苔草屬植物遺傳多樣性研究進展
2021-07-05 02:21:08劉凌云范希峰岳躍森常智慧武菊英
草地學報 2021年6期
劉凌云, 范希峰, 滕 珂, 岳躍森, 常智慧, 武菊英*
(1.北京林業大學草業與草原學院, 北京 100083; 2.北京市農林科學院北京草業與環境研究發展中心, 北京 100097)
苔草屬(CarexL.)是莎草科(Cyperaceae)多年生草本植物,多為自花授粉、雌雄同株。其分布范圍廣,世界約有2 000種,在中國有近500種,是莎草科中的最大屬之一,僅次于大戟屬和胡椒屬[1-5]。苔草具有自繁能力強、耐蔭、耐旱、耐瘠薄等優點,具備一定的觀賞價值,不但可以作為優質的草坪地被植物,還能夠作為優良的牧草[6-7]。苔草屬植物的遺傳多樣性是長期進化的產物,是其生存和進化的前提[8]。對于植物育種工作來說,研究遺傳多樣性具有重要指導意義[9]。目前苔草的育種工作也一直是研究的重點,本文主要介紹苔草屬植物遺傳多樣性方面的研究進展,以期為苔草屬植物的育種工作和進一步開發利用奠定基礎。
1 苔草屬植物自然分布及分類學現狀
苔草屬植物是分布最廣泛和具有重要生態意義的植物之一。從分布范圍看,莎草科植物遍布世界各地,是植物中少有的遍及亞洲、歐洲和美洲的世界性植物[10],其適應力強,從沙漠到熱帶雨林甚至北極等各種氣候類型條件均可生存[11]。就種類、擴繁能力和生物量來說,苔草在許多區域都是建群的優勢種。《中國植物志》收錄近500種苔草資源,分布于全國各省區(圖1)。苔草屬可分為二柱苔草亞屬、復序苔草亞屬和苔草亞屬3個亞屬[12-13]。結合《中國植物志》的數據,我們發現二柱苔草亞屬分布較為廣泛,復序苔草亞屬主要分布在長江以南地區。苔草亞屬全國范圍內均有分布,四川、云南、浙江、臺灣等省的分布密度較為集中。我國苔草屬資源的自然分布情況表明苔草屬可較好地適應我國的不同氣候類型,是一類具有重要開發潛力的資源。
近年來,國內外也不斷發現苔草屬新物種,包括東亞、北美、歐洲地區,以及我國的青藏高原和海南地區等[14-18]。在過去5年里,苔草近源種及種間關系的研究豐富,而缺乏對其系統進化的研究。由于其形態復雜、高變異以及易雜交等特點,苔草純種和雜種的鑒定比較困難。遺傳變異與多樣性也隨之提高,分類比較困難[3,10,19-20],使苔草成為世界上植物譜系最多樣化的植物[21]。對部分地區苔草進行的資源調查與多樣性分析證實其群落的物種多樣性指數越大,則均勻度指數、物種豐富度指數越大,并且優勢度指數相應越小[22-23]。盡管對苔草研究的深度和廣度都在增加,但隨著苔草屬的不斷進化與重組,對苔草屬內的植物分類仍是一大難題。
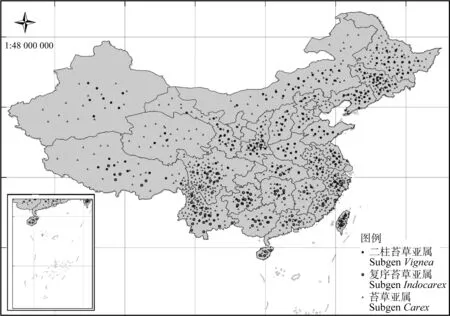
