青稞普通根腐病的調查與病原鑒定
2021-07-19 07:09:04李雪萍劉梅金許世洋郭建煒漆永紅李敏權
草業學報 2021年7期
關鍵詞:研究
李雪萍 ,劉梅金 ,許世洋 ,郭建煒 ,漆永紅 ,李敏權 *
(1. 甘肅省農業科學院植物保護研究所,甘肅蘭州730070;2. 甘肅省甘南州農業科學研究所,甘肅合作747000;3. 甘肅農業大學草業學院,甘肅蘭州730070)
青稞(Hordeum vulgarevar.nudum)是多棱裸粒大麥統稱,在我國主要種植于西藏、青海、云南、甘肅、四川等省區海拔較高的地區,其成熟期短、耐寒性強,在海拔超過4200 m 的高寒地區,青稞是唯一能夠正常成熟的谷物,因此,在古埃及和我國的藏區也常被當地居民作為主食[1]。同時,青稞在澳大利亞、歐洲馬其頓、美國北部大平原、加拿大西部等多個國家地區及我國多個地區常被作為飼用作物,其富含多種氨基酸、膳食纖維、維生素、β?葡聚糖以及鈣、鎂、磷、鋅、錳、硒等礦質元素,亦常被用來釀酒及現代健康食品的加工等[2]。由此可見,青稞的生產不僅保障了高原地區畜牧業的發展及當地居民的基本生活,更為現代人民健康生活水平的提升做出了貢獻。
然而,植物病害嚴重影響了包括青稞在內的大麥類作物的生產。據Murray 等[3]報道,澳大利亞在1998?2007 年這10 年間,由植物病害造成大麥(Hordeum vulgare)損失每年達2.52 億美元,是總產值的19.6%。在美國北達科他地區及加拿大,根腐病造成大麥減產近10%[4]。青稞感染根腐病后根和芽的鮮重、干重、谷粒數均大幅下降,從而導致總產量降低;分析其營養成分發現,其總碳水化合物下降23.28%~82.77%,蛋白質下降0.55%~76.62%,脂肪下降28.45%~79.25%[5]。目前,對大麥類作物病害的研究主要集中在大麥條紋病[6?7]、銹病等[8?9]方面,對于根腐病的研究較少,對青稞根腐病的研究僅有本課題組報道[10?11],而根腐病病害類型復雜,病原多樣,因寄主、環境、氣候、土壤等各種因素而異。鑒于此,本研究對西北青藏高原地區的青稞根腐病進行了調查,明確其危害程度及發病癥狀,并采集樣品進行病原分離鑒定、致病性測定,以期為青稞根腐病的防控及致病機理的研究提供理論依據。
1 材料與方法
1.1 調查研究區概況
