miR-215-5p通過靶向NCOA3基因抑制固始雞腹部前脂肪細(xì)胞的增殖和分化
2021-07-08 01:01:18靳文姣翟彬苑鵬濤范升鑫李遠(yuǎn)方閆峰賓孫桂榮田亞東康相濤李國喜
畜牧與獸醫(yī) 2021年7期
靳文姣,翟彬,苑鵬濤,范升鑫,李遠(yuǎn)方,閆峰賓,孫桂榮,田亞東,康相濤,李國喜
(河南農(nóng)業(yè)大學(xué)動物科技學(xué)院/河南省家禽種質(zhì)資源創(chuàng)新工程研究中心,河南 鄭州 450046)
家禽生產(chǎn)中,腹部脂肪組織的特征與雞的經(jīng)濟(jì)性狀高度相關(guān)[1]。腹腔內(nèi)脂肪過度沉積常常對肉質(zhì)性能產(chǎn)生負(fù)面影響,并增加飼料成本[2-4]。近年來,隨著規(guī)模化養(yǎng)殖的發(fā)展,地方雞種腹脂過度沉積現(xiàn)象也越來越嚴(yán)重[5]。地方雞種是我國家禽育種和優(yōu)質(zhì)雞生產(chǎn)的重要素材,因此,理解地方雞腹部脂肪組織發(fā)育的分子機制,將有益于資源的保護(hù)和開發(fā)利用。固始雞是主要分布于河南省固始縣境內(nèi)的蛋肉兼用型地方品種,因其肉質(zhì)具有鮮嫩、可口和風(fēng)味獨特的優(yōu)良特征,而經(jīng)常被用作育種和生產(chǎn)的素材。但目前仍然缺乏對固始雞腹部脂肪組織發(fā)育分子機制的理解。
MicroRNA (miRNA)是一類長度18~24 nt的內(nèi)源性小的非編碼RNA,可與特定mRNA的3′-UTR區(qū)結(jié)合,在轉(zhuǎn)錄后水平上調(diào)控基因的表達(dá)[6],廣泛參與機體發(fā)育、代謝以及細(xì)胞增殖、分化、凋亡等生物學(xué)過程的調(diào)控[7-8]。脂肪組織發(fā)育涉及新脂肪細(xì)胞的增加(增生)和脂肪細(xì)胞中脂類蓄積的增加(肥大)等復(fù)雜的過程[9],許多研究顯示miRNA也參與脂肪細(xì)胞增殖、分化和脂類代謝等脂肪組織發(fā)育相關(guān)的許多生物學(xué)過程調(diào)控。例如,bat-miR-33b[10]、miR-21[11]、miR-204[12]、 miR-378[13]、miR-125a-5p[14]、miR-200b[15]等miRNA已被證實可參與脂肪細(xì)胞增殖或分化調(diào)控。這說明,miRNA可作為研究動物脂肪組織發(fā)育相關(guān)分子機制的新靶標(biāo)。但目前的一些研究還僅局限于少數(shù)農(nóng)業(yè)動物。
有關(guān)雞腹脂發(fā)育相關(guān)miRNA的研究已有報道[16-20]。例如,Huang等[18]通過深度測序,從93日齡慢長型中國地方雞北京油雞(低腹脂)和快長型商業(yè)化肉雞品系科寶雞(高腹脂)的腹脂中鑒定到了230種已知miRNA和83種潛在的新miRNA。Yao等[19]和Wang等[20]分別從雞腹脂衍生前脂肪細(xì)胞中檢測到159種已知miRNA和33種差異miRNA。這些研究表明,雞腹脂發(fā)育受許多miRNA的調(diào)控。但先前的研究還主要集中于少數(shù)品種和腹脂發(fā)育特定階段miRNA鑒定,而對特定miRNA在雞腹脂發(fā)育中的作用機制研究還很少。
