日本無針烏賊胚胎期眼部發育相關基因的研究
2021-07-21 09:28:40李少剛李永芹許樂樂陳道海王鋰韞
水產科學 2021年4期
李少剛,李永芹,2,許樂樂,2,陳道海,2,王鋰韞,2,3
( 1.嶺南師范學院 生命科學與技術學院,廣東 湛江 524048; 2.廣東省粵西海鮮資源可持續利用工程技術研究中心,廣東 湛江 524048; 3.廣東省特殊兒童發展與教育重點實驗室,廣東 湛江 524048 )
日本無針烏賊(Sepiellajaponica)屬軟體動物門、頭足綱、十腕目、烏賊科、無針烏賊屬,因其高度進化、發展并集中的神經系統,被認為是研究眼睛形態發育的模式生物[1]。在胚胎發育過程中,有關頭足類動物光敏度的研究表明,在孵化前,胚胎在視網膜最終分化之前會變得光敏[2-3]。與其他頭足類動物不同,烏賊卵周圍有黑色的包膜,從而減弱了進入胚胎的光線,烏賊胚胎會在黑暗的視覺環境中發育,當眼部器官分化到第一個視網膜色素出現的時期,烏賊胚胎對光照做出反應[4-8]。目前針對日本無針烏賊胚胎期眼部發育的研究多集中于外源性因素,如光照、飼料、藥物等,而關于轉錄水平調控機制對日本無針烏賊胚胎期眼部分化的影響卻鮮有報道。
多細胞生物視覺結構的形態發生受到一個保守的基因轉錄控制網絡所調控[9]。在基因轉錄控制網絡中,paired-box 6(pax6)是高度保守的轉錄因子家族paired-box的成員之一[10-11],也被認為是控制眼睛形態發生的通用基因,有研究表明,其在多細胞生物眼睛光敏感受器的發育過程起到重要作用[12-13]。除pax6之外,SIX Homeonox 3 (six3)、Eyes Absent (eya)基因也參與了多細胞生物視細胞的增殖、視網膜前體的分化、光敏感受器神經元的分化和維持,以及其他非視網膜組織和器官的發育過程[14-17]。筆者在日本無針烏賊胚胎不同發育期轉錄組測序分析的水平[18]上進一步探討與日本無針烏賊胚胎眼部發育過程相關調控基因six3、eya及pax6的表達變化。