圖1 中國苔草屬資源分布情況Fig.1 The geographical distribution of Carex Linn. in China注:地圖審圖號為GS(2014)5260Note:The map verification number is GS (2014) 5260
2 苔草屬植物遺傳多樣性研究概況
2.1 形態多樣性
苔草屬植物眾多,形態各異(圖2),評價苔草的形態多樣性主要根據色澤、株高、冠幅、葉片長度、葉片寬度、葉片質地等指標[24-25]。苔草屬于須根、擴葉型草種,其莖最高能夠達到120 cm,粗度最高能夠達到4 mm。苔草葉片較為粗糙具有韌性,花序呈圓柱狀[26]。種子成熟后為長方形,花朵中雄蕊有3個。苔草屬于自花授粉植物,繁殖力強,花粉傳播廣是苔草擴展能力強的重要原因[27-28]。
形態多樣性一定程度上決定了不同的苔草屬植物的分布狀態。梁芳等[29]對北京地區苔草屬植物的研究表明,華北苔草(C.hancockiana)、溪水苔草(C.forficula)等自身株高較高的種類易倒伏,從而形成種群小、叢狀分散的分布狀態;而生長均一性好,叢高、葉寬相仿的矮叢苔草和披針葉苔草則能形成較大種群的片狀分布。梁芳等[30]進一步對20種北京地區引種苔草進行形態學的研究,發現形態高度相似的披針葉苔草、早春苔草和矮叢苔草,在開花結實期間有較大差別,據此可以對這幾種苔草進行區分。不足的是,該研究并未通過形態指標對苔草進行遺傳多樣性的分析。劉凌云等[31]對12份苔草屬植物材料的表型性狀測量得到的數據進行聚類分析,結果證明地理來源與形態多樣性無直接相關性。

圖2 北京市農林科學院保存苔草的部分照片Fig.2 Some Carex materials preserved in Beijing Academy of Agriculture and Forestry Sciences注:從左至右依次是矮叢苔草、橫果苔草、尖嘴苔草、狼牙棒苔草、藍苔草、異鱗苔草、棕紅苔草、白穎苔草、棕櫚葉苔草、澇峪苔草、卵穗苔草、英國苔草Note:From left to right,it’s C. callitrichos,C. transversa,C. leiorhyncha,C. grayi,C. flacca,C. heterolepis,C. buchananii,C. duriuscula,C. muskingumensis,C. giraldiana,C. ovatispiculata,C. england
新品種多是由于表型變異而產生的,而表型變異是由遺傳物質決定的。Schmidt等[32]研究了C.flava,C.viridula以及它們的雜交種C.subviridula在相同條件下的生存、生長、繁殖和形態特征,發現遺傳變異以及基因型與環境的相互作用都是密切相關的,并且常見的苔草屬種類比不常見的種類具有更強的適應性。