脂肽微生物的研究進展
2021-07-09 03:07:08陶雪菊茍興華何成霞
成都大學學報(自然科學版) 2021年2期
陶雪菊,茍興華,何成霞
(成都大學 食品與生物工程學院,四川 成都 610106)
0 引 言
微生物脂肽有天然微生物脂肽和合成微生物脂肽兩大類.其中,天然微生物脂肽是微生物經過活化培養產生的具有抗菌活性的次級代謝產物;合成微生物脂肽是通過在天然微生物脂肽的結構基礎上進行結構修飾而得到[1-2].產生脂肽的微生物有多種,如細菌、放線菌、藍細菌及真菌等,目前為止,芽孢桿菌是發現產脂肽最多的屬類,最早發現的抗菌脂肽也是由芽孢桿菌產生的[3].由于這些微生物具有不同的結構和特性,產生的脂肽也具有不同的抑菌效果[4],如表面活性素類是一種表面活性劑,也具有一定的抗菌能力,伊枯草菌素類具有抗細菌能力,豐原素類具有抗真菌能力.脂肽與其他一些抑菌物質,如與細菌素相比,具有更強的耐熱性和耐蛋白水解酶降解能力.脂肽在生物防治、食品加工、石油開采及醫藥行業有廣泛的應用,目前,對脂肽類物質的化學結構及功能研究比較深入,但對脂肽的抗腫瘤、殺蟲及溶血栓等生物學活性,有待于進一步研究,這些就激發了許多的科研工作者從不同原料上篩選產脂肽的微生物,以挖掘出新的脂肽菌源,研究不同抗菌物質的生理活性,進而推進脂肽在更多領域的應用[5].
1 抗菌脂肽的結構特征與作用機制
1.1 抗菌脂肽的結構特征
抗菌脂肽一般由7~10個氨基酸殘基組成,包含親水性肽鏈和親油性脂肪酸鏈,其中親油性脂肪酸鏈由β-羥基脂肪酸鏈及β-氨基脂肪酸鏈兩部分組成.微生物脂肽在結構上分為環狀脂肽和線形脂肽兩大類.環狀脂肽又分為脂肪酸成環、接環和離環3種,第一種是脂肪酸鏈上存在的一個-OH或一個-NH2與肽鏈上氨基酸的-COOH結合構成酰胺鍵或者內酯鍵,導致肽鏈閉合后形成環狀脂肽;第二種是肽環的-NH2直接與肽環相連,肽鏈的C-端的羧基與N-端氨基酸的-OH或非α-氨基相連形成環狀結構稱為脂肪酸接環的環狀脂肽;第三種是脂肪酸鏈間接和含有肽鍵和多種其他酯鍵、酰胺鍵等的肽環相連而形成的脂肪酸離環的環狀脂肽.線形脂肽又分為螺旋形態和TAN1511(包括TAN1511A、TAN1511B和TAN1511C)兩大類,前者為直鏈結構,后者具有支鏈結構[6].
研究發現,脂肽類抗菌劑主要包含表面活性素、豐原素、伊枯草菌素、桿菌霉素、抗霉菌枯草桿菌素及制磷脂菌素等[7-8].芽孢桿菌屬能產生表面活性素、豐原素和伊枯草菌素3大類脂肽化合物[9].
表面活性素是一類穩定性很高且擁有較高表面活性的表面活性劑,還具有抗真菌、抗細菌、抗病毒和抗腫瘤活性[10].表面活性素肽鏈典型的氨基酸順序為Glu(L)-Leu(L)-Leu(D)-Val(L)-Asp(L)-Leu(D)-Leu(L),其分子結構如圖1所示[11-12].