不同研究者對烏賊胚胎發育的分期各有不同,劉振勇等[19]將烏賊受精卵到出膜幼體分為22期;蔣霞敏等[20]從胚胎的受精卵階段至孵化階段將烏賊卵的發育過程分為12個階段;von Boletzky等[21]則將從受精卵到形態結構與成體基本相似的烏賊的胚胎發育分為30個時期。綜合參考上述分期方法,筆者擬將日本無針烏賊胚胎發育過程分為3個時期、8個階段,并對不同發育階段烏賊胚胎中的six3、eya、pax6基因相對表達水平進行分析。
剛孵化的烏賊對視覺或氣味的提示會立即做出反應并能表現出對某些重復視覺刺激的習慣,表明烏賊胚胎在孵化之前的發育過程中便能探測到刺激[6,22]。筆者探討分子水平日本無針烏賊胚胎期眼部發育相關調控基因的表達變化,旨在查明烏賊相關調控基因對眼睛發育的影響機制,為探究烏賊的視覺機制,如烏賊快速適應偽裝的視覺機制,同時為日本無針烏賊胚胎的人工繁育和增養殖提供分子水平數據參照。
1 材料與方法
1.1 樣品采集
當天產卵的日本無針烏賊卵采自中國南海汕頭海域,轉移至實驗室烏賊孵化養殖缸中培養,養殖缸規格0.4 m×0.2 m×0.3 m,烏賊卵附著于尼龍繩上, 并在下方用塑料籃盛起,24 h人工增氧。孵化用水為砂濾海水,水溫(28±1) ℃,鹽度27±1,pH 7.5~8.2,連續充氣,每日換水,換水量為養殖缸的1/3[23]。
自獲得烏賊卵當日起,觀察和記錄每日培育狀況及養殖環境。同時,隨機挑取3~5顆烏賊卵,在體視顯微鏡(Nikon SMZ800N,Tokyo,Japan;目鏡C-W10×B/22,物鏡 Plan Apo 1×WF WD:70)下觀察和記錄,并進行分期及細分階段。每個階段取50顆烏賊卵,迅速用預冷的PBS溶液(Rnase free)沖洗,然后將每個卵分別保存于液氮預冷的滅菌密封袋中,立即置于液氮中冷凍3 h,置于-80 ℃超低溫冰箱凍存備用。
1.2 總RNA提取及cDNA反轉錄