在越南地區發現的棕葉苔草(C.kucyniakii)是3個物種的復合體,并發生了顯著的形態變異,特征是無毛的周膜、鱗片和花序。而都具有短柔毛的C.geographica和C.thinii可以根據雌蕊和雄蕊鱗片形態和花藥長度的差異進行區分[33]。在海南新發現的尖峰嶺苔草(C.jianfengensis)具約1 cm長的鞘,花序有4個穗,果實呈卵形,是與遵義苔草(C.zunyiensis)相似但是不同的新種類[14]。
對形態水平的研究有助于植物生物學上的識別,但外觀差異不明顯使得人們無法從肉眼直接區分苔草的種類,雖可以通過開花結實期的果囊形態等進行分辨,但苔草結實期短,此種方法并非快捷有效,這造成了苔草分類及鑒別困難。
2.2 細胞多樣性
苔草細胞學遺傳多樣性的研究主要集中在葉片細胞、果皮細胞以及種皮細胞的研究方面。馬萬里等[34]通過苔草葉片的硅細胞硅體突起方向差異特征對苔草屬進行種間的分類。除了葉片細胞的觀察,張樹仁等[35]通過對果實的表皮細胞進行了苔草的遺傳多樣性研究。通過電子顯微鏡對15種苔草屬植物的果皮微形態進行特征分析,將15種苔草屬植物進行了區分,證明果皮的微形態特征在種內很穩定,種間存在不同程度的差異。這與馬萬里等[36]研究發現的苔草種內果實的表皮細胞的形態差異較小,亞屬之間存在交叉與滲透的現象相一致。丁雪珍[27]對苔草葉片表皮微狀態的研究證明苔草種間可以通過脈區間長細胞的形態、刺毛的有無、氣孔的行數、保衛細胞的形態等確定苔草屬的屬下分組,對苔草屬的分類有一定的意義。
明確苔草屬植物染色體的數量是了解其核型特征和染色體進化在物種多樣性中作用的基礎。研究證明,苔草屬植物染色體數目的非整倍性特點造成其染色體數目的多態性。Tanaka[37]對13個種的不同組合進行雜交,獲得了19個雜交種,并對這32份材料進行了染色體數目統計,發現了33種單倍體染色體數目類型,包括6,8,9,12,13,15,16,17,18,19,23~43,54,56,同時也有非整倍性和多倍體類型出現。多倍體類型的C.siderosticta苔草具有76條染色體,遵循倍數規律,證明苔草屬中具有一定程度的天然雜交種。WhikusF等[38]對苔草亞屬Ovales包含的8種苔草(C.bebbii,C.crawfordii,C.integra,C.microptera,C.pachystachya,C.praticola,C.preslii和C.subfusca)進行了染色體計數,發現了染色體改變的2種形式分別是結構重排和非整倍性的數目變化,這對苔草屬植物的進化與發育、苔草屬植物分類等具有重要指導意義。Hipp等[39]研究了北美東部和西部的Ovales,計算表明北美洲東部苔草種質的二倍體染色體計數平均值為68.38,北美西部的二倍體染色體平均計數為79.23。