前期,我們完成了6、14、22、30周齡固始雞腹脂miRNA的高通量測序,篩選鑒定到507種已知miRNA和53種新的miRNA,發(fā)現(xiàn)miR-215-5p在固始雞腹脂4個發(fā)育階段間高豐度差異性表達(dá)[21]。NCOA3基因可與核激素受體互作,與脂類代謝和沉積密切相關(guān)[22]。Wang等[23]研究顯示,NCOA3基因的表達(dá)量隨著肌內(nèi)脂肪含量的增加而升高,表明NCOA3基因?qū)χ境练e可能存在正向調(diào)控作用。靶基因預(yù)測表明,在NCOA3基因3′-UTR上,存在miR-215-5p的結(jié)合位點。這表明,miR-215-5p可能通過靶向作用于NCOA3而調(diào)控固始雞腹脂發(fā)育。基于此,本文研究了miR-215-5p對固始雞腹部前體脂肪細(xì)胞增殖和分化的影響及其與NCOA3基因的靶向關(guān)系,所得結(jié)果為更好地理解固始雞腹部脂肪組織發(fā)育的分子機制提供新的見解。
1 材料與方法
1.1 試驗動物與樣品采集
選擇14周齡健康固始雞6只,解剖后,取心臟、肝臟、脾臟、胰腺、腿肌、皮脂、腎臟、腹脂等組織,液氮速凍后保存于-80 ℃冰箱待用,用于miR-215-5p組織表達(dá)譜分析。另外,選擇14日齡固始雞雛雞,解剖分離腹部脂肪組織,用于原代前體脂肪細(xì)胞分離培養(yǎng)。
1.2 原代前脂肪細(xì)胞分離培養(yǎng)
取14日齡固始雞雛雞,無菌條件下處死后于75%酒精中浸泡10 min,解剖分離腹部脂肪組織,用含0.8%鏈霉素/青霉素的PBS連續(xù)沖洗3次后剪碎。將剪碎后的組織塊置于含1 mg/mL膠原蛋白酶Ⅱ(9001-12-1,索萊寶)的DMEM(SH30023.01,HyClone)中,37 ℃消化1 h。消化完全后加入紅細(xì)胞裂解液,室溫下裂解10 min,隨后分別通過100目、200目和500目的篩網(wǎng)過濾,1 000 r/min 的轉(zhuǎn)速離心濾液5 min。用完全培養(yǎng)基(DMEM+10% 胎牛血清(04-001-1ACS,BI)+1%雙抗(P1400,索萊寶)重懸細(xì)胞后,鋪于培養(yǎng)瓶中,在37 ℃、5% CO2培養(yǎng)箱中培養(yǎng)。前脂肪細(xì)胞的誘導(dǎo)分化,于6孔板中進(jìn)行,當(dāng)細(xì)胞達(dá)到90%融合時,用含160 mol/L油酸的完全培養(yǎng)基進(jìn)行誘導(dǎo)分化,每隔1 d更換1次培養(yǎng)基,分別在誘導(dǎo)后0、2、4、6、8 d收集細(xì)胞樣品。
1.3 細(xì)胞轉(zhuǎn)染試驗
采用lipofactamine 2000試劑(11668019,invitrogen),按照試劑盒操作說明,將濃度為50 nmol/L 的miR-215-5p mimics(5′-AUGACCUAUGAAUUGACAGCA-3′,5′-CUGUCAAUUCAUAGGUCAUUU-3′)和mimics NC(5′-UUCUCCGAACGUGUCACGUTT-3′,5′-ACGUGACACGUUCGGAGAATT-3′)以及濃度為100 nmol/L的miR-215-5p 抑制劑(5′-GUCUGUCAAUUCAUAGGUCAU-3′)和抑制劑對照 (5′-CAGUACUUUUGUGUAGUACAA-3′)(銳博生物)分別轉(zhuǎn)染到前體脂肪細(xì)胞中。試驗在12孔板或96孔板中進(jìn)行,當(dāng)細(xì)胞融合達(dá)到60% ~ 70%(增殖試驗)或70% ~ 80%(分化試驗)時,完成轉(zhuǎn)染;轉(zhuǎn)染后6 h更換為完全培養(yǎng)基,48 h后收集細(xì)胞樣品。
1.4 實時熒光定量RCR(qRT-PCR)分析
