山黧豆根腐病病原菌亞洲鐮孢菌的分離鑒定
2021-07-05 02:22:46邢會琴王春明金社林周天旺
草地學報 2021年6期
關鍵詞:研究
邢會琴, 王春明, 金社林, 周天旺, 郭 成
(1. 河西學院農業與生態工程學院, 甘肅 張掖 734000;2. 甘肅省農業科學院植物保護研究所, 甘肅 蘭州 730070)
山黧豆(Lathyrussativus)作為豆科(Leguminosae)山黧豆屬(Lathyrus)一年生作物,廣泛種植于世界各地[1]。其營養豐富,含有蛋白質、氨基酸、淀粉、碳水化合物和脂肪等[2],為飼肥兼用作物[3],主要分布在干旱、半干旱地區[4]。目前常被用作綠肥作物在生產上推廣應用。山黧豆具有抵御各種逆境的優良性狀,對環境的適應性強,能在干旱、貧瘠、鹽堿和水澇等惡劣條件下生長[5-7],且山黧豆作為豆科作物,本身具有很強的固氮能力[8],在其生長過程中,形成的根瘤可以固定空氣中存在的氮素,固定的氮素一部分用于山黧豆自身生長所需,另外一部分增加了土壤中的氮素含量,具有改良土壤、培肥地力的功效。山黧豆是可持續農業發展農業模式中的重要作物[9],尤其適宜種植于我國西北地區干旱、高寒、貧瘠的山區及丘陵溝壑區[10]。研究表明,長期種植林生山黧豆的人工草地土壤鹽分含量明顯降低,土壤pH值顯著降低[11]。近年來,在山黧豆種植過程中根腐病發生率日益增加,本研究于2018年8月在四川省南充市調查時發現,山黧豆根腐病發生嚴重,通過采集病樣、分離純化共獲得39株單孢分離物,并采用貼菌餅法和拌菌土法接種山黧豆對其致病性進行研究,結果發現其中3株分離物對山黧豆具有很強的致病性。為了確定這3株分離物的分類地位,本研究通過形態學和翻譯延伸因子(Translation elongation factor 1-agene,EF-1α)序列分析對其進行鑒定,旨在為山黧豆根腐病的綜合防控提供理論依據。
1 材料與方法
1.1 病樣采集及癥狀觀察
2018年8月從四川省南充市采集感病的山黧豆植株,觀察記錄發病癥狀,并帶回實驗室進行分離鑒定。
1.2 病原菌分離純化
采用組織分離法[12],將發生根腐病的山黧豆根部清洗干凈,植株的根頸部作為分離材料,切取發病部位根段浸泡在70%乙醇中進行表面消毒30~60 s后,無菌水沖洗3次,用滅過菌的剪刀將消過毒的發病根段剪成2 mm×5 mm大小的組織塊,用滅菌濾紙吸干組織塊表面的水分,置于PDA(DifcoTMPotato Dextrose Agar 39 g、蒸餾水1 000 mL)平板上,每皿放5個組織塊,將平皿倒置于培養箱中,25℃恒溫黑暗培養。待接種組織塊邊緣長出菌絲后,根據顏色和形態的差異輕輕挑取菌絲接種到PDA平板上進行初步純化后,采用稀釋法[13]進行單孢分離,并將分離物接種于PDA斜面上培養好后4℃保存備用。
1.3 致病性測定
貼菌餅法:將供試菌株Shan-27,Shan-28和Shan-29接種到PDA平板上培養5 d后,用滅菌打孔器打取直徑5 mm的菌餅貼于催芽后的山黧豆胚根處,對照貼空白PDA,將接種后的山黧豆放置于培養皿內,以無菌水浸濕的濾紙進行保濕,2 d后移去菌餅,正常保濕培養,逐天觀察山黧豆胚根發病情況。