調查研究區選擇甘肅省甘南藏族自治州合作市、卓尼縣、臨潭縣以及青海省海北藏族自治州剛察縣、海東市互助土族自治縣等我國重要的青稞生產區。其位于青藏高原東部及北部邊緣,平均海拔2800 m 以上,無霜期短,日照時間時長,是典型的大陸性氣候,青稞是該地區適生作物,也是該地區最主要的糧飼作物。
1.2 調查采樣
參考《植病研究法》[12]對該區域青稞發病率進行調查統計,避開地邊5 m,挑選植株枯黃或長勢明顯很弱,且發病植物地下部普遍有根腐癥狀的地塊,統計其占總地塊的面積比例,即其發病率,據此計算同一地區所有地塊的平均發病率,即該地區青稞普通根腐的發生率;采用多點采樣法采集發病特征典型的疑似青稞普通根腐病植株,低溫運輸至實驗室進行研究。
1.3 病原的分離與純化
刷掉青稞病株根部土壤,清洗干凈并自然風干。從根部的病健交界處剪下根段,70%的酒精消毒2~3 s 并用無菌水沖洗干凈,放入0.1%的升汞溶液中消毒10 s,再用無菌水洗4 次,用無菌濾紙吸干表面水分;然后用無菌解剖刀切去根段兩端,將中間的根段接于馬鈴薯葡萄糖瓊脂(potato dextrose agar,PDA;培養基成分:馬鈴薯200 g,葡萄糖20 g,瓊脂20 g,水1000 mL)平板之上;約3~4 d 后,將根段的兩端長出的菌絲轉移到另一PDA 平板之上,并采用單孢分離法[13]對其進行純化。
1.4 菌株鑒定
1.4.1 形態鑒定 觀察并記錄菌株在PDA 培養基上的菌落形態特征,在顯微鏡下觀察其菌絲及孢子形態,參照《真菌鑒定手冊》[14]、《中國真菌志》第十六卷(鏈格孢屬)[15]、第三十卷《蠕形分生孢子真菌》[16],確定其分類地位。
1.4.2 分子生物學鑒定 將待鑒定菌株活化后接于馬鈴薯葡萄糖培養液中,120 r·min?1、25 ℃搖床培養5 d,收集菌絲體,Fungal DNA Kit 試劑盒(OMEGA)提取DNA(操作步驟依據試劑盒說明書)。選用ITS1(5′-TCCGTAGGTGAACCTGCGG-3′)和 ITS2(5′-TCCTCCGCTTATTGATATGC-3′)引物進行 PCR 擴增。反應體系為:總體積 25 μL、基因組 DNA 1.0 μL、10×Buffer(含 2.5 mmol·L?1Mg2+)2.5 μL、Taq 聚合酶(5 U·μL?1)0.5 μL、dNTP(10 mmol·L?1)1.0 μL、Primer(±10 μmmol·L?1)1 μL、ddH2O 補至 25 μL。