分別采用Primer SeriptTMRT reagent Kit和Mir-XTMmiRNA First-Strand Synthesis試劑盒(R323,R312,諾維贊)對總RNA進(jìn)行反轉(zhuǎn)錄,獲得mRNA、miRNA所對應(yīng)的cDNA。定量PCR分析,在Roche LightCycler?96儀器上進(jìn)行;mRNA所用引物序列見表1,由尚亞生物技術(shù)有限公司合成;miR-215-5p和U6的反轉(zhuǎn)錄和定量引物購自銳博生物有限公司,RT-PCR反應(yīng)程序為 :95 ℃預(yù)變性 5 min;95 ℃變性 12 s,60 ℃退火 40 s,72 ℃延伸 30 s,共40個循環(huán)。所有試驗均重復(fù)3次。分別以U6、β-actin作為miRNA和mRNA定量的內(nèi)參基因,采用2-ΔΔCt法計算miRNA或mRNA相對表達(dá)量。

表1 qRT-PCR分析所用引物
1.5 CCK-8和EdU分析
通過CCK-8 (MA0218,美侖生物) 和EdU(C10310-1,銳博生物)分析,評價miR-215-5p對前脂肪細(xì)胞增殖的影響。當(dāng)接種細(xì)胞生長至60% ~ 70%融合時,將miR-215-5p mimics和mimics NC以及miR-215-5p inhibitors和inhibitors NC轉(zhuǎn)染細(xì)胞。對于CCK-8分析,按照試劑盒操作說明在96孔板上進(jìn)行,分別于轉(zhuǎn)染后12、24、36、48 h,向每孔細(xì)胞各加入10 μL CCK-8試劑,置于37 ℃孵育2 h,用酶標(biāo)儀(波長450 nm處)檢測細(xì)胞活力。對于EdU分析,在6孔板上進(jìn)行,轉(zhuǎn)染后 36 h,用25 μmol/L濃度的EdU溶液孵育細(xì)胞4 h,染色后使用熒光倒置顯微鏡拍照。上述試驗各處理均設(shè)置6個重復(fù)。
1.6 油紅O染色和甘油三酯含量分析
將前體脂肪細(xì)胞接種于12孔板內(nèi),待細(xì)胞70%~80%融合,轉(zhuǎn)染miR-215-5p mimics和mimics NC以及miR-215-5p inhibitors和inhibitors NC,并于轉(zhuǎn)染24 h后更換含160 mol/L油酸(112-80-1,索萊寶)的誘導(dǎo)培養(yǎng)基,誘導(dǎo)分化后24 h,進(jìn)行油紅O染色和甘油三酯含量分析,以評價脂肪細(xì)胞充脂情況。對于油紅O染色,用4%多聚甲醛溶液固定處理細(xì)胞30 min后,加油紅O染液,室溫孵育20 min,棄掉染液用PBS清洗細(xì)胞3次,熒光倒置顯微鏡下拍照。然后,向每孔染色后的細(xì)胞中加入500 μL異丙醇,10 min后用酶標(biāo)儀在500 nm波長處測量OD值。另外,采用組織細(xì)胞甘油三酯(TG)酶法試劑盒 (E1013-105,普利萊) 檢測脂肪細(xì)胞內(nèi)甘油三酯含量,細(xì)胞裂解后,按照試劑盒操作對裂解后的細(xì)胞進(jìn)行處理,用酶標(biāo)儀檢測蛋白含量(595 nm處)和甘油三酯含量(550 nm處)。上述試驗各處理均設(shè)置3個重復(fù)。
1.7 載體構(gòu)建和雙熒光素酶活性分析
