小麥干物質(zhì)積累與分配規(guī)律研究進(jìn)展
2021-07-16 08:53:44柯媛媛倪芊芊張樂(lè)樂(lè)魏鳳珍李金才
大麥與谷類(lèi)科學(xué) 2021年3期
柯媛媛,陳 翔,倪芊芊,張樂(lè)樂(lè),魏鳳珍,李金才,2*
(1.安徽農(nóng)業(yè)大學(xué)農(nóng)學(xué)院/農(nóng)業(yè)部華東地區(qū)作物栽培科學(xué)觀測(cè)站,安徽 合肥 230036;2.江蘇省現(xiàn)代作物生產(chǎn)協(xié)同創(chuàng)新中心,江蘇 南京 210095)
自1860年工業(yè)革命以來(lái),全球氣候顯著變暖[1]。據(jù)最新的全球表面溫度觀測(cè)數(shù)據(jù)集[2-3]分析得出,1900—2017年間全球陸地平均氣溫升高(1.00±0.06)℃/100年。氣候變化增加了糧食生產(chǎn)的不穩(wěn)定性,而2050年全球人口預(yù)計(jì)將達(dá)到96億,保障全球96億人口的糧食安全問(wèn)題將成為2050年面臨的最大挑戰(zhàn)。為滿(mǎn)足日益增長(zhǎng)的糧食需求,小麥生產(chǎn)力每年需增加1.6%[4]。解偉等通過(guò)構(gòu)建Meta回歸模型得出,全球平均氣溫每升高1℃,將導(dǎo)致我國(guó)主要糧食作物的單位面積產(chǎn)量降低2.6%[5]。小麥作為世界三大糧食作物之一,全球約60%的人口以小麥為主食[6]。在全球氣候復(fù)雜多變和人口激增的背景下,維護(hù)小麥生產(chǎn)的穩(wěn)定性對(duì)保障糧食安全具有極其重要的意義。
小麥干物質(zhì)的積累與分配過(guò)程反映了小麥群體生長(zhǎng)狀況的好壞,也是決定小麥籽粒產(chǎn)量的重要因素。了解小麥在大田生長(zhǎng)中干物質(zhì)積累與分配的一般規(guī)律,以及基因型、主要栽培因子和生態(tài)因子對(duì)小麥干物質(zhì)積累與分配的影響,對(duì)小麥生長(zhǎng)發(fā)育狀態(tài)的調(diào)控及建立高產(chǎn)群體結(jié)構(gòu)具有重要意義。本文系統(tǒng)綜述了小麥干物質(zhì)積累與分配的一般規(guī)律,并闡述基因型、栽培因子和生態(tài)因子對(duì)小麥干物質(zhì)積累與分配的影響,以期為改善小麥的種植制度,完善農(nóng)田生態(tài)系統(tǒng)以及提高我國(guó)小麥生產(chǎn)能力提供參考。
1 小麥干物質(zhì)積累與分配的一般規(guī)律
1.1 小麥干物質(zhì)的積累規(guī)律
作物產(chǎn)量的高低與干物質(zhì)積累過(guò)程密切相關(guān),分析作物生長(zhǎng)過(guò)程中干物質(zhì)的積累動(dòng)態(tài)對(duì)揭示作物產(chǎn)量形成和構(gòu)建高效群體具有重要實(shí)踐價(jià)值[7]。研究表明,隨著小麥生長(zhǎng)發(fā)育進(jìn)程的推進(jìn),葉片和莖鞘的干物質(zhì)積累量呈單峰曲線(xiàn),而穗和植株的干物質(zhì)積累量呈“S”型曲線(xiàn)變化[8]。小麥在不同生育時(shí)期,干物質(zhì)積累速率并不相同。陳宏等研究發(fā)現(xiàn),開(kāi)花期以前是小麥單莖干物質(zhì)積累量的緩增期,開(kāi)花期至灌漿期干物質(zhì)積累量急劇加快,灌漿期后減緩[9]。