PCR 參數為:94 ℃預變性 5 min,94 ℃變性 30 s,58 ℃退火 45 s,72 ℃延伸 80 s,重復擴增35 個循環,72 ℃延伸 7 min,4 ℃保存。反應完成后,取2 μL PCR 產物進行1%瓊脂糖凝膠電泳檢測,檢測合格后委托北京天一輝遠生物科技有限公司進行測序。得到測序結果后在NCBI 中進行Blast 比對并選取參考序列,利用MEGA 7.0 中的最大似然法(1000次重復)構建系統發育樹,確定其分類地位,并將結果提交至GenBank,獲得登錄號。
1.5 致病性測定
根據前期研究結果,燒杯水瓊脂法可以較好、較便捷的反應菌株的致病性[13],具體為:1)將待測菌株活化后接種于1.2%的水瓊脂(water agar,WA)培養基上,置于25 ℃培養3 d。2)將供試種子(品種為藏青2000,由甘南州農業科學研究所提供)消毒后25 ℃催芽48 h。3)挑取發芽一致的種子均勻擺放在帶菌的WA 培養基上,并在不接菌的WA 培養基上接入種子作為對照,每杯放入10 粒,錫紙封口后放入人工氣候箱(兩個階段:第一階段為時間8 h,光80%,濕度60%,溫度25 ℃;第二階段為時間16 h,光照為0,濕度80%,溫度20 ℃)。待種苗長至燒杯口處時揭去錫紙,打開紙盒蓋,繼續在人工氣候箱中培養,8 d 后觀察發病情況,參考田間發病情況及發病癥狀,確定分級標準,統計病級,計算其發病率及病情指數。
分級標準:0 級?健康無病,葉綠,根與根莖白;1 級?根系有褐色斑點出現,但不超過30%;2 級?根系成段變褐,但不斷裂,不超過30%;3 級?根系成段變褐甚至變黑,容易斷裂,不超過50%;4 級?根系成段變褐甚至變黑,斷裂腐爛,大于50% 或整個植株死亡。

將致病性測定后病株每菌隨機挑出3 株,剪下其種子或病根,接種于PDA 培養基上,于25 ℃下培養7 d 后鏡檢觀察其孢子形態,在此期間不斷觀察其菌落形態,確認是否與接入時的菌株一致。
1.6 數據分析
相關數據整理及分析采用Excel 2007 和DPS 15.10 完成,采用Duncan 新復極差法進行差異顯著性分析(P<0.05)。
2 結果與分析
2.1 發生分布
青稞普通根腐病主要分布在甘肅省甘南藏族自治州臨潭縣古戰鄉,卓尼縣柳林鎮、阿子灘鄉、完冒鄉,合作市卡加曼鄉以及青海省海北藏族自治州剛察縣,海東市互助土族自治縣林川鄉。發生較為普遍,田間發病率為5%~15%(表1)。
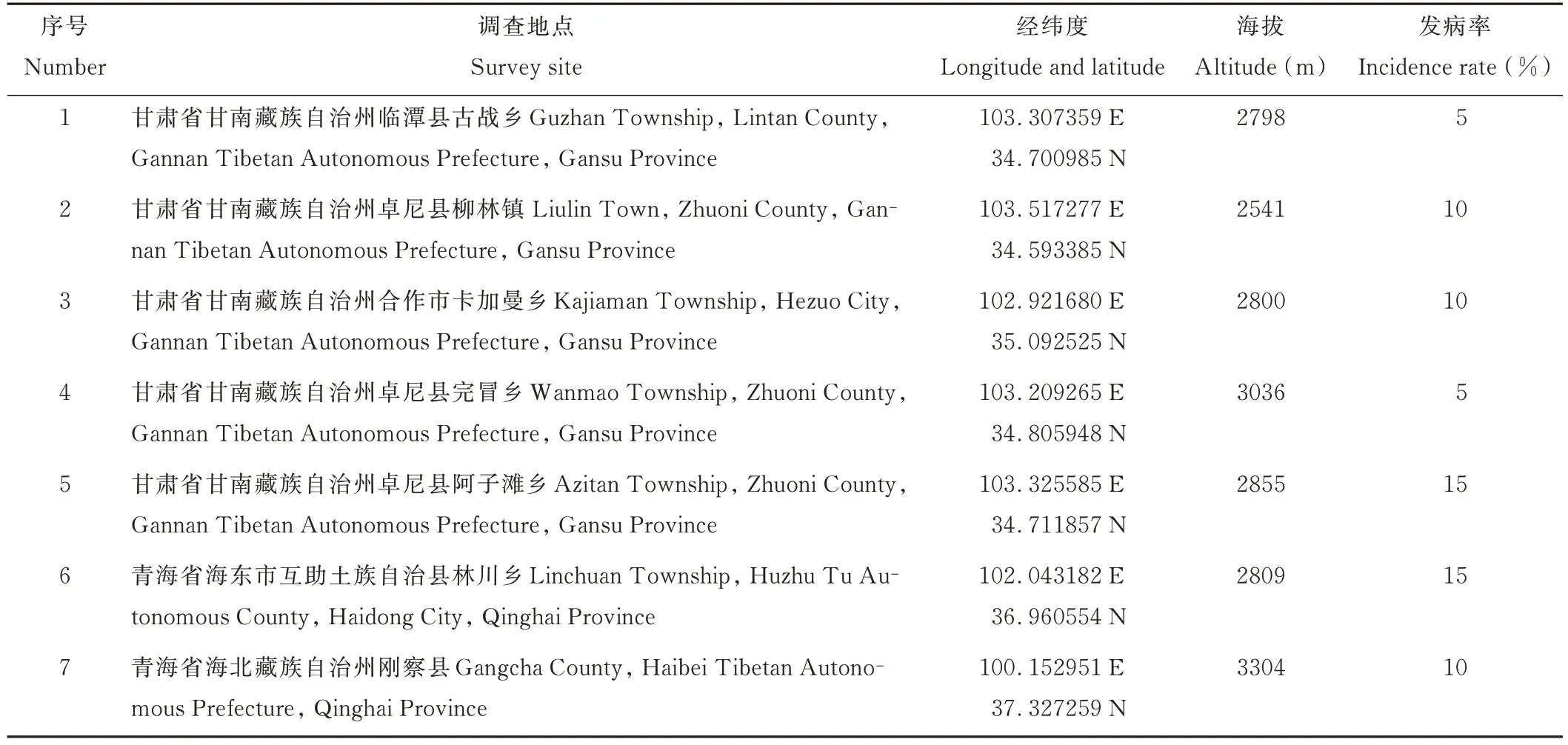