每個階段的日本無針烏賊胚胎各取6顆,于研缽中加液氮充分碾磨,每個胚胎轉移100 mg粉末于1.5 mL離心管中,采用RNAiso Plus (TaKaRa, 日本)試劑盒提取總RNA,提取方法依據說明書進行操作。經微量核酸測定儀(NanodropTMOne, Thermo ScientificTM, 美國)及凝膠電泳測定RNA質量濃度及純度后,使用反轉錄試劑盒 (Transcriptor First Strand cDNA Synthesis Kit, Roche,瑞士)對RNA進行反轉錄,反轉錄方法依據說明書進行操作,反轉錄后cDNA置于-80 ℃冰箱中保存備用。
1.3 引物設計
根據已構建的日本無針烏賊轉錄組測序及注釋信息[18],結合美國國立生物技術信息中心數據庫中已報道的軟體動物基因組序列,采用Primer Premier 5.0軟件對six3、eya、pax6、gapdh(內參)、rpl23a(內參)基因進行引物設計,由通用生物公司設計合成,引物序列見表1。
1.4 熒光定量PCR
以提取到的8個階段烏賊樣品的cDNA(與轉錄組取樣一致,每個胚胎取100 mg)作為模板(n=6),以gapdh和rpl23a作為內參基因,使用PowerUpTMSYBRTMGreen Master Mix (Thermo Fisher Scientific, Foster, USA),通過BioRad公司的熒光定量PCR儀(CFX96 Touch, California, USA) 對six3、eya、pax6基因在不同階段的烏賊胚胎中的相對表達水平進行檢測。反應程序為:95 ℃ 10 min;95 ℃ 15 s,60 ℃ 1 min,40 個循環; 95 ℃ 15 s,60 ℃ 1 min,95 ℃ 30 s。每個樣品及內參均設置 2個平行,采用GraphPad Prism 7軟件對數據進行處理,并根據Ct值,用2-ΔΔCt法對結果進行分析。
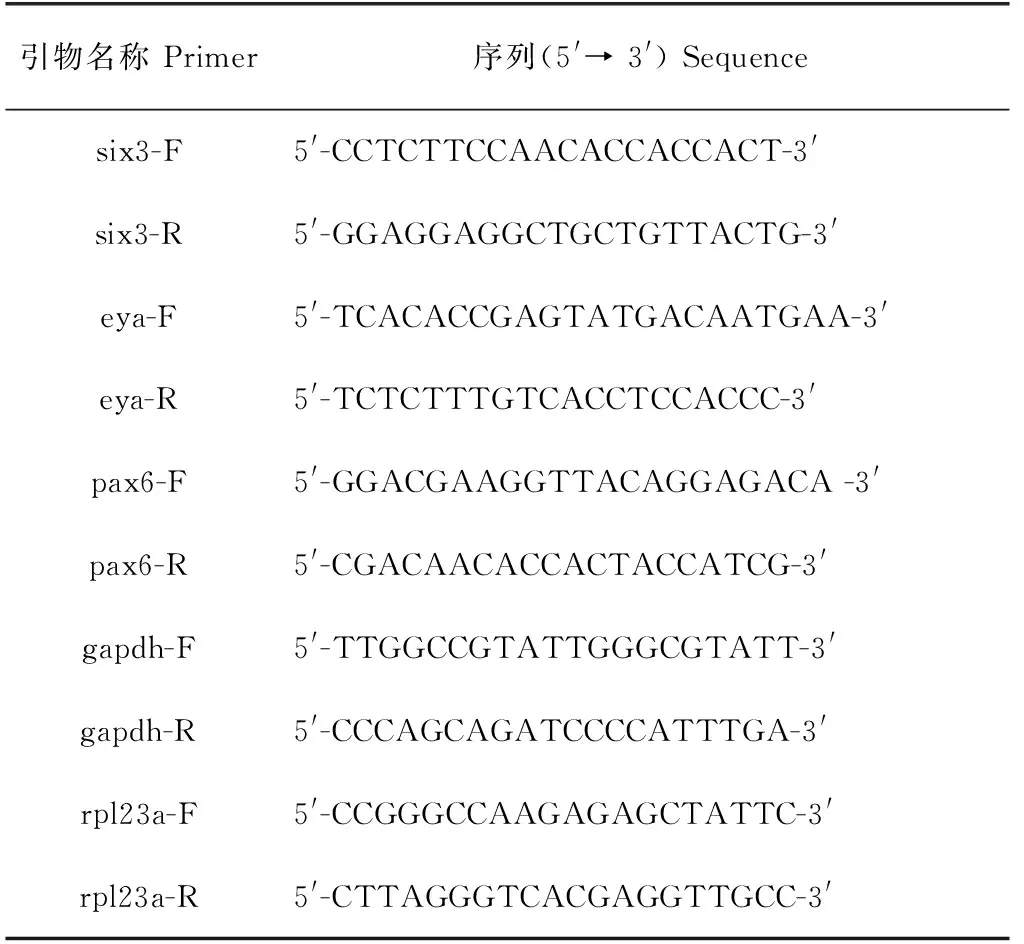
