不同氣候條件下南海西沙海域鳶烏賊(Sthenoteuthis oualaniensis)漁業(yè)生物學(xué)比較研究*
2021-07-21 12:02:56陸化杰張羽翔陳子越陳新軍
海洋與湖沼 2021年4期
陸化杰 寧 欣 劉 維 張羽翔 陳子越 陳新軍, 3, 4, 5, 6
不同氣候條件下南海西沙海域鳶烏賊()漁業(yè)生物學(xué)比較研究*
陸化杰1寧 欣1劉 維2張羽翔2陳子越1陳新軍1, 3, 4, 5, 6
(1. 上海海洋大學(xué)海洋科學(xué)學(xué)院 上海 201306; 2. 海南省海洋與漁業(yè)科學(xué)研究院 海口 570100; 3. 國(guó)家遠(yuǎn)洋漁業(yè)工程技術(shù)研究中心 上海 201306; 4. 大洋漁業(yè)資源可持續(xù)開發(fā)省部共建教育部重點(diǎn)實(shí)驗(yàn)室 上海 201306; 5. 農(nóng)業(yè)農(nóng)村部大洋漁業(yè)開發(fā)重點(diǎn)實(shí)驗(yàn)室 上海 201306; 6. 農(nóng)業(yè)農(nóng)村部大洋漁業(yè)資源環(huán)境科學(xué)觀測(cè)實(shí)驗(yàn)站 上海 201306)
為探討不同氣候條件對(duì)鳶烏賊()漁業(yè)生物學(xué)影響, 根據(jù)2015年8月(厄爾尼諾年)、2016年8月(拉尼娜發(fā)生期)和2017年8月(正常時(shí)期)我國(guó)燈光罩網(wǎng)船在南海采集的1 970尾鳶烏賊樣本, 對(duì)其胴長(zhǎng)、體重、性腺成熟度、胃飽滿度、初次性成熟胴長(zhǎng)進(jìn)行了比較研究。結(jié)果表明: 不同年間鳶烏賊雌雄比例不同, 2017年最高, 2015年次之, 2016年最低; 優(yōu)勢(shì)胴長(zhǎng)組不同, 2015年最大, 2017年次之, 2016年最小; 優(yōu)勢(shì)體重組也不同, 2015年最大, 2017年次之, 2016年最小。協(xié)方差分析表明不同年間胴長(zhǎng)與體重的關(guān)系均不存在性別間顯著性差異, 2015、2016和2017年分別最適用線性、冪和指數(shù)函數(shù)表示; 不同年間樣本性成熟組成存在差異, 雌雄樣本性成熟樣本分別占對(duì)應(yīng)總數(shù)的30.96%和25.40%、46.41%和11.15%、7.35%和28.46%; 胃飽滿度組成也不同。不同年間初次性成熟胴長(zhǎng)不同, 2015、2016和2017年雄性初次性成熟胴長(zhǎng)分別為184.12、144.41、132.70 mm; 雌性分別為192.66、143.68、188.52 mm。結(jié)果表明, 不同氣候變化引起的海洋環(huán)境的變化對(duì)鳶烏賊漁業(yè)生物學(xué)產(chǎn)生影響: 溫度升高導(dǎo)致鳶烏賊個(gè)體減小, 性成熟胴長(zhǎng)減小, 雌雄間的個(gè)體差異變大。
鳶烏賊; 漁業(yè)生物學(xué)特性; 性成熟; 厄爾尼諾; 拉尼娜; 南海