表1 青稞根腐病的調查Table 1 Investigation on root rot of qinke barley
2.2 發病癥狀
青稞苗期普通根腐病不易識別,其癥狀為地上葉片多表現為葉色淡,呈黃綠色,幼苗瘦弱或死亡,有明顯的發病中心,挖出洗凈后可發現其根部發黑,容易斷裂或腐爛;青稞成株期普通根腐病多表現為穗白粒癟,莖稈甚至葉片呈黑褐色,挖出洗凈后亦可發現根部黑褐斷裂或腐爛(圖1)。

圖1 青稞普通根腐病癥狀Fig.1 Qingke barley common root rot symptoms
2.3 病原鑒定
2.3.1 形態特征 共分離得到13 株青稞普通根腐病病原,其中8 株在PDA 培養基上均呈現青灰褐色,氣生菌絲發達,厚絨狀,易產生大量分生孢子,孢子呈淡黃褐色至灰褐色,易從兩端萌發,擬紡錘形,多數較直,少數彎曲,平均大小為(63.62~79.36)μm×(18.99~22.46)μm(圖2),初步鑒定為麥根腐平臍蠕孢(Bipolaris sorokiniana)。
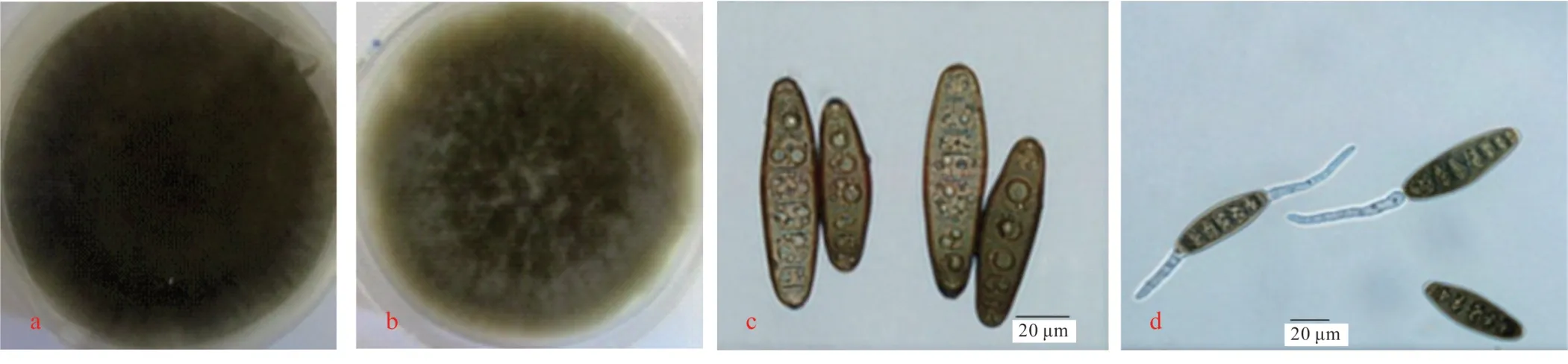
圖2 麥根腐平臍蠕孢的形態特征Fig.2 Morphological characteristics of B. sorokiniana
其中5 株菌落呈青灰褐色,菌絲發達,在PDA 培養基上容易產生大量的分生孢子,分生孢子黃褐色至青灰褐色,形狀大小不一,有橫隔膜,斜隔膜和縱隔膜,平均大小為(24.86~41.66)μm×(8.32~13.92)μm(圖3)。初步鑒定為鏈格孢(Alternaria alternata)。
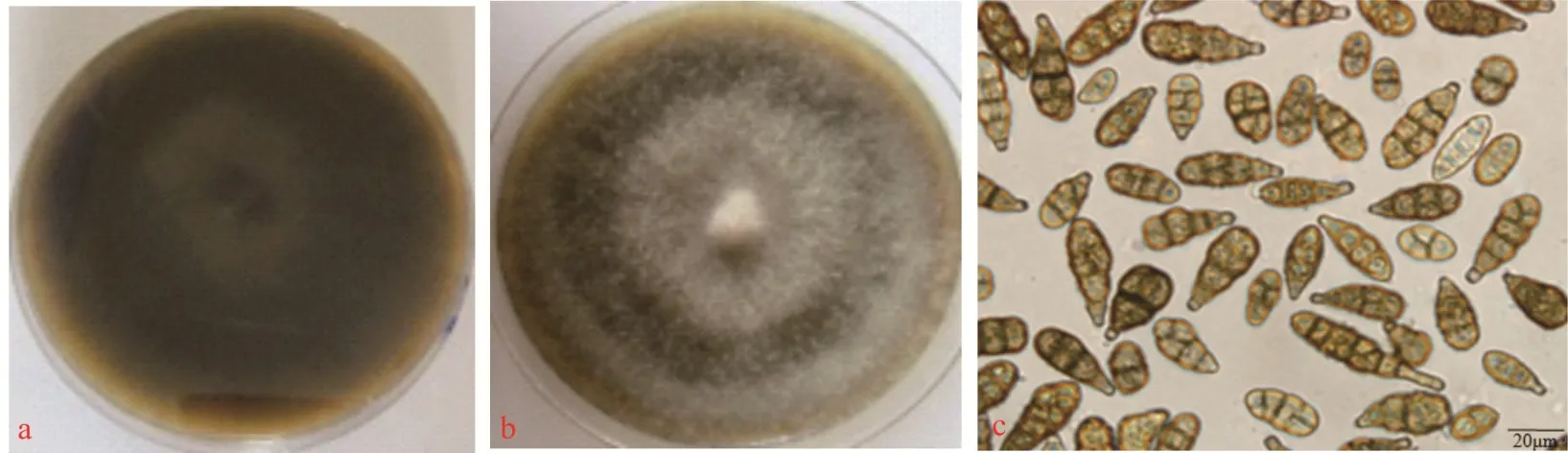
圖3 鏈格孢的形態特征Fig.3 Morphological characteristics of A. alternate
2.3.2 分子鑒定 如圖4 所示,以木賊鐮孢及鏈格孢等作為外群,構建系統發育樹發現,D26-2、D23-1、D22-1、D13-1、D32-6、D28-3、NQ4-3、Q7-6 這 8 株菌與麥根腐平臍蠕孢聚為一類,遺傳距離為 0,自展支持值為 100(1000 次重復)。結合形態特征鑒定的結果,確定本研究分離到的此8 株菌均為麥根腐平臍蠕孢。提交序列至GenBank,獲得的登錄號分別為 KY365567(D26-2)、KY365566(D23-1)、KY365565(D22-1)、KY365575(D13-1)、KY365569(D32-6)、KY365568(D28-3)、KY365583(NQ4-3)、KY365590(Q7-6)。