表1 試驗用引物序列
1.5 統計分析
對獲得的熒光定量結果采用SPSS 23軟件進行統計分析,差異顯著水平檢驗采用單因素方差分析,*表示差異顯著,其中,*表示P<0.05,**表示P<0.01,***表示P<0.001。
2 結果與分析
2.1 日本無針烏賊胚胎發育過程
根據日本無針烏賊胚胎發育特征(圖1),及基因表達分析研究,將日本無針烏賊胚胎發育劃分為3個時期[受精卵—眼原基形成期(SJ1)、功能器官分化期(SJ2)、幼體孵化期(SJ3)]、8個階段[受精卵—胚芽原基及胚體形成(SJ1-1),口、漏斗原基出現(SJ2-2),眼點形成(SJ2-3),紅眼(SJ2-4),墨囊形成(SJ2-5),黑眼(SJ2-6),內殼形成(SJ2-7),剛孵化幼體(SJ3-8)](表2)。
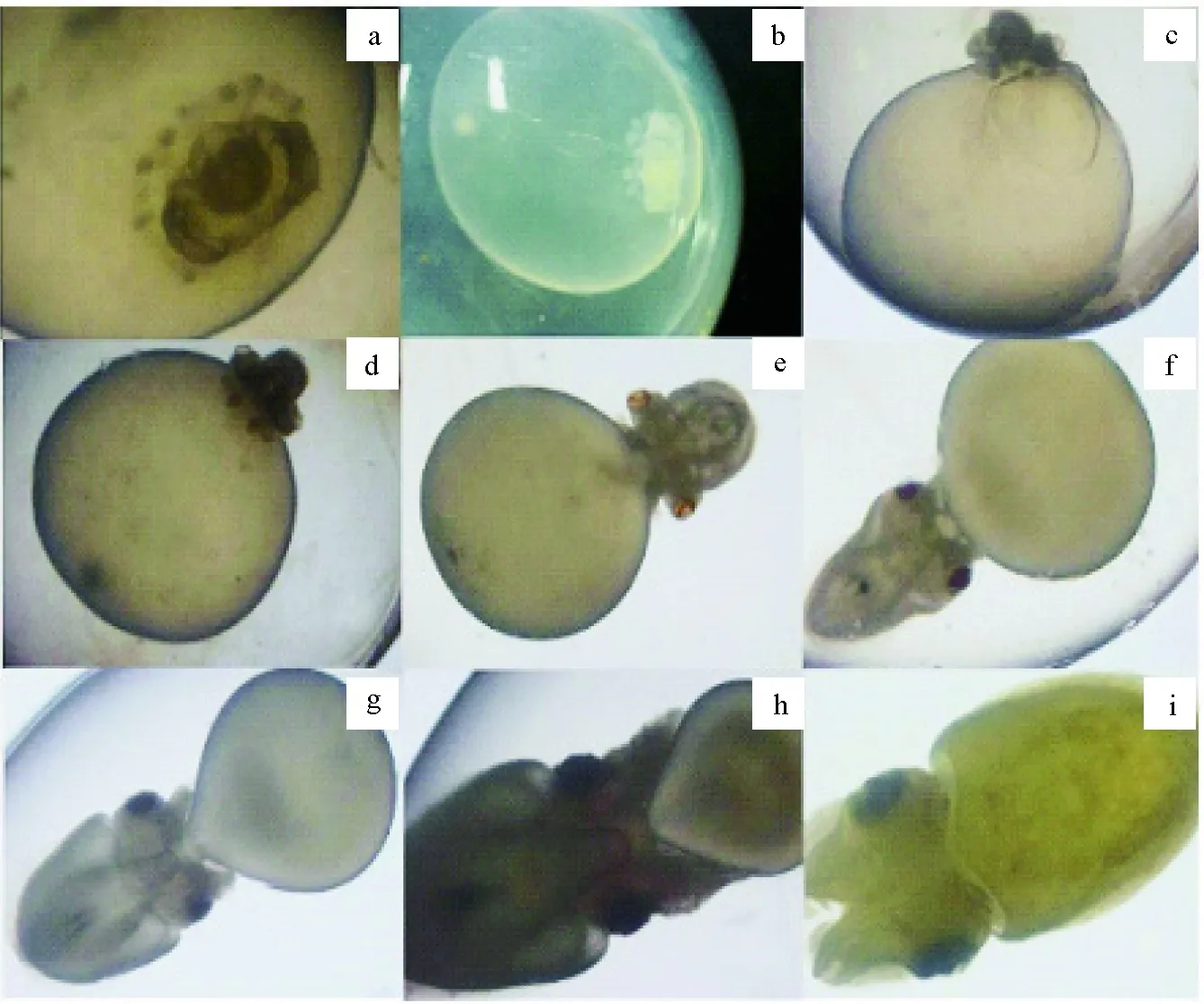
圖1 日本無針烏賊胚胎發育過程特征觀察Fig.1 Observation of the developmental characteristics of embryos in Japanese spineless cuttlefish S. japonicaa~b.受精卵—眼原基形成期(SJ1-1); c~h.功能器官分化期(c.SJ2-2; d.SJ2-3; e.SJ2-4; f.SJ2-5; g.SJ2-6; h.SJ2-7); i.幼體孵化期 (SJ3-8);放大倍數10×.a—b.fertilized egg-eye primordium formation period (SJ1-1); c—h.functional organ differentiation period (c.SJ2-2; d.SJ2-3; e.SJ2-4; f.SJ2-5; g.SJ2-6; h.SJ2-7); i.larval incubation period (SJ3-8);magnification 10×.