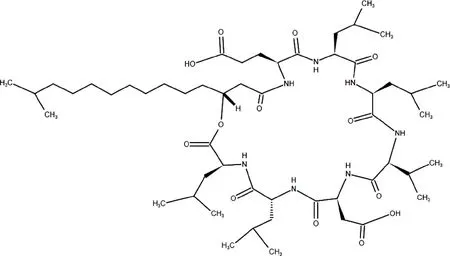
圖1 表面活性素分子結構圖
豐原素是一類抗真菌抑制劑,特別是絲狀真菌產生的豐原素有兩種類型,即豐原素A和豐原素B.其中,豐原素A類的肽鏈第6位上是Ala,豐原素B類的肽鏈第6位上是Val.豐原素A的分子結構如圖2所示[13].

圖2 豐原素A分子結構圖
伊枯草菌素是一類脂肽化合物,有多種類型,包括伊枯草菌素A、B、C、D和E,芽孢菌素D、F和L,以及抗霉枯草菌素等.其中,伊枯草菌素A的分子結構如圖3所示.

圖3 伊枯草菌素A分子結構圖
微生物產生的抗菌脂肽種類較多、結構較復雜、性質差異也較大.但是其中一部分脂肽化合物會有諸多類似物,且結構相類似,性質上也相近,因此在篩選及分離純化上存在困難,而且產量較低,這就給大規模的分離純化工藝帶來了麻煩.
大部分的芽孢桿菌都能分泌具有抗菌活性的物質,主要包括表面活性素、桿菌霉素、乳酸鏈球菌肽、豐原素和伊枯草菌素等(見表1).對脂肽的篩選鑒定及其活性探索也主要集中于芽孢桿菌類,霉菌和乳酸菌的研究次于芽孢桿菌,且這些能產脂肽物質的菌株主要是革蘭氏陽性菌,有的具有廣譜抗菌性,一部分只具有抗真菌特性等.
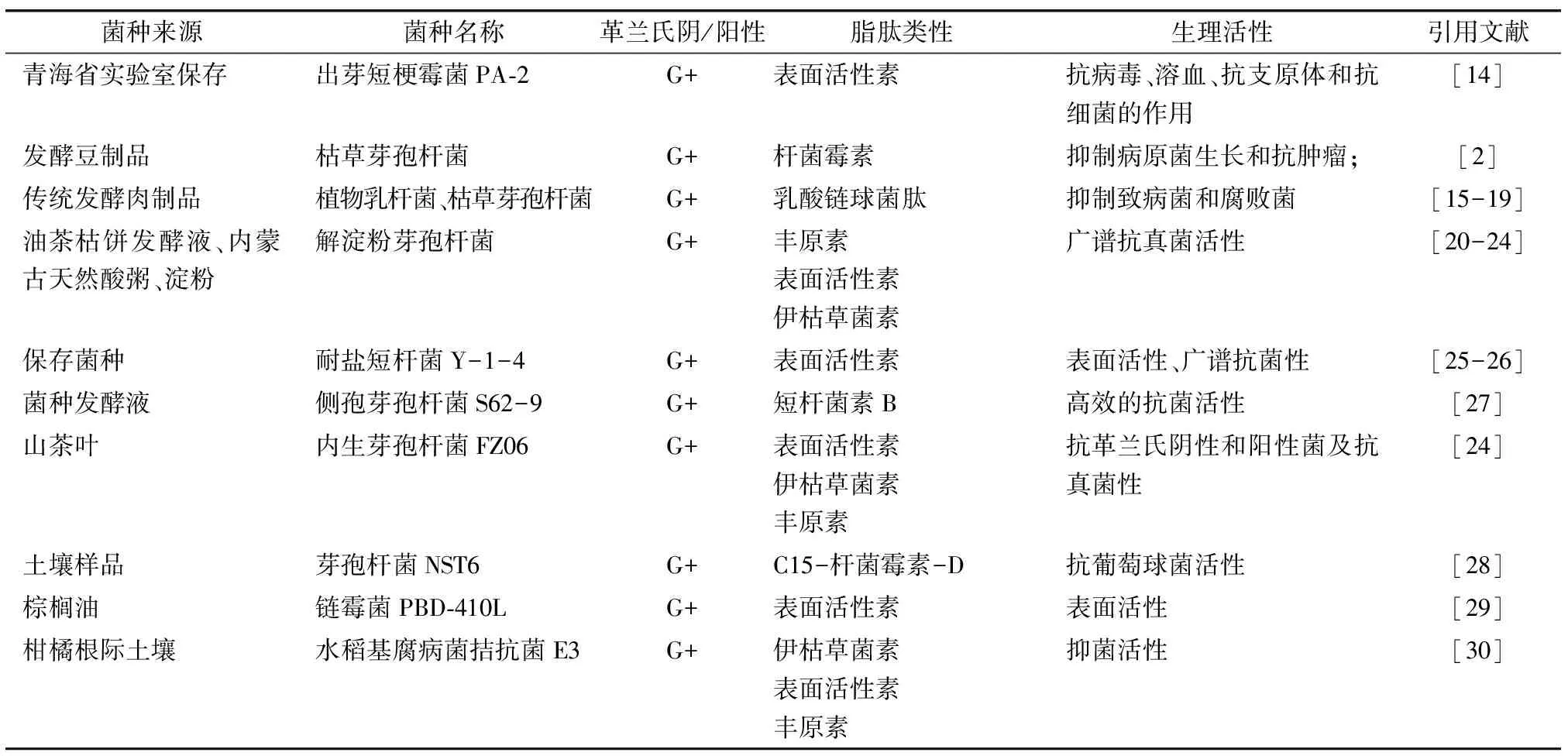
表1 部分脂肽及微生物類型及活性概述
1.2 抗菌脂肽的作用機制
1.2.1 抗菌脂肽與細菌細胞壁的作用