苔草中染色體數目與基因組大小之間存在一定相關性。Escudero等[40]從Carexgr.laevigata的20個種群中選取了26個已知染色體數的個體,并通過流式細胞術評估了基因組的大小,發現苔草屬植物基因組大小和染色體數目之間存在顯著的正相關關系,揭示了核型進化在苔草屬植物物種多樣性中的重要作用。Liu等[41]通過染色體的數目對苔草的亞屬進行了分類,發現二柱苔草亞屬在切片水平上顯示相似的染色體計數,難以進行鑒定,而亞屬下的C.repens染色體數目明顯不同,極可能為雜交產物,這說明染色體數目與分類學相結合的方法為苔草屬植物的分類提供新的方向。
2.3 分子遺傳多樣性
除了細胞學和形態學標記,現代分子生物學技術也在苔草研究中不斷應用和發展。Bogucka-Kocka等[42]嘗試通過測定不同酚酸類化合物的含量來對苔草進行劃分,結果發現化合物的含量大多偏高,并不能作為苔草分類學依據。相比于細胞學和形態學標記,分子生物技術更快速高效和準確。在分子水平上最廣泛應用于遺傳多樣性分析的是微衛星標記,包括以核糖體DNA(nrDNA)為基礎的內部轉錄(Internal transcribed spacer,ITS)和外部轉錄(External transcribed spacer,ETS)的方法[43-46];葉綠體DNA(cpDNA)為基礎的matK條形碼技術;通用引物ISSR標記法[47];轉錄組測序EST-SSR分子標記法;單核苷酸多態性(Single nucleotide polymorphism,SNP)基因芯片技術等(表1)。這些方法是研究苔草的系統進化與遺傳多樣性的有力工具,極大地推動了苔草遺傳多樣性的研究進展。
2.3.1ETS,ITS,matK條形碼技術 ETS,ITS和matK都屬于通過擴增基因序列達到鑒別植物或微生物的方法,可用于區分親緣關系非常近的種。不同的是,ITS序列是指rDNA基因中的16S rDNA和23S rDNA基因間隔序,而matK條形碼技術是通過擴增葉綠體基因實現的[45,48]。Starr等[4]通過克隆測序獲得的matK條形碼對3個苔草屬亞族進行區分,結果47%的苔草屬試驗材料可被區分,而且證明可以通過matK條形碼技術發現新物種,但是無論是通過單位點和多位點條形碼組合都無法識別超過60%的苔草物種。為提高鑒別效率,Míguez等[49]通過開發ITS,ETS,matK標記組合的方式,研究了氣候對苔草分布的影響,為苔草的系統地理學提供基礎。
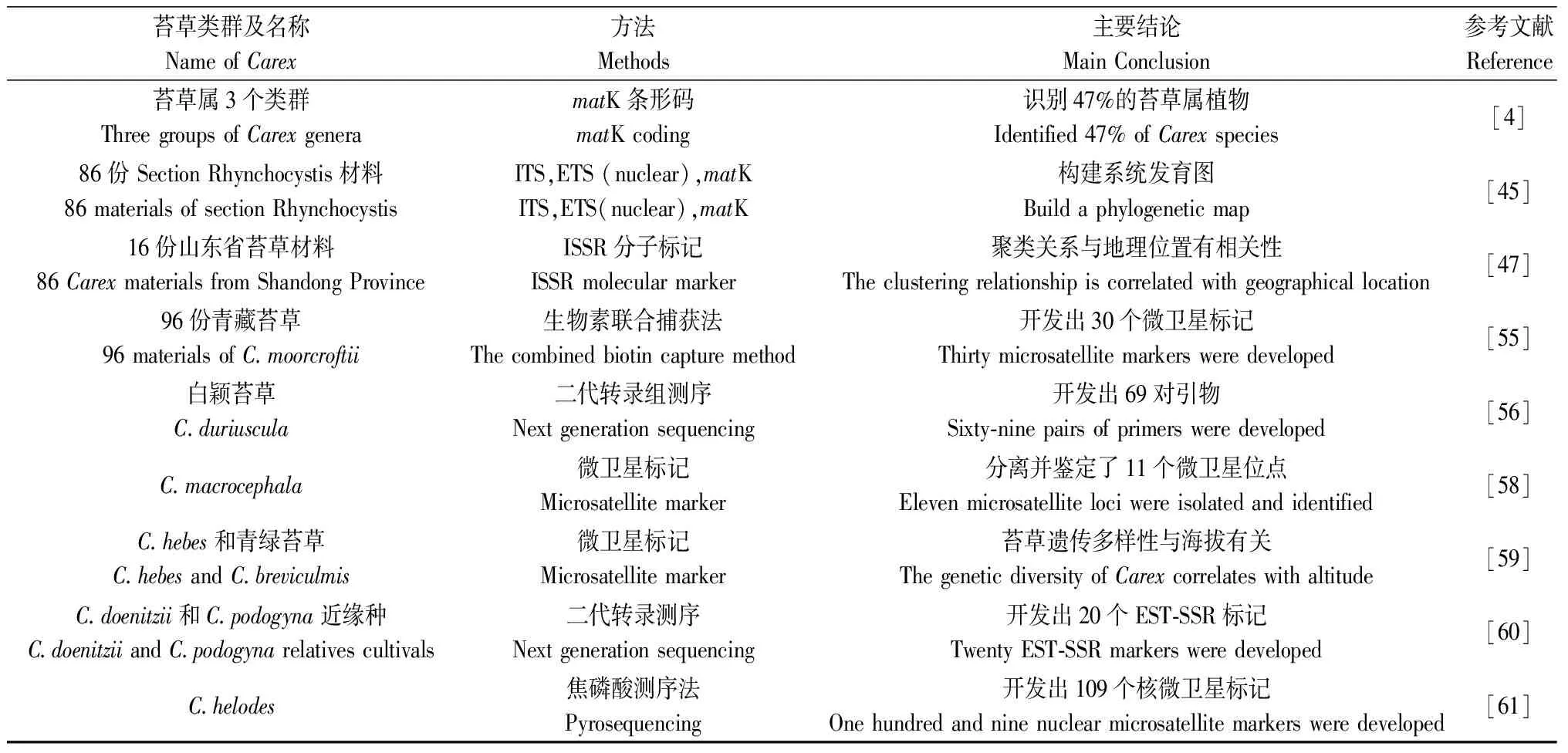
表1 近十年對苔草屬植物遺傳多樣性研究的方法及主要結論Table 1 Methods and main conclusions of the research on genetic diversity of Carex Linn. in the last decade