表2 日本無針烏賊胚胎發育過程
2.2 日本無針烏賊胚胎不同發育階段six3、eya、pax6基因表達分析
基因表達分析結果表明,以gapdh、rpl23a為內參,six3、eya基因表達量均隨烏賊胚胎發育從受精卵—眼原基形成期到幼體孵化期3個時期8個階段逐漸增加;six3基因在幼體孵化期第8階段相對表達量相比受精卵—眼原基形成期第1階段及功能器官分化期6個階段均顯著增加,功能器官分化期第7階段相比受精卵—眼原基形成期第1階段及功能器官分化期第2、3階段均顯著增加;eya基因在幼體孵化期第8階段相對表達量相比受精卵—眼原基形成期第1階段及功能器官分化期第2階段顯著增加,功能器官分化期第7階段相比受精卵—眼原基形成期第1階段顯著增加。pax6基因相對表達量隨烏賊胚胎發育從受精卵—眼原基形成期到功能器官分化期有所增加,但從功能器官分化期第2階段開始直至幼體孵化期第8階段基因表達量逐漸降低;以gapdh基因為內參,pax6基因在幼體孵化期第8階段相對表達量相比受精卵—眼原基形成期第1階段及功能器官分化期第2~6階段顯著降低,而以rpl23a基因為內參,3期相對表達量無顯著差異(圖2)。
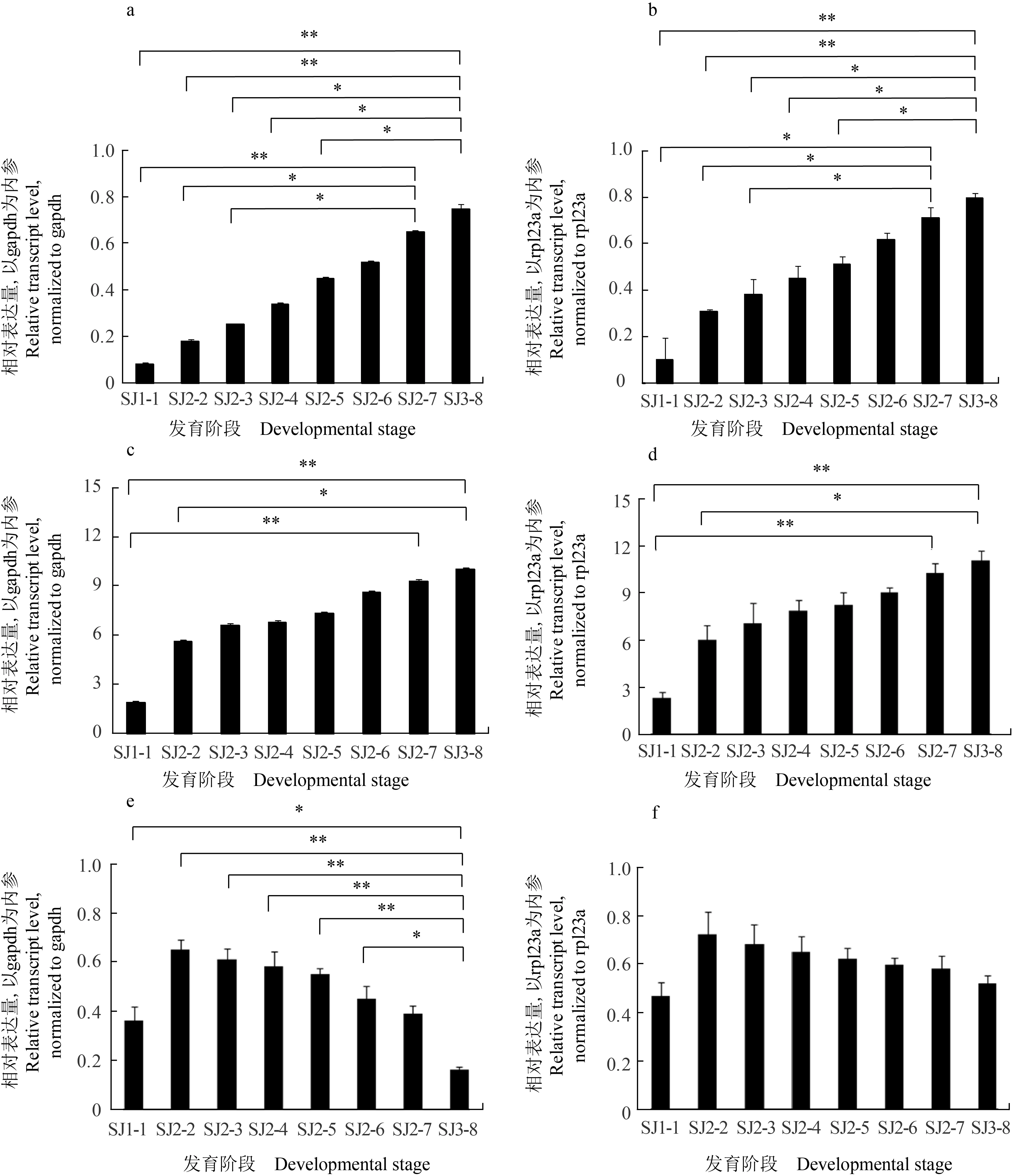
圖2 日本無針烏賊不同胚胎發育階段six3、eya及pax6基因相對表達量(n=6)Fig.2 The relative transcript levels of six3,eya,and pax6 genes in different embryonic developmental stages of Japanese spineless cuttlefish S. japonica a~b.six3基因; c~d.eya基因; e~f.pax6基因; 圖中數值為平均值±標準差;*.P<0.05, **.P<0.01.a—b.six3 gene; c—d.eya gene; e—f.pax6 gene; values are expressed as the mean±SD; *.P<0.05, **.P<0.01.
3 討 論
3.1 six3、eya及pax6基因在日本無針烏賊胚胎發育期中的表達量
本項目組日本無針烏賊轉錄組數據庫[18]使用短reads組裝軟件Trinity組裝拼接后獲得58 054條Unigene,這些Unigene中,作為研究對象的six3基因,其Unigene長度為1220 nt,GC含量為48.61%;eya基因,其Unigene長度為4194 nt,GC含量為39.96%;pax6基因,其Unigene長度為1036 nt,GC含量為37.45%。