拌菌土法:接種體的制備參考玉米鐮孢莖腐病繁殖體的制備[14]略作修改,將供試菌株Shan-27,Shan-28和Shan-29接種到PDA平板上培養7 d后,用滅菌打孔器打取直徑10 mm的菌餅5塊,接種于滅菌的玉米粒基質中,室溫培養15 d后備用。將山黧豆種子在75%乙醇中浸泡5 min進行表面消毒后,用無菌水沖洗3次,晾干后播種于裝有滅菌蛭石的花盆中(每盆播種20~25粒),每盆接種10 g接種體(分別接種了分離物Shan-27,Shan-28和Shan-29菌株的玉米粒),對照不接菌,每處理3次重復,置于室溫下正常管理,逐天觀察發病癥狀,于播種后25 d統計發病率并拍照。
1.4 病原菌的形態觀察
將分離物接種于PDA平板上于25℃恒溫培養,并觀察菌落形狀、色澤及生長速度等特征。培養5 d后在顯微鏡下觀察分生孢子的形狀、大小、隔膜數等。參考Leslie等的鐮孢菌實驗手冊[15]對病原菌進行形態學鑒定。
1.5 EF-1a序列分析
在PDA培養基上將菌株Shan-27,Shan-28和Shan-29進行培養,直接刮去菌絲后用濾紙吸干水分備用。選用EF-1α引物[16]進行PCR擴增。參考Geiser等[17]的方法進行PCR擴增并測序。獲得的序列與從GenBank數據庫中獲得的相關鐮孢菌EF-1α基因序列進行比較,利用Megalign 7.1軟件以NJ法構建系統發育樹,確定菌株Shan-27,Shan-28和Shan-29的分類地位。
2 結果與分析
2.1 山黧豆根腐病田間發病癥狀
通過田間調查發現,受害山黧豆植株初期葉片發黃,隨著病情進一步擴展植株出現萎蔫、倒伏,嚴重者植株枯死(圖1A),拔出植株觀察其根莖部發現發病植株地下部分須根數減少,根系生長受阻,嚴重的根系變褐腐爛。正常山黧豆植株葉色濃綠,根系發達,生長良好(圖1B)。

圖1 山黧豆根腐病田間發病癥狀Fig.1 The symptom of L.sativus root rot in the field注:A為發病山黧豆;B為正常山黧豆Note:A,disease L.sativus;B, normal L.sativus
2.2 山黧豆根腐病分離純化結果
通過組織塊分離、單孢純化,從感病植株根頸部共分離得到39株分離物,其中3株分離物初步鑒定為禾谷鐮孢菌復合種(F.graminearumspecies complex)(菌株編號為:Shan-27,Shan-28,Shan-29),分離頻率為7.69%。
2.3 致病性測定結果
選取初步確定的3株禾谷鐮孢菌復合種(F.graminearumspecies complex)分離物,采用胚根貼菌餅法接種山黧豆胚根處。結果表明,接種山黧豆后胚根均能發病,發病部位初現褐色,進而隨著病斑的擴展,整個根系及根尖均變褐(圖2A),3個菌株的發病率均達到100%,而對照均無明顯發病癥狀(圖2B)。從病根處再次分離得到的菌株其性狀均與第一次分離獲得的菌株相同。
用初步確定的3株禾谷鐮孢菌復合種分離物的帶菌基質拌土接種山黧豆種子。結果表明,接種后山黧豆植株均能發病。接種后20 d植株出現萎蔫,隨著病情進一步發展,植株倒伏、枯死(圖3A),拔出植株,可見其根頸部發褐,嚴重的根系變褐腐爛、生長受阻(圖3B),對照均未發病(圖3C,D)。對發病根莖部進行病原菌再分離、純化培養,得到與原接種所用菌株相同的病原菌,分離率為100%。說明分離物Shan-27,Shan-28及Shan-29是山黧豆根腐病的致病菌。同時致病性測定表明,分離物Shan-27,Shan-28和Shan-29接種山黧豆后,引起的植株發病率分別為73.75%,66.22%和89.20%,3個菌株均表現對山黧豆具有較強致病性,可作為后續研究菌株。