2.3.2ISSR通用分子標記引物 簡單重復序列間區標記(Inter-simple sequence repeat,ISSR),不需要預先對植物的轉錄組進行測序,而是通過生物基因組中出現頻率較高的簡單重復序列設計引物。包括1~4個堿基組成的串聯重復序列及非重復的錨定堿基組成,在PCR作用下通過與基因組DNA中串聯重復序列的3′或5′末端結合,凝膠電泳顯示多態性[50-51]。為了提高鑒別效率Ning等[47]用簡單序列重復標記研究16份山東苔草屬材料的遺傳多樣性,并且根據非加權平均法聚類(Unweighted pair-group method with arithmetic mean,UPGMA)樹狀圖發現地域分布與苔草遺傳多樣性具有相關性。不足的是ISSR是植物的通用引物,可能造成結果不準確,開發苔草本屬植物的特異標記更有利于苔草遺傳關系的研究。
2.3.3SSR分子標記 簡單重復序列(Simple sequence repeat,SSR),也叫微衛星(Microsatellites)序列,是指可以根據保守的單拷貝序列兩端的序列設計一對特異引物,利用PCR技術將其擴增出來并通過電泳獲得多態性的一種標記方法[52-54]。其優點是可以直接分析遺傳物質,并能快速有效地評估種質資源的遺傳多樣性,而不受環境條件和基因表達的影響,在苔草中也廣泛應用。
苔草屬植物的微衛星標記研究主要集中在親緣多態性關系的研究上,通過開發、分離及鑒定對苔草屬植物進行遺傳關系的分析。Liu等[55]從青藏苔草(C.moorcroftii)的96個個體中開發分離并鑒定出30個微衛星引物,其中22對引物顯示多態性,每個位點有2到11個等位基因,觀測雜合度在0.00~1.00之間,期望雜合度在0.00~0.78之間。Li等[56]通過二代轉錄組RNA測序在白穎苔草中成功開發出144對引物,并且對2種不同基因型的白穎苔草‘綠坪1號’品種進行擴增,發現69對引物表現多態性,為分子標記鑒定不同基因型的品種提供了新思路。Ohsako和Yamane[57]從C.kobomugi中開發了14個微衛星標記,研究了C.kobomugi的24個分型和6個群體中的8個不同個體的多態性。結果證明每個基因座有2 ~ 8個等位基因,期望雜合度在0.04 ~ 0.72之間。基于該研究結果,Kim等[58]從與C.kobomugi可形成復合體的C.macrocephala里分離了11個微衛星標記,證明雖然基因位點的多態性高,但群體的觀測雜合性較低,同時開發出的標記在C.kobomugi中也能進行擴增證明了引物的可轉移性。M’Baya等[59]利用C.kobomugi開發出的微衛星標記研究了C.hebes和青綠苔草(C.breviculmis)的遺傳多樣性與海拔的關系,認為2種苔草植物都具有高水平的遺傳變異,但克隆度低、近交系數高、隔離程度高。