動物鼠疫周期流行的生態位影響因素研究進展
2021-07-16 07:57:28秦婧靚楊瑞馥崔玉軍
中國人獸共患病學報 2021年6期
關鍵詞:生態
秦婧靚,楊瑞馥,崔玉軍
傳染病是病原微生物或寄生蟲感染人體后產生的具有傳染性的疾病,可在人群中傳播與流行[1]。地理、氣候、社會因素等對傳染病的流行及靜息均可產生影響。若該傳染病在當地一直處于停息狀態但沒有被根除,認為其處于靜息期(latent enzootic phase);若該病發病率顯著高于歷年一般水平時,則認為該病處于流行期(epizootic phase)[1]。鼠疫(Plague)是我國法定的甲類傳染病,其發病急、傳染性強,治療不及時病死率高,不依賴于人類而獨立存在于自然疫源地中[2]。在鼠疫自然疫源地中,可觀測到鼠疫存在周期性流行,即流行期與靜息期發生交替的現象。但是交替周期并不固定,其中靜息期的時間跨度可長達數十年或更久。如:我國廣西鼠疫靜息53年后,于2000年7月在隆林縣天生橋庫區及周邊發生動物間疫情,并波及人間[3-4]。云南省動物間鼠疫于1982年在滇西復燃,隨后滇南、滇中、滇東也出現疫情。其中普洱、思茅、石屏、宜良縣鼠疫疫源地分別靜息72年、93年、100年、134年后又開始活躍并出現人間疫情[5]。
生態位(niche)是有機體種群對所生存環境條件的耐受性和生境資源需求的綜合。鼠疫生態位包括疫源地的環境、宿主、媒介等因素[6]。值得注意的是:鼠疫耶爾森菌(Yersiniapestis,簡稱鼠疫菌)本身也是鼠疫生態位中重要一環。鼠疫菌會通過對媒介/宿主的影響,改變生態位的整體平衡。在適宜生態條件下,鼠疫菌、宿主動物、媒介生物的種群數量達到動態平衡,鼠疫菌可進行穩定的定植和傳播,導致鼠疫在動物間流行。而當生態位的動態平衡被破壞時,動物間鼠疫就可能靜息或者隨宿主、媒介遷徙溢出到其他地區[7]。現有研究表明動物間鼠疫周期性流行特征的出現與鼠疫自然疫源地生態位的動態變化密切相關[8-9]。例如中世紀鼠疫大流行時,歐洲大部分地區存在鼠疫疫情,但嚙齒類動物種群改變[10];居民生活水平提高[10-11];動物棲息地喪失和工業化發展[12]等因素破壞原生態位的動態平衡,則使歐洲鼠疫菌滅絕,疫源地消失[13]。本文將從影響鼠疫自然疫源地生態位穩定的主要因素入手,對宿主動物、蚤類媒介、土壤、鼠疫菌多樣性及人類活動5個方面的相關研究進行梳理總結(圖1、表1)。

表1 影響動物間周期性鼠疫流行的主要因素Tab.1 The key factors affecting periodic plague epizootics
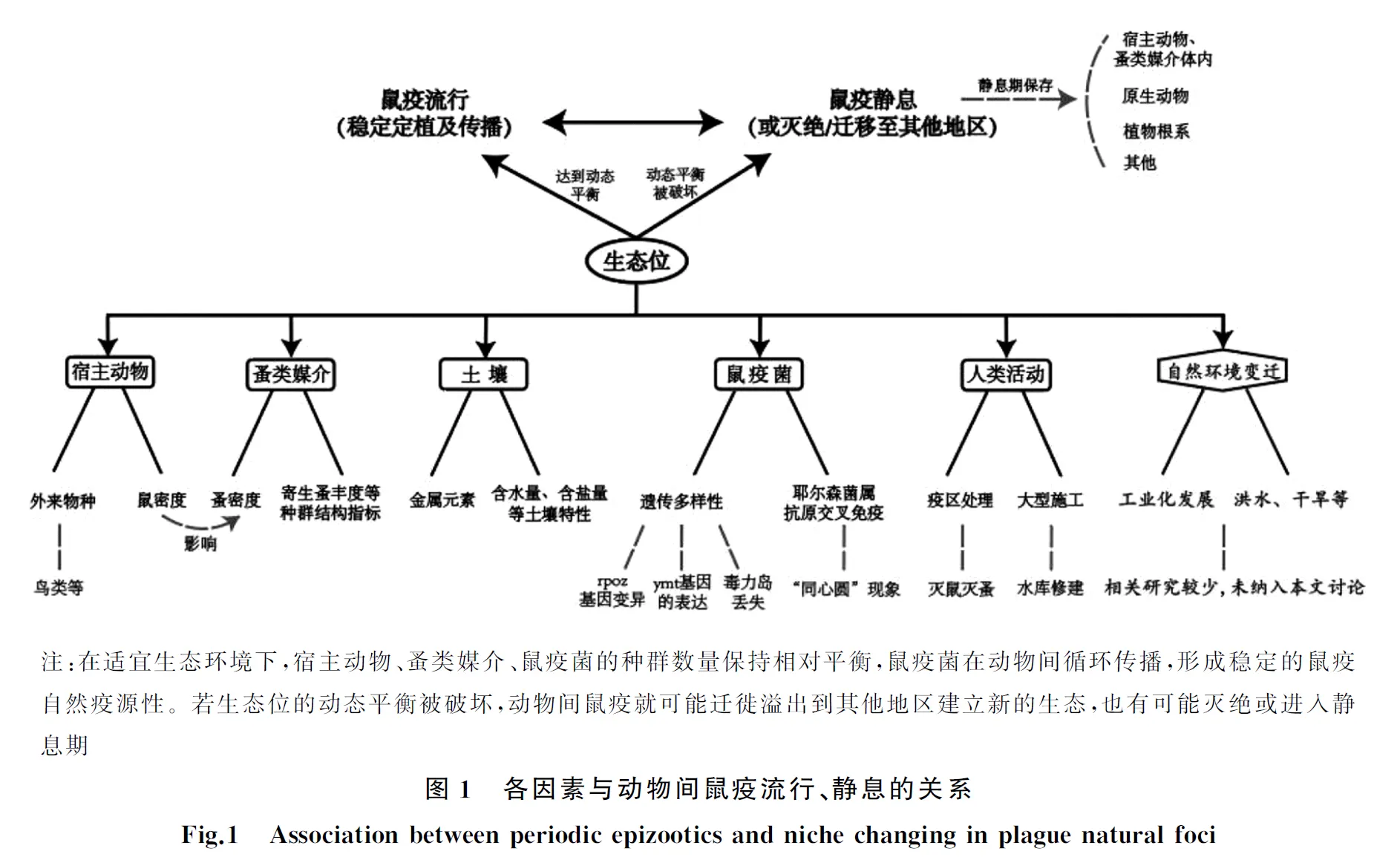
1 宿主動物與生態位