圖2 接種菌株Shan-29后山黧豆根莖部發病癥狀Fig.2 The symptom of L.sativus after inoculation with Shan-29 strain注:A為接種7 d后的癥狀;B為對照Note:A,the symptom after inoculation seven days;B,the control

圖3 接種菌株Shan-29后山黧豆發病癥狀Fig.3 The symptom of L.sativus after inoculation with Shan-29 strain注:A,B為接種后25 d發病癥狀;C,D為對照Note:A,B,the symptom after inoculation twenty-five days;C,D,the control
2.4 病原菌菌落形態及顯微特征
該菌在PDA培養基上氣生菌絲白色(圖4A),絮狀、繁茂、致密,培養7 d后菌落背面產生玫瑰紅色素(圖4B)。25℃和28℃條件下在PDA上培養3 d的平均生長速度分別為20.2 mm·d-1和20.7 mm·d-1。在CLA上培養20 d后顯微觀察產生大型分生孢子,大型分生孢子體長(2.95~4.88)μm×(14.65~40.48)μm,隔膜多見4~5個,偶見3個或6個,頂胞呈鳥嘴狀,稍彎,足胞不明顯(圖4C,D)。
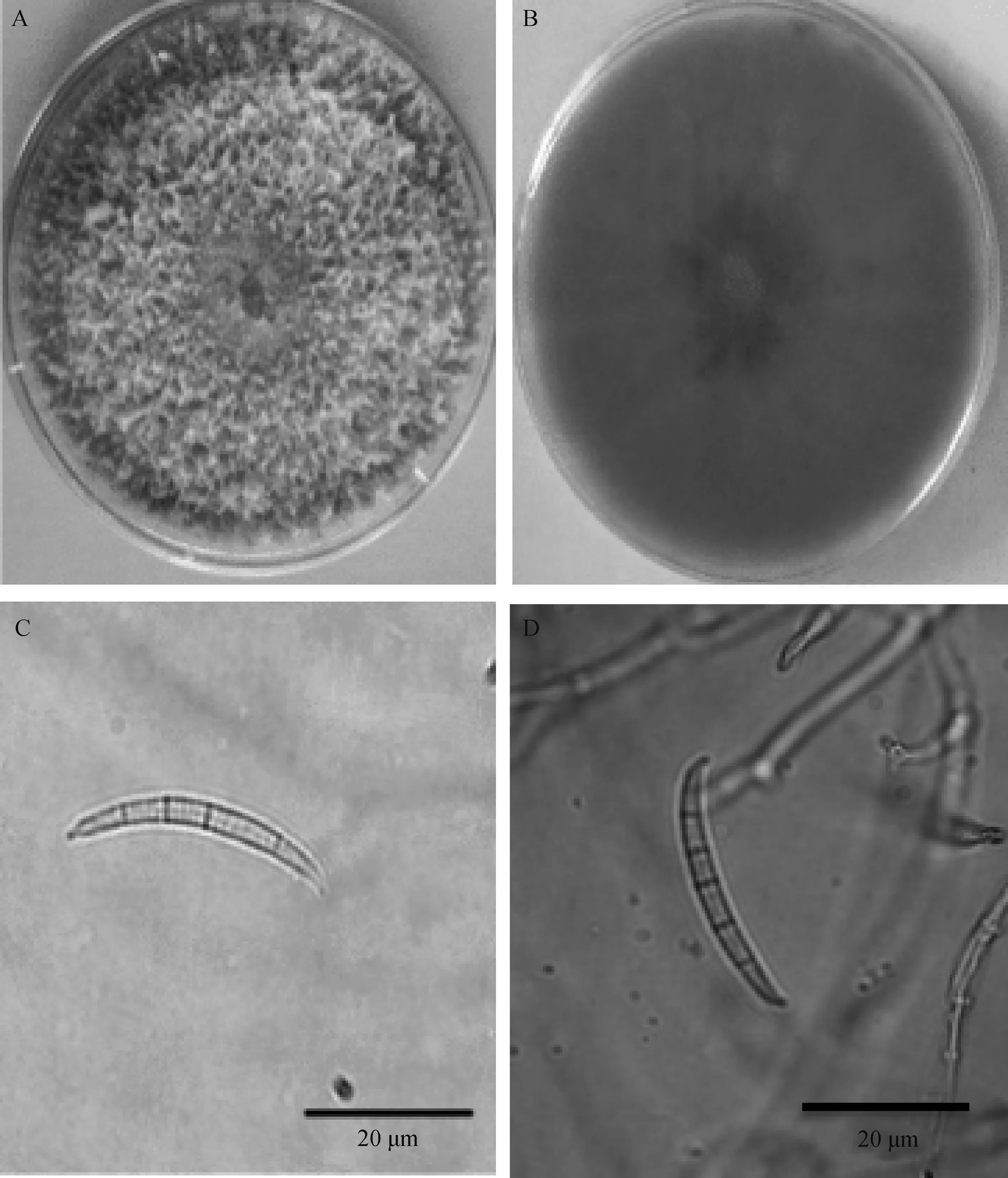
圖4 菌株Shan-29形態特征Fig.4 The morphology of Shan-29 strain注:A為PDA上培養7 d的菌落形態;B為PDA上培養7 d的菌落背面及產生的色素;C,D為分生孢子Note:A,the colony front morphology on PDA after cultivation seven days;B,the colony back morphology on PDA after cultivation seven days;C,D,conidium
2.5 EF-1α基因序列分析
測序后所得序列經與GenBank數據庫中鐮孢菌屬EF-1α基因序列比對發現:菌株Shan-27,Shan-28和Shan-29的EF-1α基因序列與亞洲鐮孢菌(KM062024,LC546964和LC500692)的親緣關系最近,同源性達99%以上。利用Megalign 7.0軟件,采用NJ法構建基于EF-1α基因序列系統發育樹,3個菌株與亞洲鐮孢菌(KM062024,LC546964和LC500692)均聚類于同一分支上。因此,結合形態學觀察與分子生物學分析,將分離獲得的3個致病菌株Shan-27,Shan-28和Shan-29鑒定為亞洲鐮孢菌(F.asiaticum)。

圖5 基于EF-1α建立的菌株Shan-29,Shan-28和Shan-27的系統發育樹Fig.5 Phylogenetic tree of isolated Shan-29,Shan-28 and Shan-27 based on EF-1α
3 討論