Nagasawa等[60]通過二代轉錄組數據開發了C.angustisquama的EST-SSR標記,分析C.angustisquama及其近源種C.doenitzii和C.podogyna的多態性關系,其中期望雜合度在0.00 ~ 0.58之間,并發現種群的進化關系可導致較低的多態性,這與Arroyo等[61]對分布在葡萄牙南部和西班牙西南部的苔草C.helodes開發微衛星的研究結果一致,其通過開發的91個多態性位點和18個單態性位點發現基因的變異水平較低。Liu等[62]對同為苔草屬的79份苔草材料進行SSR分子標記分析遺傳多樣性,證明了所選的苔草屬植物均存在跨種擴增的現象,這與Gillespie等[11]結果一致。
對苔草屬植物的微衛星標記研究證明,苔草屬植物具有較高水平的遺傳多樣性,通過有利等位基因的交流可以減少近交系的衰退,有助于促進適應性差異,同時標記的通用說明微衛星標記在親緣物種間具有可轉移性,這對研究苔草屬植物不同種間的遺傳多樣性具有重要意義。
3 苔草屬植物遺傳多樣性研究的應用
3.1 遺傳進化關系
苔草的遺傳多樣性研究可以對苔草進化史以及基因與地域的關系的研究提供指導。Starr等[5]和GlobalCarexGroup通過研究苔草的遺傳多樣性研究其分類譜系,結果證明苔草族植物被分為5大譜系(the CoreCarex,Schoenoxiphium,CoreUnispicate,Vignea和SiderostictaeClades)(圖3),并且證明苔草屬植物在各譜系中均有分布,且許多早期分化的部落譜系都起源于東亞[3,63]。在地理位置與基因的關系的研究上Vellend等[64]認為苔草C.rariflora基因變異與地理因素有關系,并且因為地理位置的因素使亞洲苔草基因變異率明顯低于北美苔草。Benítez等[65]的研究也印證了這一觀點,通過擴增片段長度多態性(Amplified fragment length polymorphism,AFLP)方法對苔草進行系統發育與地理位置和基因流動的關系進行研究,發現地理因素阻礙了基因的流動,對研究苔草的進化史提供了有力依據。

圖3 基于分子發育系統的莎草科系統發育樹(根據Global Carex Group,2015改繪)Fig.3 Generalized phylogenetic tree of Cyperaceae based on molecular phylogenetic studies(Redrew according to Global Carex Group,2015)注:實線代表的是在大多數研究都支持的關系,虛線則是常見但在研究中不一致的關系;正方形框中代表每個分支又單獨分出的2~4個亞群Note:Full lines show relationships that are supported by all or most studies. Dotted branches show relationships that are frequently seen but more inconsistent among studies. The square box represents two to four subgroups that each branch divides into separately