目前,世界上共發現2 369種嚙齒類動物[14], 截至2017年8月底,我國記錄在冊嚙齒目達到9科78屬220種[15],染疫嚙齒類動物53種[16],傳播鼠疫菌的主要宿主動物(鼠類)有14種[17],分布于我國北部、西北部、西南部及東南沿海地區[18]。嚙齒類動物作為鼠疫菌的主要宿主,對維持鼠疫自然疫源地生態位的動態平衡具有重要作用。我國鼠疫歷年監測數據顯示,動物間鼠疫流行及靜息與該疫源地主要宿主動物的密度有直接的關聯。研究人員通過對甘寧阿拉善黃鼠疫源地50年間鼠疫疫情進行分析,發現該疫源地發生的3次動物間鼠疫流行期的年均鼠密度顯著高于靜息期平均值,流行期當地黃鼠年均密度均高于1只/hm2[19]。高少坤等[20]發現河北省鼠疫自然疫源地1993-1995年、2001-2005年宿主動物密度明顯高于1981-2013年的平均水平,使鼠疫在動物間流行可能性增大,影響了鼠疫疫情的發生和發展。也有學者認為外來物種介入會對疫源地生態位的相對平衡產生影響,如:被鼠疫感染的鳥類或其他物種進入處于靜息期或鼠疫非流行地區時,會引起該疫源地鼠疫再次流行[21]。
2 媒介生物與生態位
據統計,全世界已發現蚤類2 500余種和亞種,分別隸屬于5總科16科238屬。我國已發現4總科10科75屬655種和亞種[22],傳播鼠疫菌的主要媒介昆蟲(蚤類)有28種[23]。蚤類作為鼠疫菌傳播的主要媒介及儲存宿主,主要通過在不同宿主間吸血傳播鼠疫菌。蚤類密度增高不僅會增加對宿主的叮咬次數,而且提高了鼠疫菌在宿主間的傳播概率,對疫源地內動物間鼠疫流行和鼠疫菌延續起重要作用[24]。在漫長的生物進化中,蚤類與宿主形成寄生或共生關系,蚤類數量的變化往往又受宿主數量的影響。前期研究表明:體蚤指數與鼠密度呈正相關關系,鼠密度增加則體蚤指數相應增高[25]。若生態環境改變,疫源地的主要宿主動物隨之變化,也間接影響了相應的主要寄生蚤數量及分布。此外,鼠疫生態系統的穩定還與蚤類群落其他結構指標密切相關。葉超等[26]研究發現,準噶爾盆地大沙鼠疫源地內寄生蚤對大沙鼠的侵染能力(染蚤率)和寄生數量(總蚤指數)的季節性變化與該地鼠疫季節性的流行特征一致。與此同時,蚤類群落的多樣性和豐富度增高、優勢度降低。這些結構指標的變化有利于維持疫源地生態位的動態平衡。
3 土壤與生態位
土壤是微生物的聚集地,它與宿主、媒介構成的小型生態系統共同維持鼠疫菌生命的延續[27]。鼠疫菌能夠在土壤中生存,其與含鹽量、含水量、金屬元素等土壤特性息息相關。相對濕潤、中等鹽堿的土壤不僅為微生物和地表動物的生存和繁殖提供了有利條件,而且益于鼠疫菌在疫源地的長期保存[28]。此外,土壤中的金屬元素(例如鐵、鈣)可參與調節自然界中動物感染鼠疫的過程[29],促進細菌生長,增強其毒力[30],提高鼠疫菌感染宿主的能力[31]。但金屬元素過量或缺乏都會影響細菌的生存和繁殖,鼠疫菌對各類金屬元素的攝取應受到精確調控[32]。當疫源地內土壤條件適宜、微生物種類豐富、金屬離子濃度相對平衡時,有利于宿主或鼠疫菌生存,可能促進鼠疫發生或流行。而暴雨洪澇等極端氣候可使土壤鹽度、含水量、金屬離子濃度發生變化,有可能導致鼠疫疫源地局部滅絕或進入靜息期[33]。