趙姣等通過(guò)Logistic曲線(xiàn)擬合越冬后小麥干物質(zhì)的積累過(guò)程,表明孕穗至開(kāi)花期小麥干物質(zhì)積累速率最大,在拔節(jié)中后期之前和花后20 d,干物質(zhì)積累緩慢增加[10]。此外,張黛靜等通過(guò)13C穩(wěn)定性同位素示蹤技術(shù)發(fā)現(xiàn),不同取樣時(shí)期小麥凈輸入光合13C總量范圍為21.71~94.06 mg/盆,隨著生育期的推進(jìn),葉和莖鞘的光合碳逐漸向穗部轉(zhuǎn)運(yùn),故葉和莖鞘部13C含量減少,穗部13C含量增加,表明小麥生長(zhǎng)發(fā)育各個(gè)階段,干物質(zhì)在不同器官的積累量也不同[11]。
1.2 小麥干物質(zhì)分配規(guī)律
干物質(zhì)在小麥各器官中的分配對(duì)于建立平衡的源庫(kù)關(guān)系及指導(dǎo)小麥高產(chǎn)優(yōu)質(zhì)生產(chǎn)具有重要意義。開(kāi)花期小麥干物質(zhì)在不同器官中分配比例一般表現(xiàn)為莖鞘>葉>穗,且莖鞘和穗的干物質(zhì)積累量與成熟期小麥植株干物質(zhì)量顯著相關(guān)[12]。郭增江等研究表明,灌漿前期籽粒中同化物占總生物量的35.25%~45.66%,穎殼+穗軸占7.40%~9.85%,莖稈占27.58%~39.75%;灌漿后期植株70%以上的同化物運(yùn)送至籽粒中[13]。Zhang等研究表明,成熟期小麥籽粒、莖葉、穗軸+穎殼和旗葉的13C分布所占比例分別為40.89%、31.09%、4.50%和2.19%[14]。此外,小麥各器官干物質(zhì)分配率達(dá)到峰值的時(shí)間不同。葉片的分配比例在拔節(jié)期達(dá)到最大,而后隨著生育進(jìn)程的推進(jìn)呈現(xiàn)出不斷降低的趨勢(shì),莖鞘所占比例呈先增后降的變化趨勢(shì)且在孕穗期達(dá)到最大;穗所占比例呈不斷增加趨勢(shì),并在成熟期達(dá)到最大值[15]。小麥干物質(zhì)在源庫(kù)之間的合理分配利用對(duì)發(fā)揮最大產(chǎn)量潛力至關(guān)重要,應(yīng)通過(guò)栽培措施的優(yōu)化來(lái)改善源庫(kù)關(guān)系,從而協(xié)調(diào)干物質(zhì)在源庫(kù)間的分配,實(shí)現(xiàn)小麥的穩(wěn)產(chǎn)、豐產(chǎn)與優(yōu)質(zhì)。
1.3 小麥干物質(zhì)積累、分配與產(chǎn)量的關(guān)系
小麥產(chǎn)量形成的過(guò)程,實(shí)質(zhì)上是同化物的生產(chǎn)制造及轉(zhuǎn)運(yùn)分配的過(guò)程。小麥籽粒產(chǎn)量主要來(lái)自于花前貯存的同化物向籽粒的轉(zhuǎn)運(yùn)及花后葉、莖鞘等營(yíng)養(yǎng)器官中光合同化物的積累[16]。花前是小麥營(yíng)養(yǎng)生長(zhǎng)的重要階段,此時(shí)干物質(zhì)的積累可為小麥產(chǎn)量的形成奠定基礎(chǔ)。研究發(fā)現(xiàn),拔節(jié)-開(kāi)花期小麥群體干物質(zhì)積累量與總結(jié)實(shí)粒數(shù)呈顯著二次曲線(xiàn)關(guān)系,說(shuō)明此時(shí)期干物質(zhì)積累量不宜過(guò)大,否則會(huì)導(dǎo)致無(wú)效分蘗增多,不利于建成平衡的群體生態(tài)結(jié)構(gòu),從而導(dǎo)致減產(chǎn)[17]。