3.2 生產應用
苔草在生產上應用廣泛,包括園林綠地、飼料、藥用、水土保持等領域[66]。苔草屬部分種類具有低矮、地下根莖發達和覆蓋度好的特點,具有開發成草坪的潛力,有助于緩解我國草坪草種過度依賴進口的現狀[67]。不同類型的苔草在園林綠化中承擔的角色各不相同,如植株低矮且耐踐踏的可作為草坪應用;而冠幅及高度都較大的可作為觀賞草應用[24,68];苔草不僅可以應用在園林綠化,在青藏高原地區也可作為牧草,是一種適口性好、營養豐富的牲畜飼料[69-71];同時部分苔草種類對改善鹽堿地,治理土壤沙化也有著重要作用[72];在藥用方面,不同季節采收的翼果苔草(Carexneurocarpa)有抗病毒作用,制備的白穎苔草片劑也廣泛應用于醫藥領域[73-74];Kamel等[75]的研究證明苔草中的多酚提取物具有血管松弛作用和抑制哺乳動物精氨酸酶的能力,對改善血管功能具有積極作用。
4 苔草屬植物遺傳多樣性研究存在的問題
研究苔草屬植物遺傳多樣性有助于新品種的培育,現代園林綠化逐漸以生態環保、低維護為優先條件,在這樣的環境下,苔草有巨大的開發潛力。目前我國苔草資源收集數量少、重點不夠突出、范圍不夠廣泛,商品化的苔草品種較為缺乏(表2)。雖已在國內外園林中開始應用,但面積不大。其次苔草的遺傳背景不清晰、親緣關系不明確、種質資源混亂等問題制約了苔草資源的引種、雜交親本合理選配以及基因工程育種工作,阻礙了苔草的進一步推廣應用。
通過在形態多樣性、細胞多樣性以及分子多樣性水平上對苔草屬的遺傳多樣性進行研究,有助于深入了解苔草屬植物,對其分類、遺傳育種以及新品種的培育都具有重要意義。目前基于轉錄組測序開發分子標記已在苔草上面廣泛應用,為苔草的分子改良、品種鑒定、指紋圖譜的構建等奠定基礎。以DNA芯片技術為核心的核苷酸多態性SNP分子標記依賴于完善的基因組信息,隨著DNA測序技術的飛躍式發展,越來越多的植物已經進行了完整的基因組測序,為基因組學研究提供了豐富的數據[76]。基因組測序對被子植物的進化歷史研究具有重要意義,國外對選育草類植物新品種的研究一直處于領先地位,且正在開展草類植物農藝性狀的選擇,我國的草類植物種質資源豐富但未能充分開發利用,近年來陸續完成了紫花苜蓿(Medicagosativa) ‘中苜一號’[77]、鴨茅草(DactylisglomerataL.)[78]、象草(Cenchruspurpureus)[79]、小嵩草(Kobresialittledalei)[80]的全基因組測定,為其性狀的改良、分子育種奠定基礎。而苔草屬植物基因組的信息并未見報道,由于缺少基因組和重要性狀遺傳信息,其遺傳與育種工作面臨極大挑戰,對其優勢性狀也僅集中在某幾個基因的克隆研究,因此十分必要對苔草的基因組信息進行開發與利用。