使用正向引物5′-CCGCTCGAGGTGCTGTACAAAAAAGGTCATATTTTTGGC-3′ 和反向引物5′-ATAAGAATGCGGCCGCGGTATTTCAGGGCTGGCTCTAC-3′擴增包含miR-215-5p結(jié)合位點的NCOA3-3′-UTR序列(NCOA3-3′-UTR-WT);使用突變引物(F:5′-CCGCTCGAGGTGCTGTACAAAAAGTTCAGTATTTTTGGC-3′, R:5′-ATAAGAATGCGGCCGCGGTATTT-CAGGGCTGGCTCTAC-3′)擴增包含突變miR-215-5p結(jié)合位點的NCOA3-3′-UTR序列(NCOA3-3′-UTR-Mut), 片段長度均為130 bp。2個片段擴增成功后,進(jìn)行膠回收,用XhoⅠ、NotⅠ限制性內(nèi)切酶進(jìn)行雙酶切并分別與T載體連接;隨后再利用XhoⅠ、NotⅠ限制性內(nèi)切酶,對NCOA3-3′-UTR-WT、NCOA3-3′-UTR-Mut和 psiCHECK2空載體進(jìn)行雙酶切,并將酶切產(chǎn)物連接psiCHECK2載體。最后,對成功連接psiCHECK2載體的菌液進(jìn)行大量搖菌,提取重組質(zhì)粒,分別命名為WT-NCOA3和MUT-NCOA3。
熒光素酶活性分析,在雞成纖維細(xì)胞系(DF1細(xì)胞)上完成,細(xì)胞于37 ℃、5% CO2培養(yǎng)箱中進(jìn)行培養(yǎng)。在24孔板內(nèi),當(dāng)細(xì)胞生長至70%融合時,利用lipofactamine 2000試劑,將miR-215-5p mimics與WT-NCOA3或MUT-NCOA3分別共轉(zhuǎn)染DF1細(xì)胞。轉(zhuǎn)染后48 h,按照Dual-Luciferase報告基因檢測系統(tǒng)試劑盒 (Promega) 操作說明,對細(xì)胞進(jìn)行裂解并檢測雙熒光素酶活性。
1.8 統(tǒng)計分析
所有數(shù)據(jù)均采用“平均值±標(biāo)準(zhǔn)誤”表示,使用SPSS 22.0軟件進(jìn)行統(tǒng)計分析。P值的計算采用非配對t檢驗,P<0.05表示差異顯著,P<0.01表示差異極顯著。
2 結(jié)果
2.1 雞miR-215-5p的表達(dá)譜特征
采用qRT-PCR分析了miR-215-5p在固始雞14周齡8種組織中以及腹部前體脂肪細(xì)胞分化過程中的表達(dá)情況。結(jié)果顯示,miR-215-5p在腹脂中相對表達(dá)量最高,其次為腎臟和皮脂(圖1A),這表明miR-215-5p可能與雞腹部脂肪沉積相關(guān)。另外,在前脂肪細(xì)胞分化的前兩天,miR-215-5p表達(dá)水平呈遞增趨勢,至第2天達(dá)第一個峰值;從前脂肪細(xì)胞分化后第4天,miR-215-5p的表達(dá)又逐漸上升,到分化后第8天達(dá)到峰值(圖1B)。前體脂肪細(xì)胞分化前期伴隨著細(xì)胞增殖,因此miR-215-5p的這種動態(tài)表達(dá)結(jié)果表明,其在雞前體脂肪細(xì)胞增殖和分化過程中可能均發(fā)揮作用。
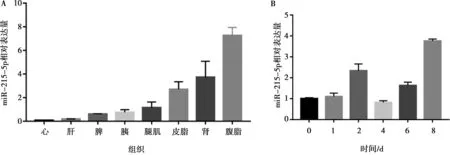
A. 在不同組織中的表達(dá);B. 在腹部前脂肪細(xì)胞分化不同時間點的表達(dá)
2.2 miR-215-5p抑制雞腹部前體脂肪細(xì)胞增殖
分別將miR-215-5p mimics和mimics NC以及miR-215-5p inhibitors和inhibitors NC轉(zhuǎn)染固始雞腹部前體脂肪細(xì)胞,以評價miR-215-5p對雞前脂肪細(xì)胞增殖的影響。當(dāng)轉(zhuǎn)染miR-215-5p mimics后,前脂肪細(xì)胞中miR-215-5p表達(dá)水平提高了350倍(圖2A);相反,轉(zhuǎn)染miR-215-5p inhibitors后,前脂肪細(xì)胞中miR-215-5p的表達(dá)水平下降了50倍(圖2B)。