楊佳鳳等研究指出,孕穗-開(kāi)花期干物質(zhì)積累量與籽粒產(chǎn)量呈二次曲線(xiàn)關(guān)系,此時(shí)期適宜的干物質(zhì)積累量是形成高產(chǎn)的基礎(chǔ)[18]。花后是籽粒灌漿的關(guān)鍵時(shí)期,該階段較高的干物質(zhì)積累量能保證小麥正常的灌漿能力[17]。研究發(fā)現(xiàn),小麥花前干物質(zhì)轉(zhuǎn)運(yùn)量對(duì)產(chǎn)量的貢獻(xiàn)率為43.10%~45.22%,與產(chǎn)量呈線(xiàn)性負(fù)相關(guān);花后干物質(zhì)積累量對(duì)產(chǎn)量的貢獻(xiàn)率為54.78%~56.92%,與產(chǎn)量呈線(xiàn)性正相關(guān)[19]。因此,小麥群體后期干物質(zhì)積累量是實(shí)現(xiàn)高產(chǎn)的關(guān)鍵,在合理調(diào)控拔節(jié)-開(kāi)花期群體干物質(zhì)積累量的基礎(chǔ)上,應(yīng)重點(diǎn)擴(kuò)大花后干物質(zhì)積累量。
2 小麥干物質(zhì)積累與分配的影響因素
2.1 基因型對(duì)小麥干物質(zhì)積累與分配的影響
因發(fā)育特點(diǎn)不同,不同基因型小麥的“源”“庫(kù)”器官在調(diào)控干物質(zhì)轉(zhuǎn)運(yùn)分配的作用上存在一定差異(圖1)。研究表明,小麥基因型是影響干物質(zhì)轉(zhuǎn)運(yùn)和分配最主要的因素[20],不同品種小麥花后干物質(zhì)的積累量和花前干物質(zhì)的運(yùn)轉(zhuǎn)能力對(duì)籽粒的貢獻(xiàn)率存在差異[21]。吳金芝等研究發(fā)現(xiàn),晉麥47較偃展4110花前積累的干物質(zhì)多,花前干物質(zhì)運(yùn)轉(zhuǎn)能力強(qiáng),且花前貯藏干物質(zhì)對(duì)籽粒的貢獻(xiàn)率大,從而獲得較高產(chǎn)量[22]。仝錦等研究表明,超高產(chǎn)品種能顯著提高花后干物質(zhì)積累量及其對(duì)籽粒的貢獻(xiàn)率,最終產(chǎn)量較高[23]。
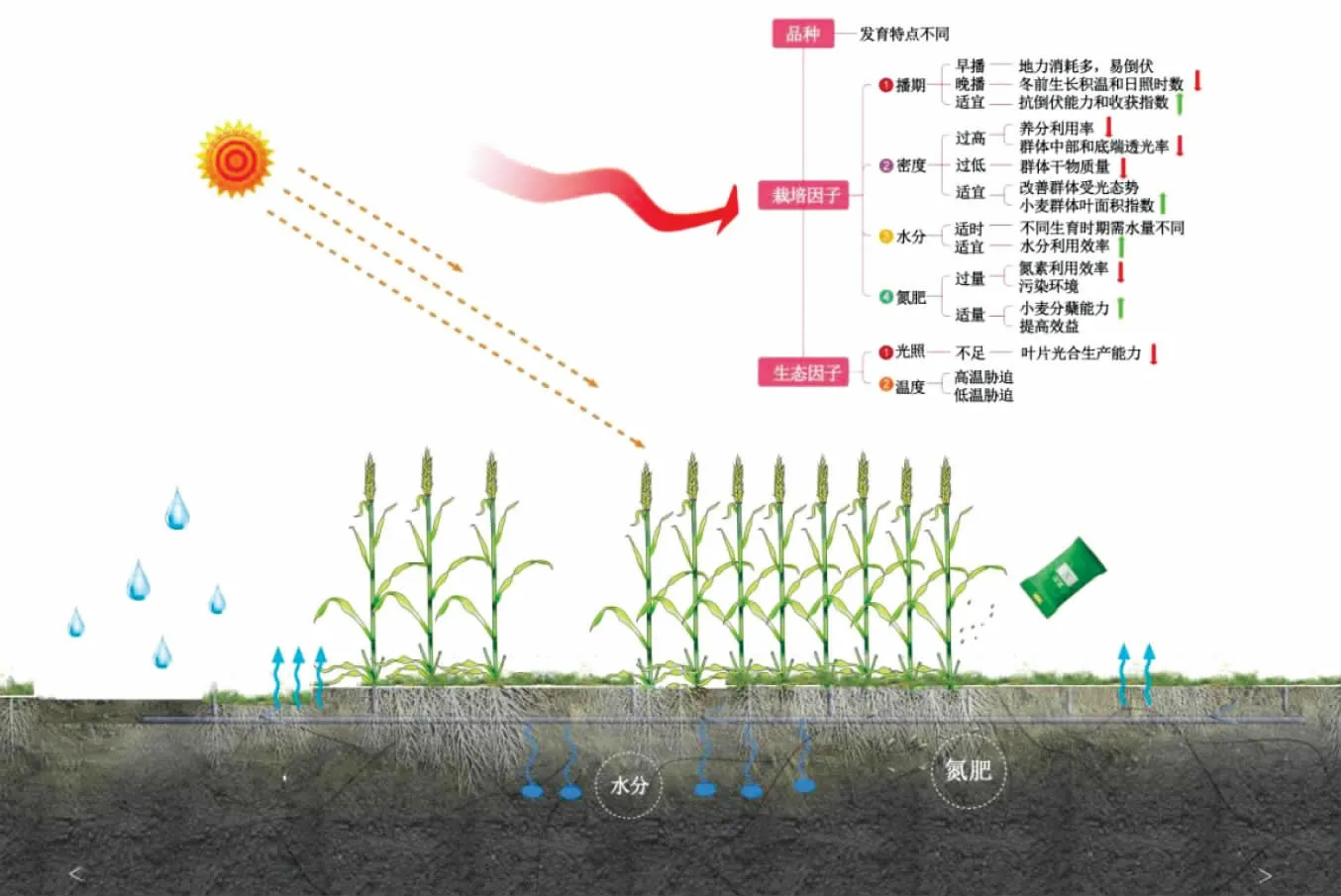
圖1 影響小麥干物質(zhì)積累與分配的因素
此外,不同基因型品種小麥的干物質(zhì)積累速率不同,從而影響成熟期小麥籽粒產(chǎn)量。張利等研究發(fā)現(xiàn),晚熟品種小麥較早熟品種的干物質(zhì)積累平均速率高,最大速率也明顯高于早熟品種,從而具有更強(qiáng)的生產(chǎn)潛勢(shì)[24]。李瑞珂等研究指出,高產(chǎn)小麥品種花前和花后干物質(zhì)積累速率較低產(chǎn)小麥分別高2.5倍和1.6倍,適宜的干物質(zhì)積累速率有利于小麥的高產(chǎn)優(yōu)質(zhì)[25]。因此,掌握不同基因型小麥干物質(zhì)積累及分配特性,通過(guò)改變其遺傳特性選取高產(chǎn)高效型小麥品種,可增加小麥“源”器官的生產(chǎn)能力和“庫(kù)”器官的容量,促進(jìn)強(qiáng)“源”暢“流”擴(kuò)“庫(kù)”,從而實(shí)現(xiàn)高產(chǎn)。