4 “外延的表型”:鼠疫菌遺傳變異與生態位
“外延的表型”(extended phenotype)是理查德道金斯在1982年提出的概念,強調基因作為被選擇的基本單元,可在生物體外產生表型效應(通過基因組變異造成個體周圍環境改變)[34],并且能夠延伸至更廣泛的環境或其他個體中。例如鳥巢就是鳥類基因組通過某種形式所編碼的具體實物。我們的研究表明,細菌中也存在類似現象[35]。因此,鼠疫菌遺傳多樣性能夠通過“外延的表型”機制,與生態位發生作用。
4.1 鼠疫菌基因組多樣性對生態位的影響和關聯 鼠疫菌可以在蚤類前胃中形成生物膜,從而阻礙血液進入蚤類消化道,使其一直處于饑餓狀態,吸血行為更為頻繁,最終促進鼠疫菌的快速播散[36-37],其可通過遺傳變異調節與蚤體內生物膜形成相關的表型,從而改變鼠疫菌經蚤傳播能力,以適應生態環境選擇壓力[35]。rpoZ基因主要編碼RNA聚合酶ω亞基,與鼠疫菌的生長速率和生物膜的形成能力有關[38]。在新疆天山古爾圖鼠疫自然疫源地的研究發現:該地區秋冬季發生異常寒冷、干旱時,鼠疫菌rpoZ基因上可觀測到很強的正向選擇信號。體外實驗表明該基因突變會導致鼠疫菌生物膜形成能力變化,從而將鼠疫菌基因變異與蚤類媒介吸血行為改變關聯起來[35]。該研究提示鼠疫菌基因組變化可以通過媒介等級聯作用,最終影響鼠疫流行強度。
絕大多數鼠疫菌攜帶3個質粒。其中pMT1質粒上的鼠毒素基因ymt(磷脂酶D)對于鼠疫菌在蚤腸道中的正常存活必不可少。它不僅可以保護鼠疫菌免受跳蚤腸道內含物的細胞毒性影響,而且有利于其在節肢動物腸道中的定殖[39]。ymt活性依賴于生物下調因子rcsA、PDE2(rtn)、PDE3(y3389)[40]和ureD[41]等基因的假基因化,假基因的形成增強了鼠疫菌生物膜的形成能力,以及經蚤傳播的能力[42]。而這些基因所參與的調控子自身表達(如RcsAB)往往又受氣候/環境信號因子(pH值、溫度、代謝物水平、離子濃度)[43]等的調控。適宜的溫度(26 ℃)可使ymt基因的表達量增高[44],利于鼠疫菌的定殖及傳播,從而促進鼠疫流行。
鼠疫菌在自然條件下或傳代過程中,有可能獲得或缺失部分質粒或毒力島,對宿主動物的侵襲能力產生影響[43-44]。以pgm位點為例:該位點是鼠疫菌染色體上一段長度為102 kb的強毒力島[45],兩側存在同向重復的插入元件IS100,可介導鼠疫菌在傳代中發生同源重組及序列缺失[46]。pgm位點的色素沉著區與pgm陽性表型相關,該表型在動物間傳播過程中起主導作用[47];而其攜帶的hmsHFRS操縱子是生物膜在蚤體內形成的關鍵基因[48],該基因的轉錄有利于鼠疫菌生物膜的形成,對跳蚤傳播鼠疫十分重要[49]。現有研究表明鼠疫菌pgm位點在實驗室及自然界傳代過程中均可發生缺失[47],其缺失形成的種群遺傳多樣性將對鼠疫菌在媒介和宿主間的傳播造成影響。