實時熒光定量PCR分析顯示,轉(zhuǎn)染miR-215-5p mimics的前脂肪細(xì)胞中,細(xì)胞增殖標(biāo)志基因(CDK1和PCNA)的表達(dá)水平比對照組極顯著下降(P<0.01),而細(xì)胞周期阻滯因子(p21)的表達(dá)水平比對照組極顯著升高(P<0.01)(圖2C)。相反,在轉(zhuǎn)染miR-215-5p inhibitors的前脂肪細(xì)胞中,與對照組相比CDK1和PCNA的表達(dá)水平顯著上升(P<0.05),p21的表達(dá)水平顯著下降(P<0.05)(圖2D)。EdU分析表明,過表達(dá)miR-215-5p可顯著降低EdU陽性細(xì)胞比例(圖3A,3B),而抑制miR-215-5p可顯著提高EdU陽性細(xì)胞的比例(圖4A,4B)。CCK-8檢測證實,轉(zhuǎn)染miR-215-5p mimics可顯著降低前脂肪細(xì)胞總數(shù)(圖3C),而轉(zhuǎn)染miR-215-5p inhibitors可顯著增加前脂肪細(xì)胞總數(shù)(圖4C)。這些結(jié)果證明,miR-215-5p可抑制雞腹部前脂肪細(xì)胞的增殖。
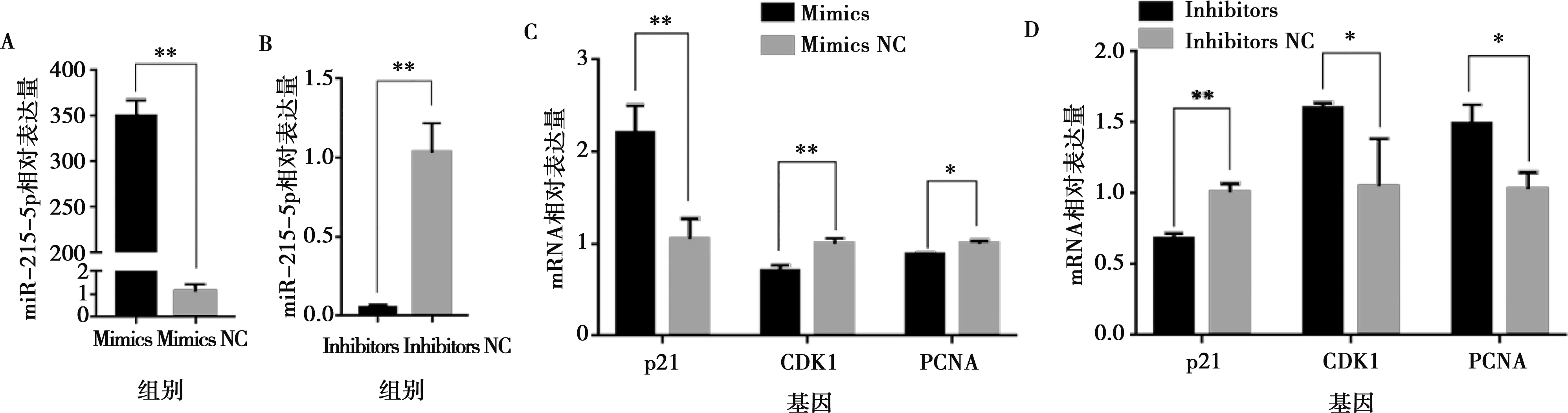
A和B分別為miR-215-5p mimics和inhibitors轉(zhuǎn)染后前脂肪細(xì)胞中miR-215-5p的表達(dá)量;C和D分別是miR-215-5p mimics和inhibitors轉(zhuǎn)染后前脂肪細(xì)胞中增殖標(biāo)志基因的表達(dá)量。*P<0.05,**P<0.01。下同

A.EdU檢測分析;B. EdU陽性細(xì)胞占比統(tǒng)計;C. CCK-8分析
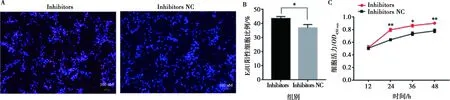
A. EdU檢測分析;B. EdU陽性細(xì)胞占比統(tǒng)計;C. CCK-8分析
2.3 miR-215-5p抑制雞腹部前體脂肪細(xì)胞分化
將miR-215-5p mimics和mimics NC以及miR-215-5p inhibitors和inhibitors NC分別轉(zhuǎn)染固始雞腹部前脂肪細(xì)胞并進(jìn)行誘導(dǎo)分化,以評價miR-215-5p對雞前脂肪細(xì)胞分化的影響。結(jié)果顯示,與對照組相比,轉(zhuǎn)染miR-215-5p mimics可顯著抑制脂肪細(xì)胞分化標(biāo)志基因PPARG、CEBPA、FABP4和LPL的表達(dá)(圖5A),而miR-215-5p inhibitors處理則得到了相反的結(jié)果(圖5B)。相應(yīng)地,過表達(dá)miR-215-5p顯著減少了分化脂肪細(xì)胞中脂滴的數(shù)量,降低了脂質(zhì)積累(圖6);抑制miR-215-5p表達(dá)則顯著增加了分化脂肪細(xì)胞中脂滴的數(shù)量,促進(jìn)了脂質(zhì)積累(圖7)。同樣,轉(zhuǎn)染miR-215-5p mimics的脂肪細(xì)胞中甘油三酯含量極顯著降低(P<0.01),而轉(zhuǎn)染miR-215-5p inhibitors的脂肪細(xì)胞中甘油三酯含量則顯著升高 (P<0.05)(圖8)。這些結(jié)果表明,miR-215-5p可抑制雞腹部前體脂肪細(xì)胞的分化。
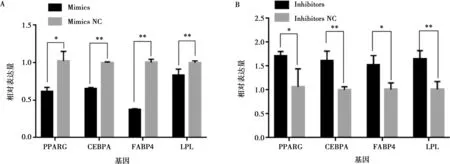
圖5 過表達(dá)(A)/抑制(B)miR-215-5p對脂肪細(xì)胞分化標(biāo)志基因相對表達(dá)量的影響
