不同化學(xué)藥劑處理對(duì)休眠期大葉樟種子萌發(fā)的影響
2021-07-09 02:03:29張麗華韓浩章李素華王曉立王芳趙榮蔣亞華
天津農(nóng)業(yè)科學(xué) 2021年4期
張麗華 韓浩章 李素華 王曉立 王芳 趙榮 蔣亞華
摘? ? 要:為研究大葉樟種子休眠的解除措施,以新采大葉樟種子為材料,研究不同濃度赤霉素(GA3)、過氧化氫(H2O2)、褪黑素(MT)、水楊酸(SA)處理對(duì)其萌芽特性和幼苗生長的影響,每種化學(xué)藥劑設(shè)置3個(gè)濃度梯度,以蒸餾水處理為對(duì)照。結(jié)果表明,同一種化學(xué)藥劑中,2%濃度的H2O2,300 mg·L-1濃度的 GA3,200 μg·L-1濃度的MT,0.5 mmol·L-1濃度的SA處理能顯著提高休眠期大葉樟種子萌發(fā)率、萌發(fā)勢(shì)、萌芽指數(shù)和萌發(fā)后的幼苗質(zhì)量,濃度過高或過低則效果變差。結(jié)合隸屬函數(shù)法的綜合評(píng)價(jià)值來看,200 μg·L-1的MT和300 mg·L-1的GA3處理效果較好,綜合隸屬函數(shù)值分別為0.805,0.761,而500 μg·L-1的MT和2 mmol·L-1的SA處理效果較差,綜合隸屬函數(shù)值分別為0.113,0.025。綜上,適宜濃度的GA3、MT、H2O2和SA處理均能促進(jìn)大葉樟種子休眠解除和提高種苗質(zhì)量,其中以200 μg·L-1的MT和300 mg·L-1的GA3處理3 h效果最好。
關(guān)鍵詞:化學(xué)藥劑處理;休眠期;大葉樟種子;萌發(fā)
中圖分類號(hào):S792.23? ? ? ? ?文獻(xiàn)標(biāo)識(shí)碼:A? ? ? ? ? DOI 編碼:10.3969/j.issn.1006-6500.2021.04.001
Abstract: In order to study the measures to relieve the dormancy of Cinnamomum septentrionale seeds, the effects of different concentrations of GA3, hydrogen peroxide(H2O2), melatonin(MT) and salicylic acid(SA) treatment on the germination characteristics and seedling growth were studied. Three concentration gradients were set up for each chemical agent, and the treatment of distilled water was used as the control. The results showed that in the same chemical agent treatments, the germination rate, germination potential, germination index and seedling quality of Cinnamomum septentrionale seeds in dormant stage were significantly improved by the optimal concentration, such as? 2% H2O2, 300 mg·L-1 GA3, 200 g·L-1 MT, and 0.5 mmol·L-1 SA, while the effect was worse if the concentration was too high or too low. Combined with the results of the membership function method,? 200 μg·L-1 MT and 300 mg·L-1 GA3 had? better treatment effect, with a synthetic membership function value of 0.805 and 0.761 respectively, while? 500 μg·L-1 MT and 2 mmol·L-1 SA had worse treatment effect, with a synthetic membership function value of 0.113 and 0.025 respectively. In conclusion, suitable concentrations of GA3, MT, H2O2 and SA could promote the dormancy dissolution and improve the quality of seedlings, which the seeds soaked 3 h with MT of 200 g·L-1 and GA3 of 300 mg·L-1 had the best effect.