4.2 耶爾森菌屬抗原交叉免疫對生態位的影響 除了鼠疫菌自身外,耶爾森菌屬其他近緣物種的抗原交叉免疫也可能對生態位產生影響。假結核耶爾森菌(Y.pseudotuberoulosis,假結核菌)與小腸結腸炎耶爾森菌(Y.enterocolitica)與鼠疫菌均屬于耶爾森菌屬。三者的基因組分析表明,其在遺傳水平上親緣關系很近[50],均含有共同攜帶的毒力質粒(pYV,在鼠疫菌中被稱為pCD1)編碼的V抗原(LcrV)及外膜蛋白A(outer membrance protein A,OmpA)等多組一致的抗原結構[51]。因而疫源地動物在感染某種致病性耶爾森菌后可能獲得保護性免疫。Harrison等[52]發現,鼠疫菌在自然疫源地中通常與假結核菌、小腸結腸炎耶爾森菌交替出現,隨后鼠疫疫情由流行轉為靜息。高少坤和杜國義[50]比較了鼠疫菌與假結核菌的基因片段和歷史上曾發生的3次世界范圍的人間鼠疫大流行,也發現鼠疫菌與假結核菌在鼠疫流行和靜息期曾交互出現。同一疫源地內動物感染鼠疫菌、假結核菌或小腸結腸炎耶爾森菌的分布此消彼長呈“同心圓”現象(concentric circle distribution)[53]。這一現象間接解釋了在鼠疫自然疫源地的嚙齒類動物中通常無法分離出其他致病性耶爾森菌的現象;反之也提示耶爾森菌屬其他物種的入侵可能是引起生態位變化、疫源地靜息的原因之一。
5 人類活動與生態位
人類活動也會引起動物間鼠疫疫情流行及靜息的交替轉換,比如以滅鼠、滅蚤為主的疫區處理工作,會造成主要宿主動物及蚤類媒介的密度急劇降低。由于疫源地內宿主動物恢復慢,減緩了動物間鼠疫傳播速度,短期內無法發生大規模動物間鼠疫疫情,鼠疫趨于靜息[54]。當疫源地經過多年的自身調節,宿主動物逐漸恢復,達到一定程度后會再次引起鼠疫流行。若動物間鼠疫流行時沒有人為干預(疫區處理),鼠疫可能會波及至人導致人間鼠疫的流行[20]。除此之外,水庫修建等大型施工活動也會破壞鼠疫的生態平衡,我國廣西南盤江流域就曾因修建天生橋水庫而引起動物及人間鼠疫的流行。在建設水庫的十年間,當地未出現過動物間疫情。但隨著水庫的蓄水將周圍農田和植被淹沒,使得原本分散的嚙齒類動物被集中在一個沒有水的地方,鼠疫菌經跳蚤叮咬人后最終引發人間疫情。隨著水庫成功修建以及人類居住環境和動物、植被的逐漸恢復,鼠疫自然疫源地轉入靜息期[3,9]。
6 鼠疫靜息機理的認識