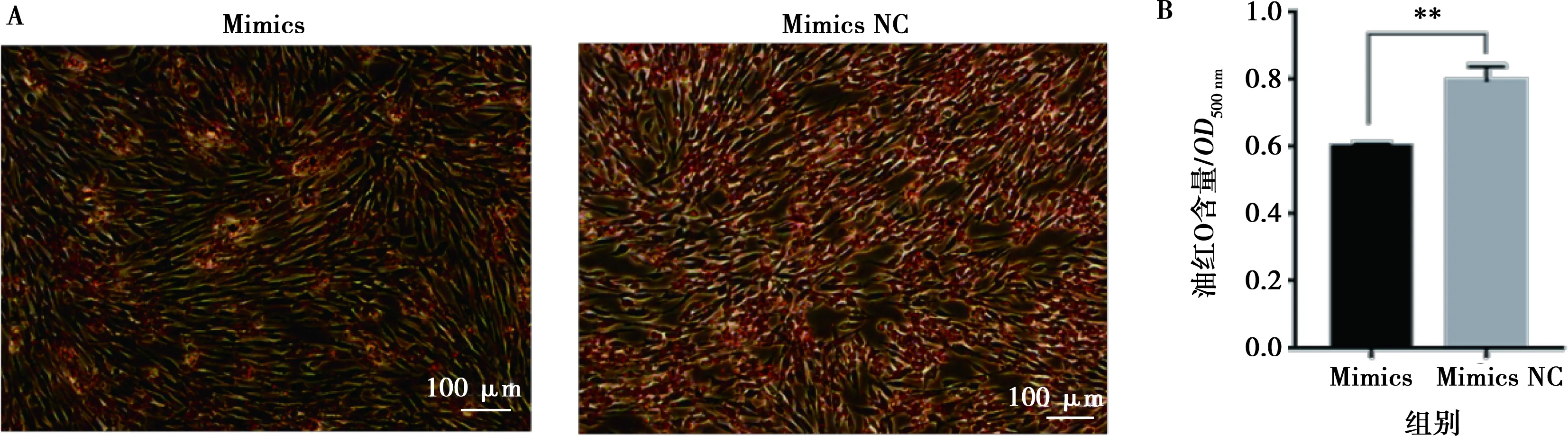
A.油紅O染色;B.油紅O含量分析(500 nm波長處讀取的OD值)
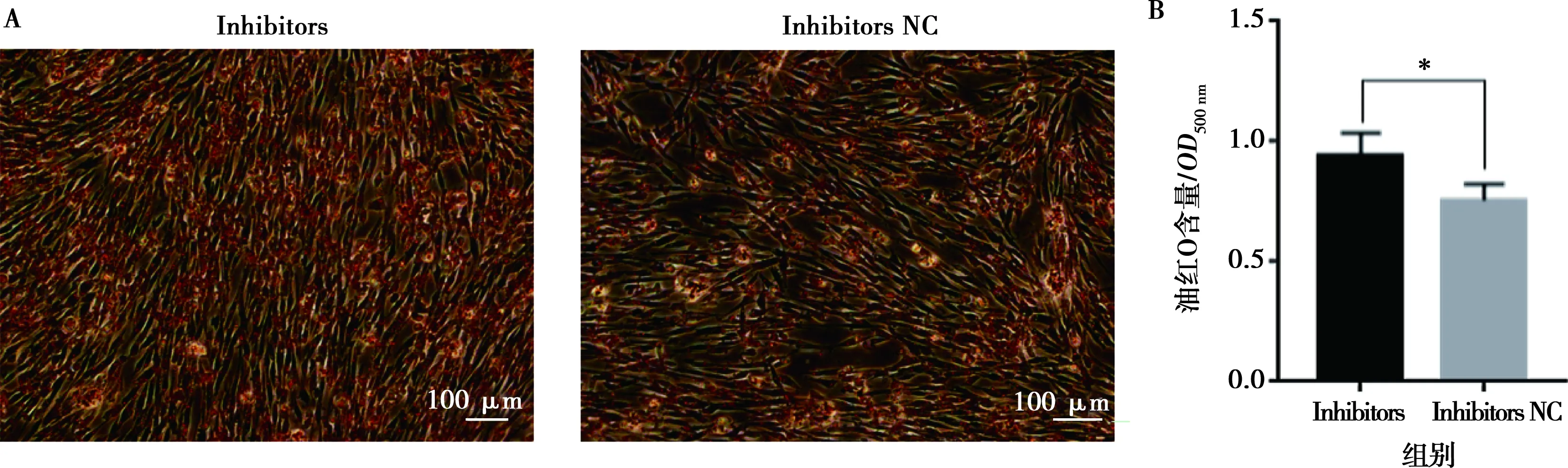
A.油紅O染色;B.油紅O含量分析(500 nm波長處讀取的OD值)
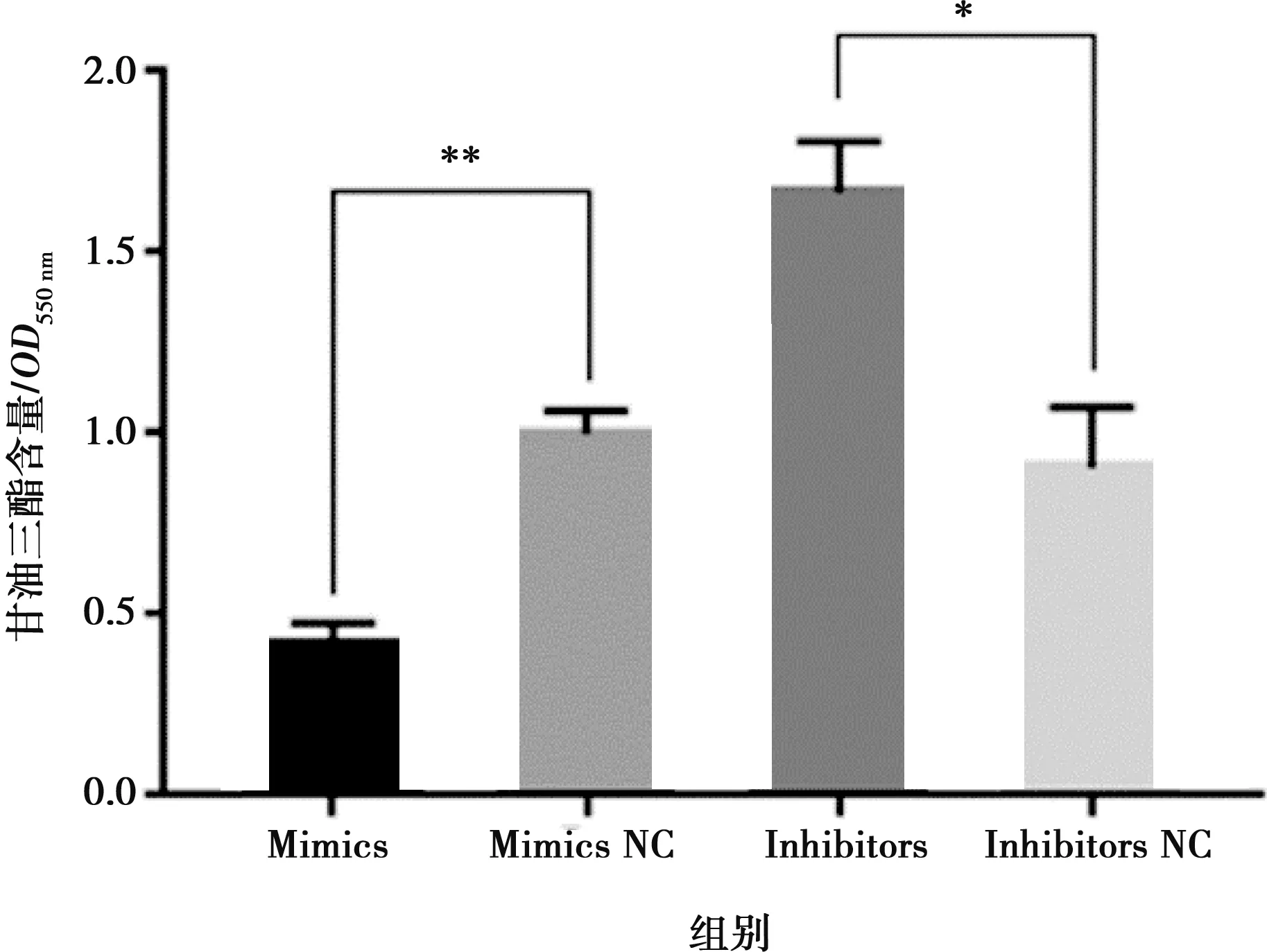
圖8 過表達(dá)/抑制miR-215-5p后脂肪細(xì)胞內(nèi)甘油三酯
2.4 miR-215-5p可直接靶作用于NCOA3基因
為揭示miR-215-5p在雞腹部前脂肪細(xì)胞增殖和分化中的作用機制,利用TargetScan在線軟件預(yù)測發(fā)現(xiàn),雞NCOA3基因3′-UTR存在miR-215-5p種子區(qū)序列的結(jié)合位點(圖9A)。在細(xì)胞中過表達(dá)miR-215-5p時,可顯著抑制NCOA3基因的表達(dá)(P<0.01);相反,抑制miR-215-5p表達(dá)時可顯著促進(jìn) NCOA3基因的表達(dá)(P<0.05)(圖9B)。這表明,在雞腹部前體脂肪細(xì)胞增殖和分化過程中,miR-215-5p和NCOA3基因可能存在潛在的靶向互作關(guān)系。基于此,在DF1細(xì)胞上,通過雙熒光素酶報告系統(tǒng)試驗,驗證了miR-215-5p和NCOA3的直接靶向關(guān)系。結(jié)果顯示,將miR-215-5p mimics和野生型或突變型psiCHECK2-NCOA3-3′UTR載體共轉(zhuǎn)染DF1細(xì)胞48 h后,miR-215-5p可顯著抑制野生型載體熒光素酶活性(P<0.01),而miR-215-5p與突變型載體共轉(zhuǎn)染則可解除該抑制效果(圖9C)。結(jié)果表明,miR-215-5p可直接靶作用于NCOA3基因。
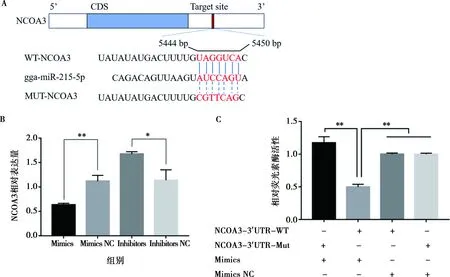
A.miR-215-5p在NCOA3基因mRNA 3′-UTR的結(jié)合位點和突變位點序列;B.過表達(dá)/干擾miR-215-5p后脂肪細(xì)胞中NCOA3的相對表達(dá)量;C.雙熒光素酶報告試驗
3 討論
miR-215屬于mir-192基因家族成員,其在物種間高度保守,對維持物種進(jìn)化具有重要作用[24]。在人類中,miR-215為基因內(nèi)miRNA,與其宿主基因一起轉(zhuǎn)錄;而miR-215宿主基因異亮氨酰tRNA合成酶2(isoleucyl-tRNA synthetase 2,IARS2)為丙氨酰tRNA合成酶的一種,在蛋白質(zhì)合成第一步催化氨基酸與同源tRNA的共價結(jié)合。研究顯示,miR-215和IARS2的表達(dá)在Hirschsprung疾病患者中呈正相關(guān)[25]。另外,miR-215位于常見的脆弱位點(common fragile site)FRA1H(1q41-q42.1)內(nèi),而該位點在許多類型的癌癥中缺失[26]。因此,miR-215是人類疾病中被廣泛研究的一個miRNA,尤其是在不同類型的癌癥中作為致癌基因而受到重視[24]。與此相比,有關(guān)雞miR-215的研究報道還很少。Cai等[27]證實,miR-215-5p在雞心臟中可靶作用于CTCF,而硒缺乏可通過直接靶向作用于miR-215-5p/CTCF軸影響心肌發(fā)育和分化;同時,也證明miR-215-5p可通過直接結(jié)合PI3K基因3′UTR,通過調(diào)節(jié)PI3K/AKT/TOR通路和 ROS-dependent MAPK通路而調(diào)控心肌細(xì)胞存活,在調(diào)控反饋環(huán)中發(fā)揮自噬調(diào)節(jié)作用,促進(jìn)雞心肌自噬[28]。