Key words: chemical treatment; dormant; Cinnamomum septentrionale seed; germination
大葉樟(Cinnamomum septentrionale Hand. Mazz)又稱銀木,為樟科樟屬常綠喬木,樹形美觀,滯塵能力強(qiáng),園林綠化中常用作行道樹、孤植樹和林用樹種,生態(tài)效益明顯,已成為長江南北地區(qū)城市綠化的主導(dǎo)品種。樟屬植物種苗生產(chǎn)在園林產(chǎn)業(yè)中占有重要地位,種子繁殖仍然是植物育苗的主要方式,但樟屬植物具有生理休眠特性,未通過休眠的種子往往發(fā)芽率極低[1]。低溫層積通常用以保存樟屬植物種子并解除休眠,但種子在低溫層積處理后的萌芽率仍然不高,仍會(huì)因種皮透氣性差和存在抑制物質(zhì)而不萌發(fā)或萌發(fā)期延長,甚至在2~3 年后還在陸續(xù)出苗,種苗質(zhì)量變差[2]。如若將楠木(Phoebe bournei)種子去皮,則可比未去皮的種子早萌發(fā)30 d,種苗質(zhì)量也顯著提高[3],但種子去皮程序繁雜,不適于種苗生產(chǎn)。赤霉素(GA3)[1]、過氧化氫(H2O2)[4]、褪黑素(melatonin, MT)[5]、水楊酸(salicylic acid, SA)[6]等常用于促進(jìn)植物種子萌發(fā),并取得較好的效果,但在樟屬植物種子休眠解除中應(yīng)用較少。張靜靜等[7]在低溫層積后采用硫酸處理破壞種皮過厚對(duì)香樟種子萌發(fā)的限制,促進(jìn)萌芽;梁機(jī)等[8]采用濕沙包埋4 ℃保存結(jié)合80 mg·L-1 GA3浸泡12 h處理使樟樹種子發(fā)芽率達(dá)到89%;肖祖飛等[1]采用陰干保存結(jié)合400 mg·L-1的GA3處理后使萌芽率達(dá)49%,但與低溫層積的差異不顯著,若單獨(dú)用0 ℃低溫保存處理可使香樟種子萌發(fā)率可提高至69%,但幼苗質(zhì)量較差。因而,樟屬植物種子休眠解除措施目前均是以低溫層積處理為前提,種子損耗量大,外源化學(xué)藥劑處理和溫水處理等方式進(jìn)行催芽的效果也不盡一致。基于此,本研究以新采大葉樟種子為材料,研究不同化學(xué)藥劑處理對(duì)其萌芽特性和幼苗生長質(zhì)量的影響,以期為大葉樟種子休眠機(jī)理研究和種苗生產(chǎn)提供參考。
1 材料和方法
1.1 試驗(yàn)材料
試驗(yàn)用種子采自宿遷學(xué)院樟屬植物資源圃,于2019年11月15日采收,去皮后用清水處理陰干,再用0.2%的KMnO4浸種15 min進(jìn)行消毒處理,備用。
1.2 處理方法
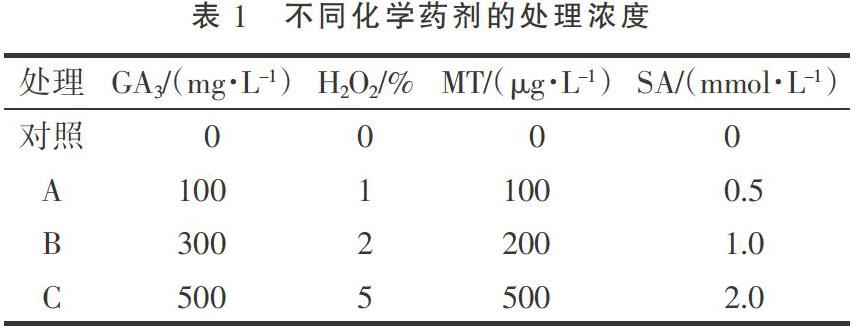
將待測(cè)種子分16個(gè)處理,每處理100粒,以蒸餾水為對(duì)照,分別采用不同濃度GA3、H2O2、MT、SA(表1,不同濃度化學(xué)藥劑處理統(tǒng)一標(biāo)記為A、B、C)在培養(yǎng)皿中進(jìn)行浸種處理3 h,浸種時(shí)將種子沒入溶液以下,浸種后用清水清洗,陰干后進(jìn)行播種。每個(gè)處理設(shè)3個(gè)重復(fù)。
2019年12月1日開始播種,播種基質(zhì)為草炭土∶蛭石∶田園土=3∶1∶2,營養(yǎng)缽高10 cm,上口直徑8 cm,5粒·缽-1。播種后置于光照培養(yǎng)箱中進(jìn)行培養(yǎng)(白天13 h,溫度30±2 ℃,濕度60%~70%;晚上11 h,溫度18±2 ℃,濕度50%~60%),正常水分管理。定期記錄其萌芽數(shù)量,根據(jù)萌芽數(shù)量分別計(jì)算其萌芽率、萌芽勢(shì)和萌芽指數(shù),90 d后,每個(gè)重復(fù)隨機(jī)采取3株幼苗,檢測(cè)幼苗株高、葉片數(shù)量、莖粗、地上部干質(zhì)量和地下部干質(zhì)量,取平均值進(jìn)行統(tǒng)計(jì)分析。