2.2 栽培因子對(duì)小麥干物質(zhì)積累與分配的影響
2.2.1 播期。播期主要通過(guò)影響小麥生育前期的積溫及光照等生態(tài)條件,使小麥生長(zhǎng)發(fā)育過(guò)程中干物質(zhì)的運(yùn)轉(zhuǎn)分配也相應(yīng)發(fā)生變化,從而影響小麥個(gè)體生長(zhǎng)和群體建成[26]。播期不同導(dǎo)致小麥冬前積溫不同,造成苗期單株和群體分蘗的干物質(zhì)積累量存在較大差異,進(jìn)而影響小麥后續(xù)干物質(zhì)的積累和產(chǎn)量形成。研究表明,因氣溫連續(xù)偏高,早播使小麥冬前出現(xiàn)旺長(zhǎng),甚至提前拔節(jié),導(dǎo)致小麥遇春季低溫易遭受凍害,進(jìn)而使群體干物質(zhì)積累量下降[27]。播期推遲,使小麥冬前生長(zhǎng)所需的有效積溫大幅度降低,導(dǎo)致小麥冬前生長(zhǎng)量不足,雖返青后干物質(zhì)積累速率加快,但無(wú)法彌補(bǔ)葉面積減少所帶來(lái)的干物質(zhì)積累量的損失,因此難以形成高產(chǎn)群體基礎(chǔ)[28-29]。適期晚播可提高小麥拔節(jié)—成熟期干物質(zhì)生產(chǎn)量,從而增加成熟期干物質(zhì)積累量,并結(jié)合較高的抗倒伏能力和收獲指數(shù)獲得較高產(chǎn)量[30]。氣候、土壤及農(nóng)民耕種習(xí)慣等因素均會(huì)影響播期,從而影響小麥最終經(jīng)濟(jì)產(chǎn)量。因此在實(shí)際生產(chǎn)中,應(yīng)通過(guò)多年播期試驗(yàn),結(jié)合作物品種、氣候、土壤條件、栽培措施等因素,同時(shí)考慮播期與密度、水分及肥料等因素的互作效應(yīng),探究播期對(duì)小麥干物質(zhì)積累與分配的調(diào)控效應(yīng),以實(shí)現(xiàn)最優(yōu)的栽培因子組合,為小麥的高產(chǎn)優(yōu)質(zhì)提供理論依據(jù)。
2.2.2 密度。密度作為高產(chǎn)栽培技術(shù)措施中重要的一環(huán),主要通過(guò)影響小麥群體結(jié)構(gòu)和光熱資源的分配來(lái)影響小麥產(chǎn)量形成[31]。密度過(guò)高會(huì)降低小麥對(duì)養(yǎng)分的利用率,加劇群體內(nèi)部競(jìng)爭(zhēng),不利于產(chǎn)量形成[32];群體密度過(guò)低,小麥干物質(zhì)在各生育時(shí)期積累量均較低,同樣不利于群體產(chǎn)量的提高[33]。因此,明確密度與干物質(zhì)積累與分配之間的關(guān)系,對(duì)于構(gòu)建合理的群體結(jié)構(gòu)從而實(shí)現(xiàn)小麥高產(chǎn)具有重要意義。劉娟等基于歸一化法,認(rèn)為小麥干物質(zhì)積累中期(快增期)是干物質(zhì)積累速率對(duì)密度的敏感反應(yīng)期[34]。而播量過(guò)大會(huì)降低小麥葉片的光能利用率,過(guò)度消耗土壤水分,導(dǎo)致葉片的光合能力下降,干物質(zhì)積累減少。丁位華等研究發(fā)現(xiàn),播量過(guò)大會(huì)使小麥整個(gè)生育期群體光合速率較高,但中部和底端透光率較小,光能利用率較低,從而導(dǎo)致小麥群體干物質(zhì)積累量和產(chǎn)量降低[35]。