這表明,miR-215在雞組織器官發(fā)育調(diào)控中也發(fā)揮重要作用。在本研究中,我們通過qRT-PCR分析了雞miR-215-5p的表達(dá)譜特征,發(fā)現(xiàn)miR-215-5p在雞腎臟組織高表達(dá),這與先前報道的 miR-215在人腎臟正常組織中高度富集的結(jié)果相一致[29]。尤其重要的是,在所檢測的8種組織中,miR-215-5p在雞腹部脂肪組織中表達(dá)水平最高,進(jìn)一步的分析顯示miR-215-5p在雞腹部前脂肪細(xì)胞分化過程呈現(xiàn)明顯的時序表達(dá)特征。這表明,miR-215-5p與雞腹脂發(fā)育或脂肪沉積密切相關(guān)。
脂肪組織發(fā)育是新脂肪細(xì)胞增生和脂肪細(xì)胞肥大的結(jié)果[9],而腹部脂肪塊的大小則由該部位脂肪細(xì)胞的數(shù)量和體積決定的。因此,揭示脂肪細(xì)胞增殖和分化機理是闡釋雞腹部脂肪組織發(fā)育及相關(guān)性狀形成分子機制的關(guān)鍵。雖然許多miRNA已被證明可參與動物脂肪細(xì)胞增殖和分化調(diào)控[10-15],但有關(guān)雞腹脂發(fā)育過程特定miRNA功能研究還很少。先前的許多研究已證實,miR-215在細(xì)胞和組織發(fā)育、細(xì)胞凋亡、細(xì)胞周期和增殖、細(xì)胞遷移和侵襲、細(xì)胞微環(huán)境和代謝等基本細(xì)胞過程中發(fā)揮重要調(diào)控作用[24]。為此,本研究通過miR-215-5p mimics和mimics NC、miR-215-5p inhibitors和 inhibitors NC轉(zhuǎn)染試驗,闡明了miR-215-5p對腹部前脂肪細(xì)胞增殖和分化的影響,以解釋miR-215-5p在雞腹部脂肪組織發(fā)育的作用。在前脂肪細(xì)胞增殖方面,我們發(fā)現(xiàn),miR-215-5p mimics處理可提高p21的表達(dá)水平,而顯著降低CDK1和PCNA的表達(dá);相反,miR-215-5p innibitors處理顯著降低了p21的表達(dá)水平,而顯著提高了CDK1和PCNA的表達(dá)水平。已知CDK是細(xì)胞周期的關(guān)鍵調(diào)節(jié)因子[30],PCNA是DNA聚合酶的一種輔助性因子[31],二者對細(xì)胞增殖起著正向調(diào)控的作用;而p21可抑制 CDK家族各成員的活動[32],從而阻滯細(xì)胞增殖。因此,上述結(jié)果表明,miR-215-5p可抑制雞前體脂肪細(xì)胞增殖,尤其是EdU和CCK-8的試驗結(jié)果也證實了這一結(jié)論。已知PPARG、CEBPA和LPL是前脂肪細(xì)胞分化早期標(biāo)志基因[33-34],而FABP4(aP2)則是脂肪細(xì)胞分化后期標(biāo)志基因[35]。當(dāng)固始雞腹部前脂肪細(xì)胞過表達(dá)miR-215-5p時,可顯著抑制這些脂肪分化標(biāo)志基因的表達(dá),同時脂滴蓄積明顯減少;而抑制miR-215-5p則獲得了相反結(jié)果。這些結(jié)果表明,miR-215-5p可抑制雞腹部前體脂肪細(xì)胞分化,這與先前在3T3-L1細(xì)胞上的研究結(jié)果相一致[36]。總之,miR-215-5p是雞腹部前脂肪細(xì)胞增殖和分化的負(fù)調(diào)控因子,其通過抑制前脂肪細(xì)胞的增殖和分化而影響雞腹部脂肪組織的發(fā)育。