1.3 試驗(yàn)方法
大葉樟種子萌芽數(shù)量統(tǒng)計(jì)時(shí)以上胚軸伸出表土2 mm記為發(fā)芽,萌芽勢(shì)為40 d萌芽種子總數(shù)占種子總數(shù)的比值,萌芽率為90 d萌芽種子總數(shù)占種子總數(shù)的比值,萌芽指數(shù)=∑(Gt/Dt)(Dt為萌發(fā)天數(shù),Gt為 t日的萌芽種子數(shù), 即與Dt相對(duì)應(yīng)的萌芽種子數(shù));株高采用直尺測(cè)定,莖粗采用游標(biāo)卡尺測(cè)定,地上部干質(zhì)量和地下部干質(zhì)量采用烘干法,于105 ℃烘箱中烘干至恒質(zhì)量。化學(xué)藥劑解除大葉樟種子休眠效果綜合評(píng)價(jià)采用隸屬函數(shù)法[9],在所有測(cè)定指標(biāo)中,萌芽率、萌芽勢(shì)、萌芽指數(shù)、幼苗株高、葉片數(shù)量、莖粗、地上部干質(zhì)量、地下部干質(zhì)量均與解除大葉樟種子休眠效果成正相關(guān),采用隸屬函數(shù)公式:
U(Xab)=(Xab-Xamin)/(Xamax- Xamin)
式中,U(Xab)為測(cè)定指標(biāo)的隸屬函數(shù)值;Xab為各處理的指標(biāo)測(cè)定值;Xamin為各處理指標(biāo)測(cè)定的最小值;Xamax為各處理指標(biāo)測(cè)定的最大值。
1.4 數(shù)據(jù)處理與分析
采用Microsoft Excel 2010進(jìn)行制表,采用SPSS 21.0進(jìn)行數(shù)據(jù)分析和隸屬函數(shù)值計(jì)算,采用最小顯著差異法(LSD)進(jìn)行數(shù)據(jù)顯著性差異分析。
2 結(jié)果與分析
2.1 不同化學(xué)藥劑處理對(duì)休眠期大葉樟種子萌芽率的影響
由表2可以看出,與對(duì)照相比,適宜濃度的GA3、SA、MT、H2O2處理均能提高休眠期大葉樟種子的萌芽率,但不同化學(xué)藥劑處理的效果不同。其中,100,300 mg·L-1的GA3處理比對(duì)照顯著提高了5.33,11.67百分點(diǎn)(P<0.05),2%濃度的H2O2處理比對(duì)照顯著提高了13.33百分點(diǎn)(P<0.05),100,200 μg·L-1的MT處理比對(duì)照顯著提高了33.67,15.33百分點(diǎn)(P<0.05);300 mg·L-1的GA3處理、1%和5%濃度的H2O2處理及0.5 mmol·L-1的SA處理與對(duì)照差異不顯著;500 μg·L-1的MT處理及1,2 mmol·L-1的SA處理分別比對(duì)照顯著降低了21.67,14.33,16.33百分點(diǎn)(P<0.05)。
2.2 不同化學(xué)藥劑處理對(duì)休眠期大葉樟種子萌芽勢(shì)的影響
由表3可以看出,與對(duì)照相比,適宜濃度的GA3、SA、MT、H2O2處理均能提高休眠期大葉樟種子的萌芽勢(shì),但不同化學(xué)藥劑處理的效果不同。其中,100,300,500 mg·L-1的GA3處理比對(duì)照顯著提高了13.67,2.00,10.34百分點(diǎn)(P<0.05),1%,2%,5%濃度的H2O2處理比對(duì)照顯著提高了11.00,14.67,5.34百分點(diǎn)(P<0.05),100,200 μg·L-1的MT處理比對(duì)照顯著提高了18.67,13.34百分點(diǎn)(P<0.05),0.5 mmol·L-1的SA處理比對(duì)照顯著提高了9.67百分點(diǎn)(P<0.05);而500 μg·L-1的MT處理與對(duì)照差異不顯著,1,2 mmol·L-1的SA處理比分別對(duì)照顯著降低了3.33,4.66百分點(diǎn)(P<0.05)。