根據每百萬Reads中來自某一基因每千堿基長度的Reads數目(RKPM)值計算,six3基因在3個時期中的表達RPKM均值為1.06;eya基因在3個時期中的RPKM均值為0.99;pax6基因在3個時期中的RPKM均值為0.40。根據高原等[24]對RPKM值劃分定義基因表達高低的方法,將基因表達量分為以下4個水平:中高表達量RPKM(30~+∞)、偏低表達量RPKM(3~30)、低表達量RPKM(1~3)、超低表達量RPKM(0~1)。本試驗中,日本無針烏賊胚胎eya基因及pax6基因在3個時期RPKM均值均在超低表達量RPKM(0~1)的區間中,即eya、pax6基因在胚胎發育3個時期的表達量均為超低表達量,six3基因在3個時期RPKM均值在低表達量RPKM(1~3)的區間中,即six3基因在胚胎發育3個時期的表達量均為低表達量,因此推測six3、eya、pax6 3個基因在基因轉錄控制網絡中以較低表達量進行調節作用,如six3以較低表達量調節視網膜發育,而過表達則可引起某些特定細胞亞群的擴增[25-29]。
3.2 雙內參基因gapdh、rpl23a在six3、eya及pax6基因表達分析中的應用
根據Pfaffl等[30-31]基因穩定性測量方法,本試驗在基因表達分析中,采用雙內參基因gapdh、rpl23a對數據進行歸一化處理。在six3、eya基因表達分析中,以gapdh及以rpl23a為內參基因結果一致,內參基因受候選基因表達強度影響較小,而在pax6基因表達分析中,以rpl23a為內參基因,所分析各時期樣品的表達水平變化較以gapdh為內參基因各時期樣品的表達水平變化大,但無顯著性差異出現,表達趨勢一致。目前有關日本無針烏賊基因表達分析內參基因選擇報道較少,該結果有助于日本無針烏賊基因表達分析選擇穩定的參考基因。
3.3 six3、eya及pax基因在日本無針烏賊眼睛發育中的調控作用
脊椎動物眼形態的發生受到包括six、eya及pax在內的幾個重要基因的調控,它們在脊椎動物和果蠅的視網膜前體細胞分化及增殖,光感受器神經元細胞的維護中起著關鍵作用[32]。Koenig等[33]研究顯示,與脊椎動物一樣,頭足類動物中six3及eya基因參與晶狀體的形成,頭足類動物中six3及eya基因在胚胎發育早期眼區(如胚胎的眼基、視葉)及在胚胎發育后期胚基板和蓋的邊緣可觀察到有明顯表達。本試驗中,six3、eya基因在日本無針烏賊不同胚胎發育時期及階段的相對表達量結果顯示,six3、eya基因在眼原基形成期有微量表達,并隨胚胎發育,從眼原基形成期到幼體孵化期8個階段相對表達量逐漸顯著增加,表明six3、eya在日本無針烏賊眼睛發育中同樣起重要作用。
