雙熒光素酶測定結果,同時綜合3個在線轉錄因子預測軟件的分析結果,在-205--190 bp區間預測到8個轉錄因子結合位點,分別為E2F3、SP2、EGR1、E2F6、CTCFL、E2F1、SP1和ETF;在-120--110 bp區間預測到4個轉錄因子結合位點,分別為HNF4G、HNF4A、NR2F1和SP1。
2.6.2 轉錄因子結合位點的驗證 利用重疊延伸PCR法對預測的ATP6V0A4基因潛在的轉錄因子結合位點-205--190 bp和-120--110 bp進行缺失突變,經酶切測序驗證后,與pGL3-Basic載體相連構成轉錄因子結合位點缺失的報告基因質粒pGL3-Basic-205和pGL3-Basic-121(數字表示突變位點)。
以pGL3-Basic-265和pGL3-Basic-185為參照,將構建的缺失突變體進行雙熒光素酶活性檢測。結果顯示缺失-205--190 bp區域,雙熒光素酶活性顯著降低,說明該區域存在正調控轉錄因子;缺失-121--110 bp區域,雙熒光素酶活性不降反而有上升趨勢,說明這個區域可能含有轉錄負調控轉錄因子(圖6)。
3 討論
目前,關于ATP6V0A4基因的研究幾乎全部集中在人類的原發性遠端腎小管酸中毒(dRTA)疾病上,dRTA是一種罕見的遺傳性疾病,其特征是遠端腎單位的尿液酸化過程受損,導致堿性尿的產生[8]。其致病機理還不清楚,但ATP6V0A4的突變與dRTA顯著相關[9]。此外,ATP6V0A4還與乳腺癌的內臟轉移相關,發生內臟轉移的乳腺癌患者該基因顯著高表達[10]。ATP6V0A4為組織特異性表達基因,在腎臟、內耳、嗅覺上皮和雄性生殖道的質膜、雌性的子宮、胚胎內臟卵黃囊等組織表達[11],說明該基因具有廣泛的生物學功能。
河北省植物生理及分子病理學重點實驗室在前期研究中發現,民豬在遭受冷應激刺激后,其背部脂肪內的ATP6V0A4基因轉錄水平顯著上升,上升倍數可達8.42倍,但在正常情況下,脂肪型的民豬與瘦肉型的大白豬相比,大白豬背脂內的ATP6V0A4基因轉錄水平比民豬的高8.83倍[12]。據此,筆者推測該基因可能與豬的脂肪代謝相關,并且受冷應激誘導,但目前還未見相關的研究報道。只知其最后的聚合產物V-ATP酶的組裝和活性會受到葡萄糖饑餓的誘導[13],而冷應激也會通過胰島素依賴途徑增強葡萄糖氧化,進而刺激外周組織中的葡萄糖的攝取[14]。為了開展ATP6V0A4基因后續的功能分析,本研究在分子水平上對該基因啟動子區的正調控轉錄因子結合位點進行了篩選與分析,發現-581--265 bp區域內存在負調控元件,-205--190 bp區域內存在正調控元件,利用在線軟件預測后發現-205--190 bp 區域內存在E2F3、SP2、EGR1、E2F6、CTCFL、E2F1、SP1和ETF共計8個轉錄因子的識別序列。
E2F3、E2F6和E2F1都屬于E2F轉錄因子家族,它們通過調節控制細胞DNA合成及與細胞增殖有關的基因表達而起作用,同時本身的活性也受到其他在細胞周期進程中有重要作用的蛋白所調控[15]。如高遷移率族蛋白B1(HMGB1)可通過上調E2F3促進卵巢癌細胞增殖和侵襲能力[16]。SP1和SP2同屬于Sp樣轉錄因子家族,該家族共有8個成員,廣泛參與細胞增殖、分化、凋亡、胚胎發育、腫瘤發生等多種生理病理過程,調控作用廣泛[17]。而EGR1、CTCFL和ETF這3個轉錄因子的研究報道較少,多集中在癌癥的相關研究上[18-20]。本研究中,確定了正、負調控元件的結合區域,但具體哪個轉錄因子會結合到ATP6V0A4基因的啟動子區正調控/負調控該基因的轉錄,還需要后續的定點缺失試驗驗證。