2.3 不同化學(xué)藥劑處理對(duì)休眠期大葉樟種子萌芽指數(shù)的影響
由表4可以看出,與對(duì)照相比,適宜濃度的GA3、SA、MT、H2O2處理均能提高大葉樟種子的萌芽指數(shù),但不同化學(xué)藥劑處理的效果不同。其中,100,300,500 mg·L-1的GA3處理比對(duì)照顯著提高了55.16%,78.81%,48.72%(P<0.05),1%,2%,5%濃度的H2O2處理比對(duì)照顯著提高了54.29%,59.54%,27.63%(P<0.05),100,200 μg·L-1的MT處理比對(duì)照顯著提高了66.26%,58.18%(P<0.05),0.5 mmol·L-1的SA處理比對(duì)照顯著提高了53.88%(P<0.05);而500 μg·L-1的MT處理比對(duì)照顯著降低了93.31%(P<0.05),1,2 mmol·L-1的SA處理分別比對(duì)照顯著降低了29.28%,172.29%(P<0.05)。
2.4 不同化學(xué)藥劑處理對(duì)大葉樟幼苗株高的影響
由表5可以看出,與對(duì)照相比,適宜濃度的GA3、SA、MT、H2O2處理均能提高大葉樟幼苗株高,但不同化學(xué)藥劑處理的效果不同。其中100,300,500 mg·L-1的GA3處理比對(duì)照顯著提高了56.82%,90.43%,71.08%(P<0.05),1%,2%,5%濃度的H2O2處理比對(duì)照顯著提高了61.91%,47.66%,29.33%(P<0.05),100,200 μg·L-1的MT處理比對(duì)照顯著提高了74.13%,70.06%(P<0.05),0.5 mmol·L-1的SA處理比對(duì)照顯著提高了40.84%(P<0.05);而500 μg·L-1的MT處理及1,2 mmol·L-1的SA處理與對(duì)照差異不顯著。
2.5 不同化學(xué)藥劑處理對(duì)大葉樟幼苗葉片數(shù)量的影響
由表6可以看出,與對(duì)照相比,適宜濃度的GA3、SA、MT、H2O2處理均能提高大葉樟幼苗葉片數(shù)量,但不同化學(xué)藥劑處理的效果不同。其中,100,300,500 mg·L-1的GA3處理比對(duì)照顯著提高了61.76%,41.18%,26.47%(P<0.05),1%,2%,5%濃度的H2O2處理比對(duì)照顯著提高了32.35%,38.24%,41.18%(P<0.05),100,200 μg·L-1的MT處理比對(duì)照顯著提高了44.12%,41.18%(P<0.05),0.5,1 mmol·L-1的SA處理比對(duì)照顯著提高了39.71%,17.61%(P<0.05);而500 μg·L-1的MT處理和2 mmol·L-1的SA處理與對(duì)照差異不顯著。
2.6 不同化學(xué)藥劑處理對(duì)大葉樟幼苗莖粗的影響
由表7可以看出,與對(duì)照相比,適宜濃度的GA3、SA、MT、H2O2處理均能提高大葉樟幼苗葉片數(shù)量,但不同化學(xué)藥劑處理的效果不同。其中,300,500 mg·L-1的GA3處理比對(duì)照顯著提高了26.51%,37.35%(P<0.05),1%,2%,5%濃度的H2O2處理比對(duì)照顯著提高了37.35%,48.19%,46.99%(P<0.05),100,200 μg·L-1的MT處理比對(duì)照顯著提高了20.48%,46.99%(P<0.05),0.5 mmol·L-1的SA處理比對(duì)照顯著提高了31.33%;而2 mmol·L-1的SA處理顯著低于對(duì)照18.01%(P<0.05),100 mg·L-1的GA3處理、500 μg·L-1的MT處理和1 mmol·L-1SA處理與對(duì)照差異不顯著。