圖4 基于rDNA ITS 區序列構建的麥根腐平臍蠕孢系統發育樹Fig.4 B. sorokiniana phylogenetic tree based on rDNA ITS sequence
同時,以麥根腐平臍蠕孢作為外群,構建系統發育樹發現,D28-1、D34-1、D3-4、NQ3-4、D33-2 這5 菌株與鏈格孢的遺傳距離為0,自展支持值大于99(1000 次重復)。結合形態特征,確定該5 株菌為鏈格孢。提交序列至GenBank,獲得的登錄號分別為 KY365561(D28-1)、KY365563(D34-1)、KY365573(D3-4)、KY365582(NQ3-4)、KY365562(D33-2)(圖 5)。

圖5 基于rDNA ITS 區序列構建的鏈格孢系統發育樹Fig.5 A. alternate phylogenetic tree based on rDNA ITS sequence
2.4 致病性分析
如表2 所示,8 株麥根腐平臍蠕孢均可導致青稞普通根腐病的發生,發病癥狀與青稞田間苗期普通根腐病癥狀一致,除D13-1 及NQ4-3 外,其他6 株菌可100%引起青稞普通根腐病的發生,其中D32-6、Q7-6、D26-2 和D28-3 這4 株菌的致病性最強,病情指數均在80%以上,且各菌株的致病性差異顯著。經再次分離病原觀察其形態特征發現,與接入時一致,確認為該種病害的病原。

表2 青稞普通根腐病病原-麥根腐平臍蠕孢的致病性Table 2 The common root rot pathogen of qinke barley-B.sorokiniana pathogenicity
如表3 所示,5 株均有致病性,致病差異顯著;鏈格孢的致病性較低,致病性最強的D28-1 發病率為89.17%,病情指數為36.42;發病癥狀與田間苗期普通根腐病輕癥一致,經再分離發現,其菌株形態與接入時一致,確認為普通根腐病病原。
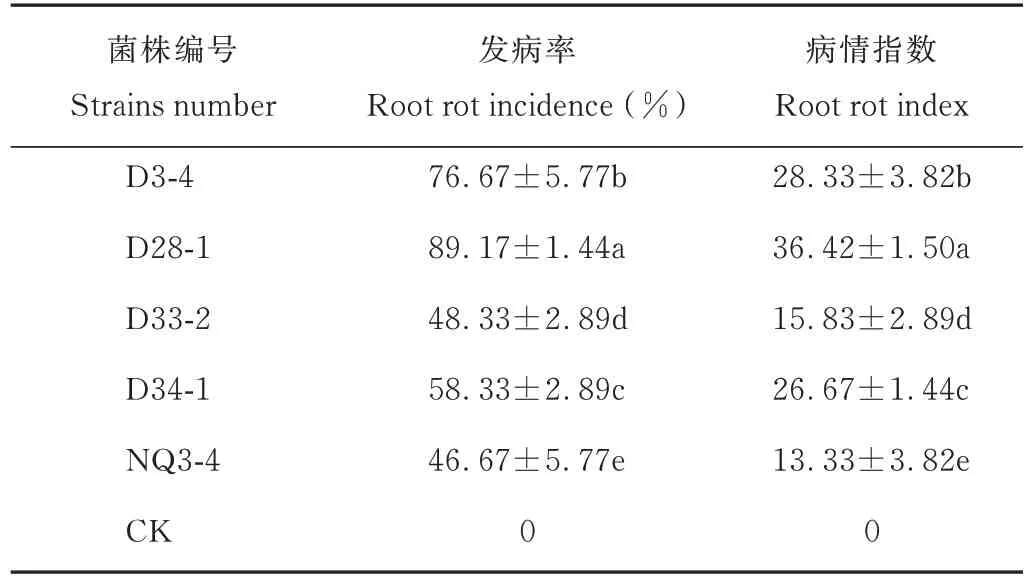
表3 鏈格孢的致病性Table 3 The A.alternate pathogenicity
3 討論