抗菌脂肽通過穿透細菌磷脂膜外面的細胞壁來抑制細菌的生長,借助其靜電引力來穿過細胞壁,抗菌脂肽與細胞壁上多糖之間的強靜電作用,與脂質之間的疏水作用以及脂質與抗菌脂肽間的氫鍵增多也會使抗菌脂肽活性增強,進而提升其抑菌率[31].例如,人源抗菌脂肽LL-37和抗菌脂肽SMAP-29被用來抑制肺病原菌,研究發現,其作用時主要借助抗菌脂肽與細菌細胞壁上多糖之間的相互作用而影響其生物學活性[32].其中,莢膜多糖主要存在于周圍介質中,在靜電引力下,與細菌膜外面的脂多糖帶相結合后,抗菌脂肽就會引起膜結構的異常變化,利用其較強的親和力能從帶有負電荷的脂多糖處轉移一部分陰離子,造成該部位的細胞壁紊亂,改變其穩定性后,抗菌脂肽便能穿過該區域來達到抑制細菌的目的.對于革蘭氏陽性菌來說,細胞壁表面的磷壁酸本身帶有的負電荷,可與抗菌脂肽的正電荷借助靜電引力并與之結合,使抗菌脂肽能夠順利通過革蘭氏陽性菌的細胞壁,再靠近細胞膜,并對細胞膜發揮作用[11].
1.2.2 抗菌脂肽與細菌細胞膜的作用
從抗菌脂肽鹵素-1與各種膜模型發生相互作用而引起的脂肽結構變化,可以得出肽模式的作用機制,該抗菌脂肽氨基酸序列在特定位置修飾,會導致兩親性的增加,蛋白質插入受限,進而致使抗菌性及溶血活性發生變化,導致對模型膜的不同反應[33].抗菌脂肽的作用方式的機制涉及脂肽與細胞膜之間膜孔的形成,抗菌脂肽與膜的相互作用在其活性機制中起到關鍵作用,并且抗菌脂肽主要以地毯式排列于脂質與水的表面.有抗菌脂肽存在時,脂質體的負電位降低,多肽的插入擾亂了有序排列的脂質,同時破壞細菌的細胞膜,降低其有效參數,便于水分子滲透過膜,并使得胞漿內物質流出,引起靶細胞的死亡[34].