2.7 不同化學(xué)藥劑處理對(duì)大葉樟幼苗地上部干質(zhì)量的影響
由表8可以看出,與對(duì)照相比,適宜濃度的GA3、SA、MT、H2O2處理均能提高大葉樟幼苗地上部干質(zhì)量,但不同化學(xué)藥劑處理的效果不同。其中,100,300,500 mg·L-1的GA3處理比對(duì)照顯著提高了31.14%,64.56%,97.09%(P<0.05),1%,2%,5%濃度的H2O2處理比對(duì)照顯著提高了22.47%,38.87%,53.09%(P<0.05),100,200 μg·L-1的MT處理比對(duì)照顯著提高了55.22%,108.87%(P<0.05),0.5 mmol·L-1的SA處理比對(duì)照顯著提高了28.96%(P<0.05);2 mmol·L-1的SA處理顯著低于對(duì)照12.14%(P<0.05),500 μg·L-1的MT和1 mmol·L-1SA處理的與對(duì)照差異不顯著。
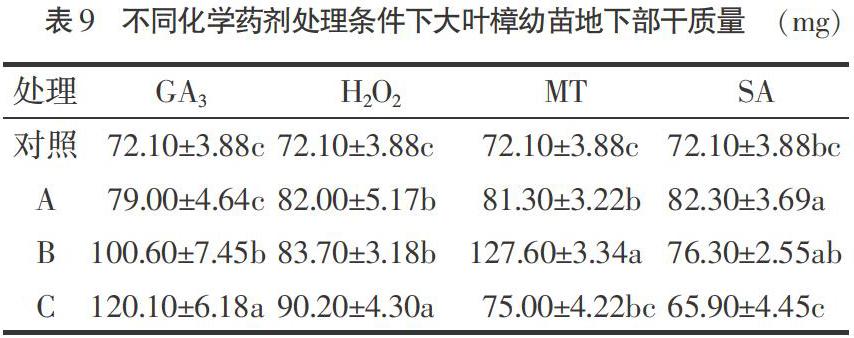
2.8 不同化學(xué)藥劑處理對(duì)大葉樟幼苗地下部干質(zhì)量的影響
由表9可以看出,與對(duì)照相比,適宜濃度的GA3、SA、MT、H2O2處理均能提高大葉樟幼苗地上部干質(zhì)量,但不同化學(xué)藥劑處理的效果不同。其中,300,500 mg·L-1的GA3處理比對(duì)照顯著提高了39.63%,66.57%(P<0.05),1%,2%,5%濃度的H2O2處理比對(duì)照顯著提高了13.73%,16.09%,25.10%(P<0.05),100,200 μg·L-1的MT處理比對(duì)照顯著提高了12.76%,76.98%(P<0.05),0.5 mmol·L-1的SA處理比對(duì)照顯著提高了14.15%(P<0.05);100 mg·L-1的GA3、500 μg·L-1的MT及1,2 mmol·L-1的SA處理與對(duì)照差異不顯著。
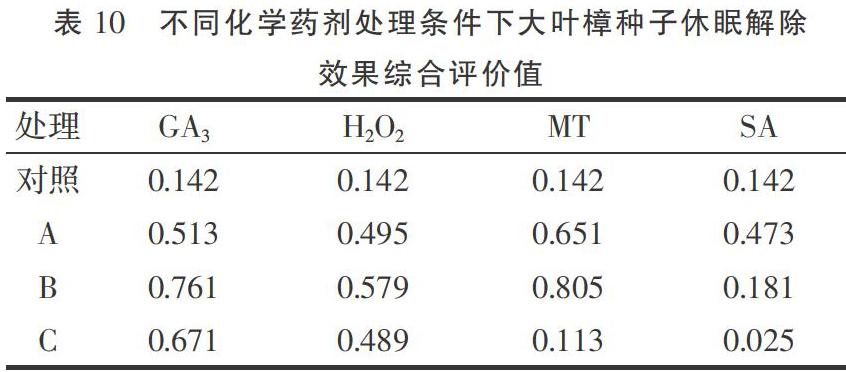
2.9 不同化學(xué)藥劑處理對(duì)大葉樟種子休眠解除效果的綜合評(píng)價(jià)
綜合評(píng)價(jià)值即綜合隸屬函數(shù)值(簡稱“D值”)表示不同化學(xué)藥劑對(duì)大葉樟種子休眠解除的綜合效果,D在0~1之間,D值越接近1表示化學(xué)藥劑解除大葉樟種子休眠的效果越好,反之則效果越差[14]。由表10可以看出,不同化學(xué)藥劑處理對(duì)香樟種子休眠解除的綜合效果不一致,其中200 μg·L-1的MT和300 mg·L-1的GA3處理效果較好,D值分別為0.805,0.761;其次為100 μg·L-1的MT和100 mg·L-1的GA3處理,D值分別為0.651,0.671;再者是2%濃度的H2O2和100 mg·L-1的GA3處理,D值分別為0.579,0.513;500 μg·L-1的MT和2 mmol·L-1的SA處理效果較差,D值分別為0.113,0.025,均低于對(duì)照。
3 結(jié)論與討論