miRNA通過與其靶基因mRNA的3′-UTR結(jié)合而在轉(zhuǎn)錄后水平調(diào)控基因的表達(dá)。因此,靶基因鑒定是揭示miRNA作用機制的關(guān)鍵。許多研究證明,一些miRNA可直接靶作用于脂肪形成相關(guān)基因而影響脂類代謝或沉積,如miR-155可直接靶作用于脂肪形成的關(guān)鍵轉(zhuǎn)錄因子C/EBPβ[37],miR-200b-3p在3T3-L1細(xì)胞中過表達(dá)可降低PPARγ的表達(dá)水平[38]。已知NCOA3基因可參與脂類和脂蛋白代謝(metabolism of lipids and lipoproteins)、脂類代謝的PPARα調(diào)控(regulation of lipid metabolism by pero-xisome proliferator activated receptor alpha)、白色脂肪細(xì)胞的轉(zhuǎn)錄調(diào)控(transcriptional regulation of white adipocyte differentiation)等與脂類代謝相關(guān)的通路調(diào)控。先前的一些研究也證實,NCOA3在調(diào)節(jié)小鼠脂肪蓄積中發(fā)揮關(guān)鍵作用[39-40];并且,敲除NCOA3基因后,小鼠模型的脂肪沉積顯著減少[41-42];此外,NCOA3能與PPARγ相結(jié)合,減少PPARγ-S114磷酸化,促進(jìn)PPARγ的轉(zhuǎn)錄活性,從而促進(jìn)脂肪沉積[43]。在豬前脂肪細(xì)胞分化過程中,NCOA3可被miR-17-5p靶向作用[22]。這些研究表明,NCOA3是脂類代謝或沉積的一個重要的正向調(diào)控因子。本研究通過過表達(dá)試驗和熒光素酶報告基因系統(tǒng)也證明,在雞腹部前脂肪細(xì)胞分化過程中,NCOA3是miR-215-5p的一個直接靶基因。因此,miR-215-5p可通過靶作用于NCOA3而負(fù)調(diào)控雞腹部脂肪形成。
生物體內(nèi),一個miRNA能夠調(diào)控許多靶基因。在不同生物學(xué)背景中,miR-215的許多靶基因已經(jīng)被確定。例如,在3T3-L1細(xì)胞中,miR-215-5p通過靶作用于III型纖維連接蛋白結(jié)構(gòu)域3B(fibronectin type Ⅲ domain containing 3B,F(xiàn)NDC3B)和連環(huán)蛋白beta互作蛋白1(catenin, beta interactingprotein 1,CTNNBIP1)而負(fù)調(diào)控3T3-L1細(xì)胞早期脂肪形成[36]。在胃癌中,miR-215可直接靶作用于轉(zhuǎn)錄因子RUNX1(runt-related transcription factor 1)[44],而RUNX1在所有造血部位表達(dá),有助于造血干細(xì)胞和祖細(xì)胞的形成[45]。然而,miR-215-5p在雞這些基因上都沒有結(jié)合位點。事實上,miRNA與基因mRNA的靶向互作關(guān)系因組織、細(xì)胞類型和生理狀態(tài)而不同[46]。因此,我們推測,在雞腹部前脂肪細(xì)胞增殖、分化過程中,miR-215-5p可能還存在許多未知的靶基因,它們形成一個復(fù)雜的調(diào)控網(wǎng)絡(luò),共同參與雞腹脂沉積調(diào)控,下一步將聚焦于雞腹脂形成相關(guān)靶基因的鑒定和互作調(diào)控機制研究。