但由于存在靜電排斥,陽離子抗菌脂肽不會與陽離子兩親體相互作用,而只會和陽離子聚集體系相互作用,不與陽離子單體發生作用[35].抗菌脂肽在高度疏水的介質中趨于α-螺旋,而在水介質中則采用隨機螺旋結構,螺旋度對抗菌脂肽與膜的親和力有影響,并且脂質囊泡可以促使肽排列成更多的螺旋結構[36].但這種螺旋結構的疏水相互作用很弱,需要具有大面積的聚集態來形成跨膜離子通道[27],由于肽鏈的疏水性會隨著肽鏈的縮短而延長,且與抗真菌活性間為協同作用,因此雙頭脂肽和脂三角相比脂五肽就會形成更小的、更具動態的聚集體,作為膠束或單體的陰離子兩親體與抗菌脂肽因為靜電作用,膜電勢遭到破壞,脂肽構象變化,最后導致細胞質膜分離和細胞質物質的損失,進而無法維持正常的生命活動而死亡[37].
1.2.3 抗菌脂肽與細菌胞質內靶目標的作用
抗菌脂肽與膜的結合不是直接將細胞殺死,而是其進入細胞后攻擊靶細胞的一個必要過程.先通過細胞壁和膜進入細胞體內,后出現核物質濃縮而發生異常間隔,進而殺死細胞.抗菌脂肽的抑菌機制因為其代謝產物的多樣性而多種多樣,如芽孢桿菌菌株w176產生的細胞上清液(CFS),有多種不同的代謝物,包括大環內酯抗生素、抗霉枯草菌素、聚酮合酶抗生素和表面活性素.當青霉菌與CFS共同培育后,其亞細胞結構發生了液泡化變化.用CFS處理馬尾松后,核糖體生物會脅迫相關基因轉錄水平發生變化,分析得出CFS是一種新的可用于防治柑橘綠霉病的生物制劑[38].除此之外,一些枯草芽孢桿菌能夠產生纖維素酶、分泌蛋白酶及脂肽類抑菌物質,這些抑菌物質會使病原菌的菌絲產生膨大畸形,從而抑制病原真菌的生長[39],并且這些產生抑菌物質的微生物和病原菌之間會存在對養分和空間的競爭,利用自身生長發育產生拮抗劑來盡可能地消耗可以利用的營養物質,從而抑制病原體的生長[40].其中,抗性誘導系統是一種常見的抑菌機制,用拮抗劑來誘導宿主菌的防御反應,促使目標基因的表達,進而激活不同的化學物質,進行防治感染[41-42].
2 抗菌脂肽類微生物源
2.1 芽孢桿菌屬
枯草芽孢桿菌是一類耐酸堿、耐熱、好氧或兼性好氧并產芽孢的革蘭氏陽性桿狀細菌[43].其分布廣泛、抗逆性強,且枯草芽孢桿菌含有適合用于合成抗生素的基因,一些菌株可以產生蛋白酶和脂肪酶等消化酶,合成多種生物活性物質及產生抗菌脂肽等[44-45].抗菌脂肽包括伊枯草菌素、表面活性素和豐原素3類[46],其產生抗菌物質分為兩個途徑:其一是通過非核糖體合成脂肽類抑菌物質,也是主要的抑菌物質之一[47];另外一種是通過核糖體合成蛋白類抑菌物質,這些蛋白類多是可以裂解細胞壁的酶類,導致細胞裂解,最后導致死亡[48].