休眠是種子植物在長期進(jìn)化過程中產(chǎn)生的適應(yīng)性性狀,植物通過抑制種子萌發(fā)保證其在不利環(huán)境中生存[10],如若種子不能適時(shí)解除休眠,則播種后表現(xiàn)為萌芽率低、萌芽推遲、萌發(fā)后種苗生長質(zhì)量差、甚至在萌發(fā)后死亡等現(xiàn)象。研究認(rèn)為,新采收的香樟種子胚在離體條件下可正常萌發(fā),屬非深度生理休眠,種皮過厚和種子內(nèi)的抑制物質(zhì)是引起香樟種子休眠的主要原因[1-2]。本試驗(yàn)中,未解除休眠的大葉樟種子萌發(fā)率、萌發(fā)勢(shì)和萌芽指數(shù)均低于適宜濃度化學(xué)藥劑處理的,萌發(fā)后的幼苗質(zhì)量較差;采用不同濃度的H2O2、GA3、MT、SA處理能影響大葉樟種子休眠解除和幼苗質(zhì)量,同一種化學(xué)藥劑中,2%濃度的H2O2、300 mg·L-1濃度的 GA3、200 μg·L-1濃度的MT、0.5 mmol·L-1濃度的SA處理能顯著提高休眠期大葉樟種子的萌芽率、萌芽勢(shì)、萌芽指數(shù)和種苗質(zhì)量,濃度過高或過低則效果變差,因而化學(xué)藥劑處理能一定程度上解除大葉樟種子休眠,但具有濃度效應(yīng)。
已有研究普遍認(rèn)為ABA和GA是拮抗調(diào)節(jié)種子休眠和萌發(fā)的主要激素,隨著種子體內(nèi)ABA含量的不斷積累,種子出現(xiàn)并維持休眠狀態(tài),當(dāng)種子內(nèi)部ABA含量下調(diào)、GA含量上調(diào)時(shí),種子開始萌發(fā)[11]。若種子內(nèi)部缺少GA,則萌發(fā)受到抑制,只有外源施加GA后種子才能萌發(fā)[12],外源GA3處理主要通過加速種子內(nèi)可溶性蛋白質(zhì)、淀粉等貯藏物質(zhì)的轉(zhuǎn)化和利用[13],激活葡萄糖轉(zhuǎn)運(yùn)蛋白功能[14],啟動(dòng)并加速糖酵解和TCA循環(huán)[15],從而增加胚的生長勢(shì),克服種子覆蓋層的機(jī)械抑制[16],促進(jìn)種苗生長。本試驗(yàn)中,300 mg·L-1濃度的GA3處理顯著提高休眠期大葉樟種子的萌芽率、萌芽勢(shì)、萌芽指數(shù)和種苗質(zhì)量,促進(jìn)種子休眠解除,與前人的研究結(jié)果[17-19]一致。據(jù)報(bào)道,化學(xué)藥劑處理調(diào)控種子的休眠和萌發(fā)與ABA或GA通路密切相關(guān)[17-18]。外源H2O2能增強(qiáng)休眠期植物種子內(nèi)CYP707A基因的表達(dá)加快ABA的分解代謝,增強(qiáng)GA生物合成基因如GA33ox和GA320ox的表達(dá)促進(jìn)GA的生物合成[19],外源MT和SA在高鹽等逆境條件下處理植物種子也有類似的現(xiàn)象[6,20]。本試驗(yàn)中,2%濃度的H2O2處理顯著提高休眠期大葉樟種子的萌芽率、萌芽勢(shì)、萌芽指數(shù)和種苗質(zhì)量,促進(jìn)種子休眠解除,但當(dāng)H2O2處理濃度增加后,種子內(nèi)部ROS積累量超過一定閾值,種子生活力和萌芽率隨之降低,ROS只有在一定濃度范圍內(nèi)才能誘導(dǎo)種子萌發(fā)[21],H2O2解除種子休眠可能還與抗氧化酶系統(tǒng)等代謝途徑有關(guān)。另外,200 μg·L-1濃度的MT處理也能顯著提高休眠期大葉樟種子的萌芽率、萌芽勢(shì)、萌芽指數(shù)和種苗質(zhì)量,促進(jìn)種子休眠解除,其D值為0.805,高于其他處理,其原因可能是大葉樟為油料種子,外源MT能顯著上調(diào)乙醛酸循環(huán)中關(guān)鍵酶異檸檬酸裂解酶和蘋果酸合酶的活性,從而調(diào)控乙醛酸循環(huán),促進(jìn)糖酵解和三羧酸循環(huán)[22],促進(jìn)種子呼吸代謝,并增加抗氧化酶相關(guān)基因的表達(dá)以增強(qiáng)對(duì)活性氧的清除能力[23],而0.5 mmol·L-1濃度SA處理雖能顯著提高休眠期大葉樟種子的萌芽勢(shì)、萌芽指數(shù)和種苗質(zhì)量,但對(duì)萌芽率影響不顯著,其D值為0.472,且隨著SA處理濃度的提高,D值降低,2 mmol·L-1濃度SA處理時(shí)的D值為0.025,遠(yuǎn)低于對(duì)照,說明SA處理對(duì)大葉樟種子休眠解除的效果總體較差。