Reddy等認(rèn)為,過(guò)高的種植密度會(huì)消耗過(guò)多的土壤水分,使小麥開(kāi)花期土壤含水量較低,導(dǎo)致氣孔導(dǎo)度和光合能力下降,從而影響干物質(zhì)的積累[36-37]。適宜播量會(huì)改變小麥群體結(jié)構(gòu),改善群體受光態(tài)勢(shì),促進(jìn)群體對(duì)光能充分利用,增加干物質(zhì)的積累,從而提高產(chǎn)量[38]。因此,適宜的密度會(huì)提高小麥群體數(shù)量和葉面積指數(shù),延長(zhǎng)葉片光合功能期,優(yōu)化群體光合性能,從而促進(jìn)小麥干物質(zhì)的積累。
2.2.3 水分。不同生育時(shí)期小麥對(duì)水分的需求不同,土壤水分過(guò)多或不足及灌水時(shí)期都會(huì)對(duì)小麥干物質(zhì)的積累與分配造成影響。Zhang等研究表明,灌溉至季節(jié)性水分蒸發(fā)量的94%時(shí),小麥成熟期干物質(zhì)積累量最高[39]。姚寧等研究發(fā)現(xiàn),在越冬期和返青期對(duì)小麥進(jìn)行干旱處理,拔節(jié)后生物量顯著降低且后期復(fù)水無(wú)法彌補(bǔ)[40]。韓惠芳等研究指出,拔節(jié)期灌溉有利于小麥貯藏物質(zhì)的積累,同時(shí)拔節(jié)水和抽穗水更有利于提高貯藏物質(zhì)的轉(zhuǎn)運(yùn)效率[41]。此外,水分脅迫對(duì)小麥不同營(yíng)養(yǎng)器官干物質(zhì)累積的影響不同,其中對(duì)葉片的影響最大,對(duì)穗的影響最小[42],復(fù)水后小麥植株各器官的干物質(zhì)累積和產(chǎn)量均有不同程度的恢復(fù)[43]。
水分是小麥干物質(zhì)積累與分配的重要限制因子,不同的灌水處理造成土壤水分的差異,而小麥營(yíng)養(yǎng)器官貯存的同化物受土壤水分含量的調(diào)控[44]。適時(shí)適量灌溉可緩解土壤水分缺乏,提高水分利用效率,促進(jìn)小麥干物質(zhì)積累[45]。谷艷芳等研究發(fā)現(xiàn),干旱脅迫下,冬小麥植株和器官水平上干物質(zhì)積累量均顯著下降,干物質(zhì)分配趨向生長(zhǎng)中心,花后同化物的積累量減少50%,但營(yíng)養(yǎng)器官干物質(zhì)的轉(zhuǎn)運(yùn)量會(huì)增加30%以上[46-47]。適宜的水分有利于增加小麥地上部植株和不同器官(葉片、莖鞘、穎殼)的干物質(zhì)量,促進(jìn)籽粒產(chǎn)量形成[48]。因此在小麥生產(chǎn)中,可在小麥關(guān)鍵生育時(shí)期通過(guò)合理供水來(lái)提高水分利用效率,從而增加干物質(zhì)積累量和調(diào)控物質(zhì)轉(zhuǎn)運(yùn),最終提高產(chǎn)量。
2.2.4 氮肥。氮肥是調(diào)節(jié)小麥干物質(zhì)轉(zhuǎn)運(yùn)能力從而實(shí)現(xiàn)小麥高產(chǎn)栽培的重要養(yǎng)分限制因子。合理增施氮肥對(duì)小麥器官建成和營(yíng)養(yǎng)物質(zhì)分配有正向調(diào)控作用,能夠協(xié)同提高干物質(zhì)積累量和干物質(zhì)轉(zhuǎn)運(yùn)能力,從而實(shí)現(xiàn)高產(chǎn)[49]。研究表明,在一定范圍內(nèi)適量增施氮肥可以提高小麥花后干物質(zhì)積累量及其向籽粒中的轉(zhuǎn)運(yùn)效率[50]。