胡美忠等[49]篩選到一株能產生抗菌脂肽的芽孢桿菌,且該抗菌脂肽抑菌譜廣,耐酸堿性和熱穩定性好.多粘類芽孢桿菌對致病菌和腐敗菌有顯著的抑制作用,該菌產生的抑菌物質為蛋白類的細菌素,目的是作用于有害菌的細胞壁及細胞膜,進而達到抑菌效果[50];地衣芽孢桿菌FJAT-4 產生的抗菌脂肽為表面活性素和豐原素兩大類物質,該抑菌脂肽會致使辣椒、香蕉和番茄尖孢鐮刀菌等多種植物致病菌,尤其是尖孢鐮刀菌的菌絲體發育畸形,從而影響該菌的正常生長[51].此外,甲基型芽孢桿菌產生的多種抑菌物質單獨使用和協同發揮的抑制使用效果不一,多種抑菌物質共同加入到含致病菌發酵液中進行發酵后,產生的上清液和沉淀里抑菌帶的明顯程度也有所差異,即表現出不同程度的抑菌活性,這為脂肽的抗菌機制的研究提供了依據,并且還能在發酵過程中產生胞外抑菌物質,對霉菌和致病菌都有一定的抑制作用,因此是一類具有潛在價值的微生物[52-53].
2.2 乳酸菌屬
乳酸菌是一種益生菌,因具有發酵產酸的特性,而被廣泛應用到酸奶、發酵面包及酵素飲料等食品中.乳酸菌還能產生一些抗菌脂肽及蛋白類物質,在食品的生物防腐等方面存在著重要的作用,在抗菌脂肽方面也存在極大的開發潛力.
依據抗菌脂肽的抑菌譜、抑菌機制和耐熱性等可以將抗菌脂肽分為以乳酸鏈球菌素為代表的羊毛硫抗菌脂肽、非羊毛硫抗菌脂肽、大分子耐熱性多聚肽以及含有多糖、陽離子和脂質的多肽這4大類[17].楊云喜等[18]篩選了一株產抗菌脂肽的乳酸菌,該抗菌脂肽對真菌無抑菌活性,但是對細菌具有廣譜抑菌性,通過優化能夠產生抗菌脂肽的培養基成分及培養條件,為其大規模地應用提供了基礎.趙乙楨等[19]從新鮮牛奶中分離到一株能產生廣譜抑菌性抗菌脂肽的鼠李糖乳桿菌ZJ225.趙冬兵等[54]從板鴨中篩選得到了一株魏斯氏菌,其產生的抗菌脂肽對較強酸堿都具有穩定性.章檢明等[17]在91株乳酸菌中篩選出了7株產生抗菌脂肽的乳酸菌,有魏斯氏菌屬、鼠李糖乳桿菌屬、葡萄球菌屬及微球菌屬4種,其中鼠李糖乳桿菌屬能產生具有廣譜抑菌性,且性質較穩定的多肽.
2.3 霉 菌
相比其他屬類的微生物而言,對霉菌產生抗菌物質的研究較少.有研究者探索了部分霉菌的抑菌活性,通過優化出芽短梗霉菌PA-2的發酵溫度、轉速及裝液量等,得出該菌產生的脂肽物質為表面活性劑,具有較好的發泡和乳化作用[14,55].李志田等[56]篩選鑒定出了一株渾圓鏈霉菌QH-16,經離體測定,其對多種細菌及植物病原菌具有很好的抑菌作用.張萍華等[57]探索了黑翅土白蟻巢中鏈霉菌發酵產物的抗菌活性,并對其抗菌成分進行研究,發現其對金黃色葡萄球菌具有很好的抑制作用,具有開發成為新型微生物源殺菌劑的潛力.
毛霉是一類高效微生物,能夠在單一培養基中利用可再生來源——碳源,來生產生物糖脂、生物乳化劑和表面活性劑等,毛霉在相同的培養基和不同的物理條件下實現了生物表面活性劑和乳化劑的生產.此外,可以肯定的是,生產的表面活性劑和乳化劑滿足可持續性和附加值的基本要求,因為它們是獲得各種工業部門日常生產的化合物的主要原料[58].