另有研究認(rèn)為,SA僅在高鹽、干旱和低溫等逆境脅迫下通過調(diào)節(jié)ABA和GA的含量促進(jìn)種子萌發(fā)[24-25],提高幼苗的質(zhì)量[26],但在正常生長條件下反而會(huì)通過抑制GA誘導(dǎo)的α淀粉酶編碼基因的表達(dá)抑制種子萌發(fā)[27],由此可見,不同化學(xué)藥劑處理促進(jìn)大葉樟種子休眠解除的機(jī)理并不一致。
綜上所述,種子從休眠向萌發(fā)階段的轉(zhuǎn)換過程十分復(fù)雜,涉及信號(hào)轉(zhuǎn)導(dǎo)、酶的活化、養(yǎng)分轉(zhuǎn)化、基因表達(dá)等多個(gè)方面。化學(xué)藥劑處理參與并促進(jìn)種子休眠向萌芽階段的轉(zhuǎn)化,適宜濃度的GA3、MT、H2O2和SA處理均能促進(jìn)大葉樟種子休眠解除,提高種苗質(zhì)量,但不同化學(xué)藥劑對(duì)種子休眠解除的效果不一致。結(jié)合隸屬函數(shù)法的結(jié)果來看,200 μg·L-1的MT和300 mg·L-1的GA3處理效果較好,D值分別為0.805,0.761,而500 μg·L-1的MT和2 mmol·L-1的SA處理效果較差,D值分別為0.113,0.025,化學(xué)藥劑促進(jìn)種子萌發(fā)可能還涉及其他代謝途徑,不同化學(xué)藥劑調(diào)控大葉樟種子休眠解除的網(wǎng)絡(luò)系統(tǒng)尚需進(jìn)一步研究。
參考文獻(xiàn):
[1] 肖祖飛, 張北紅, 李鳳, 等. 樟樹種子發(fā)芽及幼苗生長研究[J]. 安徽農(nóng)學(xué)通報(bào), 2020, 26(7): 40-45, 63.
[2] 周佑勛, 段小平, 肖東玉. 樟樹、檫木、閩楠種子的休眠和萌發(fā)特性[J]. 中南林學(xué)院學(xué)報(bào), 2006, 26(5): 79-84.
[3] 謝金蘭, 范劍明, 張冬生, 等. 去皮解除休眠對(duì)楠木種子萌芽及幼苗生長影響[J]. 園藝與種苗, 2017(8): 18-19, 45.
[4] 王付娟, 劉書含, 李淑梅, 等. 野生大豆種皮障礙休眠解除及萌發(fā)的研究[J]. 大豆科學(xué), 2019, 38(5): 733-739.
[5] 嚴(yán)加坤, 周奇, 劉莉. 褪黑素處理對(duì)干旱脅迫下玉米種子萌發(fā)的影響[J]. 山西農(nóng)業(yè)科學(xué), 2019, 47(2): 159-161, 193.
[6] 于麗麗, 曹瑞珍, 張學(xué)富, 等. 外源水楊酸對(duì)鹽脅迫下水飛薊種子萌發(fā)及幼苗生理特性的影響[J]. 分子植物育種, 2019, 17(23): 7909-7917.
[7] 張靜靜, 吳軍, 程許娜, 等. 樟樹種子休眠機(jī)制初探[J]. 河南農(nóng)業(yè)科學(xué), 2011, 40(10): 123-125, 136.
[8] 梁機(jī), 王志勇, 覃鵠, 等. 不同貯藏和浸種方式對(duì)香樟種子發(fā)芽率的影響[J]. 安徽農(nóng)業(yè)科學(xué), 2010, 38(6): 3231-3233.
[9] 羅立娜, 李紹鵬, 吳凡, 等. 隸屬函數(shù)法篩選多效唑?qū)ο憬冻看匍L效果的最佳劑量[J]. 分子植物育種, 2018, 16(12): 4079-4085.