蔡瑞國(guó)等研究指出,適量施氮提高了小麥干物質(zhì)積累量,同時(shí)促進(jìn)花前貯存干物質(zhì)向穗部的轉(zhuǎn)運(yùn),有利于產(chǎn)量三因素的協(xié)調(diào)發(fā)展[51]。李娜等研究表明,適宜的施氮量有助于增強(qiáng)植株生長(zhǎng)后期光合作用能力和提高分蘗能力,協(xié)調(diào)小麥花前和花后干物質(zhì)積累,促進(jìn)營(yíng)養(yǎng)物質(zhì)向籽粒快速轉(zhuǎn)移,從而提高花后光合產(chǎn)物對(duì)籽粒的貢獻(xiàn)率,對(duì)小麥增產(chǎn)具有重要意義[52]。
然而,在小麥生產(chǎn)中為追求高產(chǎn)過(guò)量施用化肥的現(xiàn)象屢見(jiàn)不鮮,這不僅極易造成氮素?fù)p失,導(dǎo)致小麥氮素利用效率以及氮農(nóng)學(xué)利用效率降低,不利于資源可持續(xù)發(fā)展,同時(shí)也給農(nóng)田環(huán)境帶來(lái)巨大壓力[53-54]。研究表明,目前農(nóng)業(yè)系統(tǒng)中的氮肥盈余量已經(jīng)達(dá)到175 kg/hm2[55],過(guò)量施氮使群體結(jié)構(gòu)變差,導(dǎo)致干物質(zhì)積累量減少,小麥花后物質(zhì)生產(chǎn)量增幅較小甚至下降[52]。因此,根據(jù)地區(qū)和品種的不同來(lái)判斷最適宜的氮施用量,不僅可以有效控制成本、提高效益,還可以合理分配和充分利用氮肥資源,減少農(nóng)業(yè)生產(chǎn)過(guò)程中養(yǎng)分損失和對(duì)環(huán)境造成的負(fù)面影響,使小麥同化物合成、貯存和轉(zhuǎn)運(yùn)三者相協(xié)調(diào),挖掘小麥產(chǎn)量潛力。
2.3 生態(tài)因子對(duì)小麥干物質(zhì)積累與分配的影響
2.3.1 光照。光照是影響小麥干物質(zhì)積累和產(chǎn)量的重要環(huán)境因子,小麥的生產(chǎn)潛力很大程度取決于接受太陽(yáng)光照的多寡以及光能利用率的高低。在黃淮麥區(qū),霧霾、陰雨寡照等因素引起的弱光問(wèn)題已成為限制小麥高產(chǎn)的重要因素[56],弱光造成小麥接受有效光合輻射減少,使光化學(xué)反應(yīng)速率和氣孔開(kāi)度下降,從而影響小麥干物質(zhì)的積累。據(jù)報(bào)道,黃淮海地區(qū)的光照度以每年0.6%的速度下降,導(dǎo)致小麥葉片光合速率降低,嚴(yán)重影響小麥生產(chǎn)[57]。顧蘊(yùn)倩等通過(guò)構(gòu)建SUCROS模型發(fā)現(xiàn),日總光合有效輻射低于3.71 MJ/m2且持續(xù)4 d以上對(duì)小麥干物質(zhì)的積累和產(chǎn)量產(chǎn)生顯著影響[58]。此外,弱光會(huì)降低小麥“源”器官葉片干物質(zhì)的生產(chǎn)能力,同時(shí)阻礙干物質(zhì)向“庫(kù)”器官穗部的轉(zhuǎn)運(yùn),最終導(dǎo)致產(chǎn)量降低。Xu等研究發(fā)現(xiàn),弱光會(huì)影響小麥干物質(zhì)積累和同化物向籽粒的運(yùn)輸,導(dǎo)致產(chǎn)量下降[59]。