鼠疫菌是生態位中關鍵的一環,因此,了解其在靜息期內如何保存,又以何種機制引起動物間流行對于鼠疫研究而言十分重要。鼠疫菌不僅可以在動物及媒介體內保存,還能在土壤、植物等生態因子中呈非寄生相保存。原生動物是許多病原體的宿主和傳播者[55],其對所吞噬的細菌有選擇作用,研究證實鼠疫菌也可寄生在土壤原生動物中[56]。在冬季或干旱等不利環境下,原生動物通過吞噬土壤中的鼠疫菌形成孢囊,以“休眠”狀態貯藏,保護細菌不受外界環境侵害[56-57]。有些阿米巴個體吞噬鼠疫菌后,被吞噬細菌可在其細胞內大量繁殖,最終使細胞破裂而釋放到外環境中[58]。宋志忠等[59]提出鼠疫菌還可能以植物根系正常菌群的形式保存在根系環境中。當環境內某些因素發生變化時,細菌可通過根部進入植物地上部分,通過攝食途徑感染嚙齒類動物,造成動物間鼠疫的傳播[59-60],但這一推論目前尚缺乏強有力的實驗室和自然條件證據。
7 結 語
動物間鼠疫流行、靜息期交替轉換的現象普遍存在于各鼠疫自然疫源地中。除上文總結的影響因素之外,自然災害、氣候變化等也會對當地動物及媒介生物的棲息環境產生影響。適宜的生態環境使得嚙齒類動物和媒介種群擴增,為鼠疫在動物間的定植、傳播提供機會。而洪澇、干旱、地震等自然災害的發生能短時間內破壞生態位的動態平衡,對鼠疫自然疫源地造成影響,鼠疫可能局部滅絕或進入靜息期,也可能遷徙溢出到其他地區建立新的鼠疫生態位[7-8]。此外,氣溫的季節性變化與宿主動物的活動強度、生殖腺發育、受孕和繁殖等密切相關。溫度過高或過低都能使動物活動強度降低,出現發育遲緩等現象[61-62],間接對鼠疫疫情造成干擾。
盡管動物間鼠疫流行和靜息期轉換研究一直是自然疫源性傳染病領域的重要課題,但目前已知的鼠疫生態位影響因素依然有限,鼠疫流行周期性轉化規律和詳細機制仍不明確。我國作為鼠疫自然疫源地分布和類型最為豐富的國家之一,在面臨鼠疫防控巨大挑戰的同時,也擁有獨特的研究資源。隨著生態學、生物組學、生物信息學等數據密集型學科的飛速發展,將會為鼠疫流行與靜息轉換機制的研究方法和策略帶來新的思路。尤其是將生態學與進化理論(Eco-Evo)相結合的先進研究方法,將使我們從多個維度更加充分的認識鼠疫,進一步揭示鼠疫周期性流行的轉換機制,提高預測預警能力,為鼠疫防控工作提供支撐。
利益沖突:無
猜你喜歡
保健醫苑(2021年7期)2021-08-13 08:48:02
學生天地(2020年36期)2020-06-09 03:12:30
小學科學(學生版)(2020年5期)2020-05-25 07:11:32
小學科學(學生版)(2020年4期)2020-05-21 07:30:46
小學科學(學生版)(2020年3期)2020-03-25 13:31:22
貴茶(2019年3期)2019-12-02 01:46:32
鄉村地理(2018年3期)2018-11-06 06:51:02
茶葉通訊(2017年2期)2017-07-18 11:38:40
河北城市研究(2015年4期)2015-08-23 11:53:06
浙江工商大學學報(2015年6期)2015-03-01 02:56:40