[10] 楊立文, 劉雙榮, 林榮呈. 光信號(hào)與激素調(diào)控種子休眠和萌發(fā)研究進(jìn)展[J]. 植物學(xué)報(bào), 2019, 54(5): 569-581.
[11] 劉晏, 李俊德, 李家儒. 脫落酸和赤霉素調(diào)控種子休眠與萌發(fā)研究進(jìn)展[J]. 生物資源, 2020, 42(2): 157-163.
[12] GRAEBER K, NAKABAYASHI K, MIATTON E, et al. Molecular mechanisms of seed dormancy[J]. Plant Cell and Environment, 2012, 35(10): 1769-1786.
[13] 陸秀君, 劉月洋, 李天來, 等. 變溫層積過程中GA3誘導(dǎo)天女木蘭種子貯藏物質(zhì)及糖代謝關(guān)鍵酶活性的變化[J]. 沈陽農(nóng)業(yè)大學(xué)學(xué)報(bào), 2010, 41(1): 18-22.
[14] SONG W, HAO Q, CAI M, et al. Rice OsBT1 regulates seed dormancy through the glycometabolism pathway[J]. Plant Physiology and Biochemistry: PPB, 2020, 151: 469-476.
[15] SONG Y, ZHU J. The roles of metabolic pathways in maintaining primary dormancy of Pinus koraiensis seeds[J]. BMC Plant Biology, 2019, 19(1): 550.
[16] S?魣NCHEZ-MONTESINO R, BOUZA-MORCILLO L, MARQUEZ J, et al. A regulatory module controlling GA-mediated endosperm cell expansion is critical for seed germination in Arabidopsis[J]. Molecular Plant, 2019, 12(1): 71-85.
[17] SHU K, LIU X D, XIE Q, et al. Two faces of one seed: hormonal regulation of dormancy and germination[J]. Molecular Plant, 2016, 9(1): 34-45.
[18] N?魪E G, XIANG Y, SOPPE W J. The release of dormancy, a wake-up call for seeds to germinate[J]. Current Opinion in Plant Biology, 2017, 35: 8-14.
[19] LIU Y, YE N, LIU R, et al. H2O2 mediates the regulation of ABA catabolism and GA biosynthesis in Arabidopsis seed dormancy and germination[J]. Journal of Experimental Botany, 2010, 61(11): 2979-2990.
[20] ZHANG H J, ZHANG N, YANG R C, et al. Melatonin promotes seed germination under high salinity by regulating antioxidant systems, ABA and GA4 interaction in cucumber (Cucumis sativus L.)[J]. Journal of Pineal Research, 2014, 57(3): 269-279.
[21] PARKHEY S, NAITHANI S C, KESHAVKANT S. ROS production and lipid catabolism in desiccating Shorea robusta seeds during aging[J]. Plant Physiology and Biochemistry, 2012, 57(8): 261-267.
[22] 張海軍. 褪黑素處理對(duì)氯化鈉脅迫下黃瓜種子萌發(fā)的影響及其分子機(jī)制[D]. 北京: 中國農(nóng)業(yè)大學(xué), 2016.
[23] 陳莉, 劉連濤, 馬彤彤, 等. 褪黑素對(duì)鹽脅迫下棉花種子抗氧化酶活性及萌發(fā)的影響[J]. 棉花學(xué)報(bào), 2019, 31(5): 438-447.
[24] LEE S, KIM S G, PARK C M. Salicylic acid promotes seed germination under high salinity by modulating antioxidant activity in Arabidopsis[J]. The New Phytologist, 2010, 188(2): 626-637.
[25] LI Z, XU J, GAO Y, et al. The synergistic priming effect of exogenous salicylic acid and H2O2 on chilling tolerance enhancement during maize(Zea mays L.) seed germination[J]. Frontiers in Plant Science, 2017, 8: 1153.
[26] 孫曉春, 張惠惠, 黃文靜, 等. 干旱脅迫下水楊酸對(duì)桔梗種子萌發(fā)及植物激素的影響[J]. 中國農(nóng)業(yè)科技導(dǎo)報(bào), 2019, 21(10): 74-79.
[27] XIE Z, ZHANG Z L, HANZLIK S, et al. Salicylic acid inhibits gibberellin-induced alpha-amylase expression and seed germination via a pathway involving an abscisic-acid-inducible WRKY gene[J]. Plant Molecular Biology, 2007, 64(3): 293-303.