此外遺傳多樣性分析在苔草上的應用也有所欠缺,由于苔草種類眾多,材料復雜且不易分類,單純依靠形態指標不能對其進行準確區分,DNA指紋圖譜則成為品種鑒定的一種更加便捷有效的方式。很多植物已經成功進行了指紋圖譜的繪制,包括假儉草(Eremochloaophiuroides)[9]、老芒麥(Elymussibiricus)[81]、苜蓿(Medicagosativa)[82-83]、大棗(Jujube)等[84]。其中Zhao等[81]利用不同的條帶型進行了雙親和雄性的鑒定,通過9對EST-SSR引物對不同老芒麥的品種進行了區分。閔學陽等[82]通過一對引物一次可以區分7個苜蓿品種。李懷志等[85]利用8對引物建立了茄子(Solanummelongena)的指紋圖譜,可以把‘閩長茄’與其他茄子品種完全區分開,‘敦和茄’和‘蘇大青茄’同其他茄子品種區分開來。王杰等[86]建立了20對引物的指紋圖譜,最少利用2對SSR引物組合可鑒別17份國審蘇丹草(SorghumSudanense)和高丹草(Sorghumbicolor×SorghumSudanense)。這都證明了分子標記能很好的用于建立指紋圖譜并進行品種的鑒定與分類,同時有助于推動苔草的分子標記輔助育種以及親緣物種鑒定的工作,包括鑒定半同源的父本親本和選擇雜交親本以產生優良的雜種,為之后的研究提供基礎,推動苔草在現代園林綠化的應用,而目前對苔草指紋圖譜繪制的工作十分匱乏。
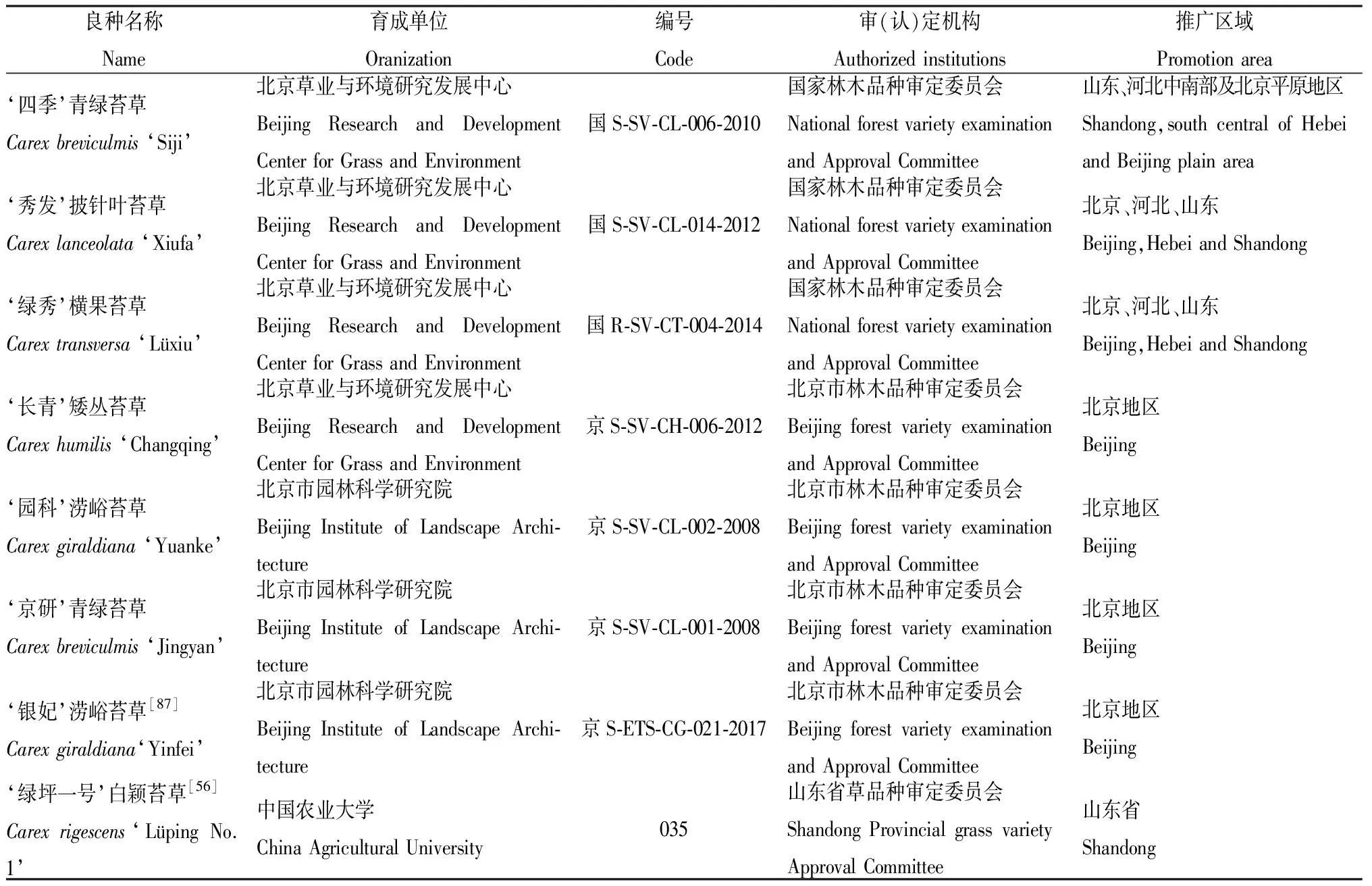
表2 我國自主培育苔草新品種統計Table 2 Statistics of the new cultivars in China
5 結論與展望
苔草屬植物作為我國的一種重要的草坪草種質資源,雖然資源豐富,且在生態修復、園林綠化方面都有巨大的應用潛力,但是現階段我們對苔草研究及利用都十分有限。因此,未來對苔草的研究方向應在苔草遺傳多樣性的基礎上,探討苔草的進化過程和優良性狀的形成機制,以期望培育園林適用的苔草新種質。同時對苔草屬植物的遺傳多樣性研究要更加深入廣泛,建立其核心種質資源庫、繪制指紋圖譜以提高品種鑒定鑒別效率、進行基因組測序等都是重要方向。最終我們通過研究苔草的遺傳多樣性作為苔草屬品種改良的基礎,推動培育新品種,發展有自主知識產權的草坪綠化草種,改變“泊來草”、“進口草”的局面,使苔草在我國園林綠化中發揮更大的作用。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