目前,關于青稞根腐病的研究較少,病原研究鮮見,可查閱到的資料多為大麥根腐病的相關研究,并非其變種青稞,因此,本研究首次明確了青稞普通根腐病的病原。本研究發現,青稞普通根腐病的優勢病原是麥根腐平臍蠕孢,與 Ilija 等[17]、Mohammad 等[18]和Zhong 等[4]對大麥根腐病病原的研究結果一致。也有眾多研究表明大麥根腐病的優勢病原為鐮孢菌,主要包括燕麥鐮孢、黃色鐮孢、禾谷鐮孢、梨孢鐮孢、擬枝鐮孢等[19?22]。然而,有研究發現,大麥根腐病是由多種病原復合侵染引起的,如 Makela 等[23]、Pua 等[24]和 Fedel 等[25]研究表明,大麥根腐病是由麥根腐平臍蠕孢和鐮孢菌屬共同引起的。Amira[26]和 Ravjit 等[27]則研究表明,絲核菌和鐮孢菌是引起大麥根腐病的優勢病原。前人對大麥根腐病病原的研究做出了巨大貢獻,但未見將根腐病害類型作以區分。究其原因,根腐類病害發生的環境復雜,受氣候、種植、土壤等多個因素的影響,不同類型的根腐類病害往往混合發生,癥狀類似,從而對根腐類病害類型的確定造成了困難。本研究通過大量調查采樣及試驗研究發現以麥根腐平臍蠕孢為優勢病原的青稞根腐類病害,其癥狀典型,與鐮孢菌根腐病等其他根腐類病害差異較大,確定為青稞普通根腐病。麥根腐平臍蠕孢是小麥(Triticum aestivum)、玉米(Zea mays)、柳枝稷(Panicum virgatum)等植物葉斑病[28?30]、根腐病[31]以及燕麥(Avena sativa)褐斑病[32]等多種作物病害的病原,其侵染規律、致病機理及各種病害之間的關系有待進一步研究。另外,近年來,越來越多的研究證明,鏈格孢能引起葉斑病、果腐病、根腐病、黑斑病等多種植物病害,其寄主廣泛,致病性強,產生毒素,危害嚴重[33?34]。本研究發現,5株鏈格孢可致病導致青稞根腐病的發生,但未見只分離到鏈隔孢的病株,其可能與麥根腐平臍蠕孢復合侵染或在麥根腐平臍蠕孢侵染后加重發病程度,其互作機理還有待進一步研究。
4 結論
我國西北青藏高原地區甘肅省甘南藏族自治州、青海省海東市、海北藏族自治州青稞普通根腐病發生普遍,發病率為5%~15%。青稞感染普通根腐病后葉呈黃綠色,穗白粒癟,幼苗瘦弱或死亡,根部發黑、腐爛或斷裂。青稞普通根腐病的病原為麥根腐平臍蠕孢和鏈格孢,麥根腐平臍蠕孢為優勢病原。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