在脊椎動物胚胎發育期,pax6基因在視網膜發育后期的腺管和神經節細胞中表達[34]。在魷魚胚胎發育期,pax6基因被認為是眼形成通用主控基因,參與光感器官的發育,pax6基因在加州魷魚(Loligoopalescens)眼睛發育過程中的表達方式與果蠅無眼基因表達方式類似,無眼基因在果蠅眼盤未分化前體細胞中表達活躍,在眼盤后端形成形態溝后的分化細胞中表達明顯下降,pax6基因在加州魷魚光區外胚層和中胚層表達明顯,而在分化的視網膜細胞中無表達[35]。本試驗中,pax6基因在日本無針烏賊胚胎發育功能器官分化期表達量增加,在眼點形成前期眼柄突出時表達最活躍,隨胚胎發育視網膜細胞分化表達逐漸降低,與加州魷魚中pax6基因表達相似,而pax6基因在日本無針烏賊眼睛發育后期調控與脊椎動物不同[36]。
3.4 six3、eya、pax6基因在基因轉錄控制網絡調控網絡中的調節作用
six3、eya、pax6基因在日本無針烏賊眼原基形成期、功能器官分化期及幼體孵化期3個胚胎發育時期8個階段不同程度的表達水平表明,這3個基因在基因轉錄控制網絡中發揮不同調節作用。有研究表明,eya基因表達的EYA蛋白是一類重要的轉錄調控因子,其在胚胎發育過程中對組織和器官的特異性分化具有重要調控作用[37-38],EYA蛋白可以與SIX蛋白發生直接相互作用[39-40],作為SIX蛋白的共激活因子激活SIX[41],因而推測基因功能關聯性可能與six3、eya基因相對表達量均從眼原基形成期到幼體孵化期逐漸顯著增加相關。頭足類動物中,pax6基因調節基因轉錄控制網絡上游[36],同時,有研究表明,在哺乳動物眼睛晶狀體誘導和分化過程中,six3為pax6的下游調控基因,激活了pax6等相關基因的表達[42-43],而在日本無針烏賊胚胎發育中,pax6基因也在較早時期(眼原基形成期至功能器官分化期)相對表達量較高,基因相對表達量在功能分化基本完成后(幼體孵化期)顯著降低。
筆者僅對日本無針烏賊胚胎不同發育階段3個眼部相關基因進行了分析研究,有關不同發育階段眼部組織學特征及不同發育階段眼部不同部位這3個基因的表達有待進一步分析。
4 結 論
根據日本無針烏賊胚胎3個發育時期轉錄組數據,發現eya、pax6基因在3個發育時期均為超低表達量基因,six3基因在3個發育時期為低表達量基因。3個基因在日本無針烏賊胚胎3個發育時期8個階段的相對表達量變化表明其在基因轉錄控制網絡中發揮不同調節作用。日本無針烏賊胚胎眼部發育過程所涉及的基因轉錄控制網絡機制尚未查明,筆者從胚胎3個發育時期轉錄組數據庫中尋找與胚胎生長發育,器官功能分化等相關基因進行分析,并對每一階段的基因變化進行研究探討,為構建基因轉錄控制網絡,了解調控機制提供數據支持,同時也為日本無針烏賊胚胎的人工繁育和增養殖提供有效的數據參照。