根腐病是豆科作物重要的土傳病害之一,給豆科作物的種植帶來了巨大的經濟損失。據報道多種病原真菌可引起豆科作物根腐病。國內王春華等[18]報道木賊鐮孢菌(F.equseti)、尖孢鐮孢菌(F.oxysporum)、終極腐霉菌(Pythiumuultimum)和立枯絲核菌(Rhizoctoniasolani)均能引起新疆花蕓豆(Phaseolusvulgaris)根腐病;鄭培娥[19]在對新疆大豆(Glycinemax)根腐病病原菌種類的研究中發現,尖孢鐮孢菌(F.oxysporum)、禾谷鐮孢菌(F.graminearum)、茄腐鐮孢菌(F.solani)、茄腐鐮孢菌藍色變種(F.solanivar.coeruleum)、立枯絲核菌(R.solani)、燕麥鐮孢菌(F.avenum)和瓜果腐霉菌(P.aphanidenatum)是引起新疆大豆根腐病的病原菌;吳仁峰等[20]對武漢豇豆(Vignaunguiculata)根腐病病原菌研究中發現,茄腐鐮孢菌可引起該地區豇豆根腐病;楊帥等[21]研究認為,層出鐮孢菌(F.proliferatum)可引起江蘇省和安徽省大豆根腐病;張河慶等[22]研究認為,共享鐮孢菌(F.commune)可引起四川省成都市豇豆根腐病;閆文雪等[23]對山東省豇豆根腐病的病原菌類型研究中發現,茄病鐮孢菌大豆專化型(F.solanif.sp.glycines)可引起該地區豇豆根腐病;從麗麗等[24]對內蒙古臨河、山西陽高縣和河北廊坊紫花苜蓿根腐病病原研究中發現,三線鐮孢菌(F.tricinctum)、腐皮鐮孢菌(F.solani)和層出鐮孢菌具有較強致病性,可引致苜蓿根腐病發生。國外Suleiman M N等[25]認為,瓜果腐霉與立枯絲核菌是尼日利亞豇豆根腐病的致病菌;Satish L等[26]認為,菜豆殼球孢菌(Macrophominaphaseolina)可引起印度豇豆根腐病。本研究結果表明,引致四川省南充市山黧豆根腐病的病原為亞洲鐮孢菌(F.asiaticum)。由此可見,引起不同豆科植物根腐病的病原菌不盡相同,不同地區的優勢菌也存在明顯差異。亞洲鐮孢菌是瘤座孢科(Tuberculariaceae)鐮孢菌屬(Fusarium)的重要植物病原真菌,可引起多種植物病害,如侵染小麥(Triticumeastivum)、大麥(Hordeumvulgare)和玉米(Zeamays)等引起赤霉病、莖腐病、穂腐病等[27-29],但作為山黧豆根腐病的病原菌在國內外鮮有報道。
亞洲鐮孢菌作為禾谷鐮孢菌復合種之一,其在形態上很接近其姊妹種禾谷鐮孢菌,因此采用傳統的形態學方法很難準確將其區分開來;為了更準確、可靠地將其鑒定到種,本研究在形態學研究的基礎上,通過EF-1α基因序列分析進一步明確其分類地位,將其鑒定為亞洲鐮孢菌。
本研究明確了亞洲鐮孢菌可引起南充市山黧豆根腐病,但在四川省其他地區或其他種植山黧豆省份是否也分布有亞洲鐮孢菌還有待于進一步研究。
4 結論
經調查發現,2018年8月四川省南充市山黧豆根腐病發生嚴重,從病株上分離獲得3株分離物對山黧豆具有較強致病性,通過形態學觀察和EF-1α基因序列分析將3個致病菌株確定為亞洲鐮孢菌(F.asiaticum)。本研究明確了四川省南充市山黧豆根腐病的病原分類地位,對該病害的進一步研究和防治提供了科學依據。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