3 抗菌脂肽的應用
3.1 在食品中的應用
隨著社會的發展,消費者對食品安全的要求在不斷提高,但是濫用及違規使用添加劑等造成的添加劑類食品安全事故也時有發生.此外,還有食品自身的變化導致的污染,如黃曲霉,其寄主范圍非常廣,辣椒、玉米、棉花以及花生等多種作物都會被其污染,不僅會造成巨大的經濟損失,還在污染作物的過程中產生黃曲霉毒素,人和畜禽誤食后會引發中毒,嚴重的可能會致癌和導致死亡[59].在這樣的環境背景下,類似于抗菌脂肽等生物制劑得到了消費者的青睞.
抗菌脂肽在食品行業的主要應用是作為生物防腐保鮮劑和食品添加劑.抗菌脂肽的表面活性劑具有乳化作用,可用作食品加工中的發泡劑和乳化劑,以改善食品的質構、色澤和口感.另外,抗菌脂肽的抑菌活性又可作為防腐保鮮劑應用于食品中.張學雯等[60]分離鑒定了黃曲霉拮抗菌B10-6-1拮抗物種類,并對其結構等進行了研究,這為黃曲霉的有效控制提供了依據.
3.2 在飼料生產中的應用
隨著消費者對肉類的需求不斷增加,促進了各地養殖業的發展,但是由于養殖地分布各地,控制上就容易出現問題.抗生素飼料的濫用會導致動物體內的抗生素通過畜產品、水、土壤等傳遞給人類,導致人體產生大量耐藥菌株,對人類的健康造成危害.因此,綠色且安全、無殘留及不易產生耐藥性的新型生物飼料添加劑備受關注,已成為諸多科研工作者的研究方向.
抗菌脂肽能夠成為新型飼料與其結構有關,其結構是由易于消化吸收的脂肪酸鏈和氨基酸肽鏈兩部分組成.抗菌脂肽可以部分取代用于飼喂的抗生素,避免抗生素在動物體內過多蓄積而沿食物鏈危害人體健康.另外,抗菌脂肽在抑制革蘭氏陽性菌和部分革蘭氏陰性菌的同時,又能夠促進其生長,且此過程不會被腸道吸收,只在消化道內起作用,因此一般無殘留,效果穩定,也不易產生細菌耐藥性[11].
3.3 在醫藥行業的應用
為了治療細菌感染,抗生素被大量應用,其作用效果是通過破壞細菌的DNA、細胞膜和細胞活性以及用其來結合細菌的一些化合物來殺死細菌.抗菌脂肽能與兩親性生物可降解聚合物自行組裝而成,形成的HnMc膠束能有效地結合細菌膜[61],從而殺滅廣譜細菌[62].抗菌脂肽因為菌源不同,以及自身結構的不同導致了抗菌性能的差異.其中,肽鏈中脂肪酸長度的改變導致伊枯草菌素具有多樣性,可以防止多種生物性植物病原菌,豐原素具有治療皮膚真菌病及具抗真菌的作用[63-64].
抗菌脂肽因其抗黏附性能,可以抑制生物膜的產生和細菌黏附在感染部位的能力[1],因此利用這種能力生產抗生素和治療性疫苗,可以治療革蘭氏陽性菌及病毒的感染.有一種環狀類型的抗菌脂肽,由非極性的脂肪酸鏈通過結合極性的氨基酸脂肪鏈組成,其在多重耐藥病原細菌方面具有獨特的抑菌能力.正是因為代謝產生的脂肽能夠較強地抑制革蘭氏陽性菌的生長,如抑制結核分枝桿菌與肺炎雙球菌等細菌,可應用于疫苗的生產中[6].有研究表明,含有抗菌脂肽的蛋白質與不含抗菌脂肽的比較,在無外部佐劑的情況下能夠誘導強烈的病毒跨膜蛋白的特異性抗體反應,此類重組基因的蛋白質有望成為一種疫苗[65].脂肽在國外已被應用于預防和治療藥物,甚至有研究表明,微生物脂肽還能在預防癌癥方面有所應用.