劉希偉等研究指出,花后不同階段遮光處理均會(huì)導(dǎo)致小麥光合速率下降,干物質(zhì)生產(chǎn)不足,籽粒灌漿速率降低,進(jìn)而影響產(chǎn)量[60]。李劉龍等研究發(fā)現(xiàn),弱光條件下籽粒的“庫(kù)吸力”降低,使植株貯藏的同化物更多地滯留在營(yíng)養(yǎng)器官中,用于維持“源”器官的功能代謝,導(dǎo)致干物質(zhì)在小麥“庫(kù)”器官穗部干物質(zhì)的積累量減少[61]。因此,光照不足會(huì)使小麥功能葉片光合產(chǎn)物供應(yīng)不足,直接影響到干物質(zhì)的積累與分配,從而導(dǎo)致減產(chǎn)。
2.3.2 溫度。溫度是小麥進(jìn)行營(yíng)養(yǎng)生長(zhǎng)和生殖生長(zhǎng)的重要生態(tài)因子,小麥完成其生命周期需要一定的積溫[62]。馬倩倩等通過(guò)皮爾遜相關(guān)性分析等方法研究發(fā)現(xiàn),小麥全年≥10℃的積溫為3 500℃·d左右,播種-成熟期>0℃積溫約為2 200℃·d[63]。小麥的生長(zhǎng)發(fā)育在不同的生育時(shí)期所要求的溫度也不同。小麥播種的適宜溫度為15~16℃,而出苗后溫度需要降到5~6℃,拔節(jié)到抽穗期則要求溫度逐漸升高,最后達(dá)到21~22℃[64]。然而,全球氣候變暖的加劇導(dǎo)致極端溫度事件頻發(fā),嚴(yán)重威脅到小麥生產(chǎn)[65]。高溫脅迫會(huì)加速葉片衰老,縮短葉片中光合產(chǎn)物向籽粒中的運(yùn)輸時(shí)間,導(dǎo)致籽粒灌漿終止早于葉片完全衰老,葉片光合產(chǎn)物被分配到其他營(yíng)養(yǎng)器官中[66],抑制了籽粒中干物質(zhì)的積累,最終影響粒質(zhì)量和產(chǎn)量。張姍等研究發(fā)現(xiàn),高溫脅迫導(dǎo)致的花后干物質(zhì)積累量和花前干物質(zhì)運(yùn)轉(zhuǎn)率共同降低可能是籽粒產(chǎn)量降低的主要原因[67]。
而低溫則會(huì)導(dǎo)致小麥葉片的光合速率、蒸騰速率和葉片氣孔導(dǎo)度顯著下降,胞間CO2濃度大幅升高,導(dǎo)致干物質(zhì)積累減少,從而造成減產(chǎn)[68]。張溪荷等研究發(fā)現(xiàn),低溫脅迫導(dǎo)致莖稈干物質(zhì)向穗部的轉(zhuǎn)運(yùn)受阻,營(yíng)養(yǎng)物質(zhì)累積在莖稈中,所以莖稈分配比例升高,穗分配比例降低[69]。因此,溫度脅迫主要通過(guò)減少小麥籽粒灌漿時(shí)間和降低葉片光合速率等一系列生理生化過(guò)程來(lái)降低干物質(zhì)的積累量和分配,最終導(dǎo)致小麥減產(chǎn)。
3 展望
3.1 運(yùn)用分子生物學(xué)技術(shù)選育小麥新品種
小麥的產(chǎn)量和品質(zhì)是受多基因和環(huán)境互作影響的復(fù)雜數(shù)量性狀,單純依靠現(xiàn)有常規(guī)育種技術(shù)耗時(shí)長(zhǎng)、成本高[70],難以滿(mǎn)足全球日益增長(zhǎng)的糧食需求,小麥基因組和分子遺傳育種的發(fā)展為進(jìn)一步解決世界糧食問(wèn)題提供了新的契機(jī)和突破口。因此,應(yīng)利用CRISPR/Cas9基因編輯技術(shù)[71]和分子標(biāo)記輔助育種,借助轉(zhuǎn)錄組、蛋白質(zhì)組、代謝組等多組學(xué)技術(shù)加快對(duì)小麥功能基因的定位,從而加快推動(dòng)種質(zhì)資源的利用和新品種的選育。