3.4 在農業中的應用
現在農業上受到多種病蟲害的威脅,不僅影響了農產品的質量,還大大降低了農產品的產量和農戶的收入[66].目前,大部分種植者的觀念仍是化學防治效果顯著,因此廣泛應用化學物質來防治農業病蟲害,但是,這些化學用品可能會對生態環境和人類生活產生負面影響.因此,近年來生物防治被廣大研究者所推崇,被認為是一種安全和可持續的方法,其中芽孢桿菌能夠產生一些抗菌脂肽類物質,具有廣譜抗菌活性,因此被廣泛用于農業的生物防治[67].枯草芽孢桿菌已被選為脂肽類抗生素的生物農藥的菌株之一,它能產生對煙草病毒及辣椒病毒有防治效果的脂肽類抗生素表面活性素[46],能用于控制柑橘綠色和藍色霉菌[68],以及防治辣椒病害、黃瓜病害及油菜菌核病[26].另外,解淀粉芽孢桿菌產生的有抑菌作用的活性物質C14-桿菌霉素-D和C15-桿菌霉素-D,可以用于合成新型生物農藥,以解決黃曲霉污染防治的瓶頸問題[59].
4 討論與展望
現在已經發現的產生抗菌脂肽芽孢桿菌種類頗多,但其他屬類的研究較少.另外抗菌脂肽的同系物種類繁多,其發酵液成分復雜,很難大規模生產和濃縮純化出足量的純品,甚至許多產物還沒有一種標準的檢測方法.因此,為了更加高效地利用這些微生物來產生抗菌物質,應該對其結構進行深入研究,才能找到提高其產量的方法[55].從目前來看,脂肽微生物的研究可以從以下幾方面進行:
1)抗菌脂肽的抑菌機制仍然主要是對細胞壁、細胞膜及靶細胞的作用,可以根據其結構進行更深入研究,探索其具體的基因作用位點、抑菌機制和應用范圍等.可以通過從基因水平來探究抗菌脂肽的結構,目的是要獲得相關的轉錄調控因子,如用自殺載體等方式來獲得突變菌株,抗菌脂肽產量會有所下降,再控制相關的正向調控因子發揮正調控作用,對突變基因進行回補后,脂肽產量會有所恢復[69].另外,還存在阻遏蛋白對脂肽進行調控,當分別含有兩種阻遏蛋白的菌株,其控制基因突變后再利用載體回補,這兩種控制基因中的任意一個發生突變,另一個阻遏蛋白的表達量有所上調,說明這兩種阻遏蛋白相互抑制,在整個系統中協調發揮作用,基因突變后阻遏蛋白表達量的增加伴隨著抗菌脂肽產量的增加,為正調控因子,阻遏蛋白的增加伴隨著抗菌脂肽產量的減少為負調控,正向調控的蛋白可以抑制阻遏蛋白而起到正向調控的作用[70].因此,產脂肽微生物的篩選鑒定及脂肽的分離純化方面可以從基因的調控入手,縮小篩選范圍、提高實驗效率.
2)抗菌脂肽在應用上還存在一些問題,如黃曲霉毒素容易污染食品而造成食品安全問題,盡管目前為止己篩選到很多抑制黃曲霉生長的化合物,但幾乎沒有應用到實踐中來,因此需要注意與實踐相結合,拓寬應用范圍[71].
3)將抗菌脂肽用于臨床上還存在許多問題,抗菌脂肽本質是蛋白質,應用過程中容易被降解,需要包埋或修飾處理,而且在毒力學和藥效學等方面研究較少,需要進一步的探討脂肽類物質對人體的安全問題的影響,提供一些支持性的研究報告.
4)基因工程是抗菌脂肽的主要獲得手段,其成本較高,生產效率較低,因此應該加大科學研究,如優化發酵條件、工藝設計,改良菌種、載體及降低成本等,逐漸向研發出性能更好、規模更大的生物制劑的方向發展[72].