3.2 利用作物表型組學(xué)、作物模型等探索小麥干物質(zhì)積累過(guò)程機(jī)理
定量分析小麥生長(zhǎng)過(guò)程中各項(xiàng)指標(biāo)的動(dòng)態(tài)變化是揭示產(chǎn)量形成和掌握群體調(diào)控指標(biāo)的重要內(nèi)容。高通量表型組學(xué)通過(guò)從微觀到宏觀、從超微到器官再到個(gè)體乃至群體的多維度數(shù)據(jù)以實(shí)現(xiàn)對(duì)作物不同層次的監(jiān)控,從而獲取作物不同生長(zhǎng)發(fā)育階段的動(dòng)態(tài)性狀,對(duì)作物群體的早期診斷和定量調(diào)控具有重要意義。今后,應(yīng)充分利用各種作物表型數(shù)據(jù)庫(kù),并結(jié)合Richards、Logistic、Compertz等生長(zhǎng)模型探索干物質(zhì)積累過(guò)程機(jī)理[71-73],深入研究各影響因子與干物質(zhì)積累的定量關(guān)系,為提升小麥干物質(zhì)積累的途徑提供新思路。
3.3 集成多項(xiàng)生產(chǎn)技術(shù)實(shí)現(xiàn)農(nóng)業(yè)可持續(xù)發(fā)展
隨著農(nóng)業(yè)資源供需矛盾的加劇,小麥生產(chǎn)的可持續(xù)性成為研究的焦點(diǎn)。未來(lái)應(yīng)集成多項(xiàng)節(jié)水、綠色、高效生產(chǎn)技術(shù)模式,通過(guò)推廣自動(dòng)駕駛、衛(wèi)星平地、精量播種、精準(zhǔn)施肥、智能灌溉、無(wú)人植保、智慧收獲等多項(xiàng)智能化技術(shù),推動(dòng)小麥生產(chǎn)向節(jié)水、穩(wěn)產(chǎn)、綠色、優(yōu)質(zhì)、高效種植發(fā)展,并且結(jié)合地區(qū)的氣候特點(diǎn)、災(zāi)害發(fā)生規(guī)律以及生產(chǎn)條件等,在提高系統(tǒng)生產(chǎn)效率和可持續(xù)性的基礎(chǔ)上實(shí)現(xiàn)精細(xì)化農(nóng)業(yè)管理,從而實(shí)現(xiàn)小麥生產(chǎn)的可持續(xù)發(fā)展。
猜你喜歡
艦船科學(xué)技術(shù)(2022年13期)2022-08-11 09:30:02
礦山安全信息(2022年40期)2022-04-07 02:16:52
今日農(nóng)業(yè)(2021年14期)2021-11-25 23:57:29
石油與天然氣地質(zhì)(2021年1期)2021-02-22 14:14:44
今日農(nóng)業(yè)(2020年20期)2020-11-26 06:09:10
鐵道通信信號(hào)(2020年9期)2020-02-06 09:15:22
中國(guó)果業(yè)信息(2019年10期)2019-11-13 01:21:34
數(shù)學(xué)大王·趣味邏輯(2019年5期)2019-06-13 20:27:43
小學(xué)科學(xué)(學(xué)生版)(2019年5期)2019-05-21 01:00:18
經(jīng)濟(jì)技術(shù)協(xié)作信息(2018年30期)2018-11-22 06:20:24