鳶烏賊()為暖水性大洋頭足類, 隸屬頭足綱(Cephalopoda)、槍形目(Idiosepiida)、柔魚科(Ommastrephidae)、鳶烏賊屬() (王堯耕等, 2005), 主要分布于東太平洋、印度洋西北部及南海(張鵬等, 2010)。鳶烏賊生命周期較短, 一般為一年生, 生長(zhǎng)速度快, 繁殖力強(qiáng)(粟麗等, 2016), 具有垂直遷移的特性, 白天位于中下層海域, 夜間上浮捕食, 分布于海面表層至1000 m水層(陳新軍等, 2009), 是我國(guó)頭足類漁業(yè)重要的捕撈對(duì)象, 具有很高的開發(fā)潛力(張宇美等, 2013)。近年來, 極端氣候頻發(fā), 引起海洋氣候環(huán)境復(fù)雜多變(Turner, 2020)。厄爾尼諾與拉尼娜事件為中東太平洋赤道海域溫度大范圍升高和降低現(xiàn)象, 是全球性年際氣候變率的主導(dǎo)模式, 對(duì)太平洋、印度洋及周圍大陸等大范圍的地區(qū)有重要影響(朱益民等, 2007)。鳶烏賊資源補(bǔ)充量大, 易受海洋環(huán)境變化影響(陳新軍等, 2006)。目前有學(xué)者對(duì)鳶烏賊漁業(yè)生物學(xué)受海洋環(huán)境的變化影響有初步的探討(陸化杰等, 2018; 江艷娥等, 2019), 但針對(duì)厄爾尼諾、拉尼娜以及正常氣候條件下鳶烏賊生物學(xué)的對(duì)比研究尚未見報(bào)道。鑒于此, 本研究根據(jù)不同氣候年間采集的1 970尾鳶烏賊樣本, 對(duì)比了其漁業(yè)生物學(xué)特性, 并結(jié)合海洋環(huán)境變化, 分析了極端氣候變化對(duì)南海鳶烏賊的影響, 為后續(xù)鳶烏賊的資源預(yù)測(cè)及可持續(xù)性開發(fā)提供參考。
1 材料與方法
1.1 調(diào)查時(shí)間與海域
采樣海域位于南海西沙群島海域, 采樣時(shí)間為2015年、2016年和2017年的8月1—31日。調(diào)查海域?yàn)?5°45N—19°16′N、109°21′—113°15′E (圖1)。每天從漁獲物中隨機(jī)抽取鳶烏賊樣本20—30尾, 記錄站點(diǎn)并冷凍保存, 不同年間采樣數(shù)量見表1。
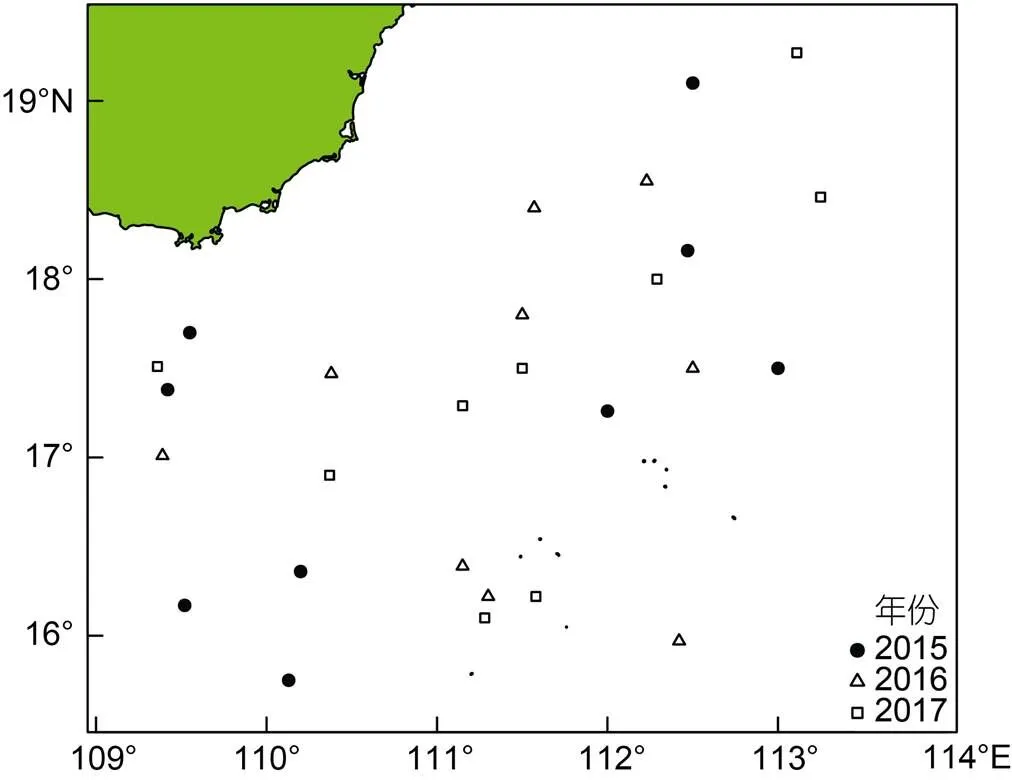
圖1 鳶烏賊采樣分布圖
調(diào)查漁船為海南省三亞榆豐漁業(yè)公司所屬的燈光罩網(wǎng)漁船“瓊?cè)齺?2057”和“瓊?cè)齺?2068”, 兩船為同一船型: 總長(zhǎng)33 m, 型寬6.2 m, 總噸位430 t, 主機(jī)功率237 kW, 副機(jī)功率163 kW, 發(fā)電機(jī)功率200 kW, 平均航速9 kn。
1.2 樣本生物學(xué)測(cè)定
樣本經(jīng)實(shí)驗(yàn)室解凍后進(jìn)行生物學(xué)測(cè)量, 胴長(zhǎng)(mantle length, ML)精確至1 mm、體重(body weight, BW)精確至1 g, 鑒定其性別(sex)、性腺成熟度(maturity stage)和胃飽滿度(stomach stage), 其中性腺成熟度分為Ⅰ—Ⅴ期, Ⅰ、Ⅱ期為性未成熟期, Ⅲ、Ⅳ、Ⅴ期為性成熟期(李建華等, 2016), 胃飽滿度分為1—5級(jí)(陸化杰等, 2014)。
1.3 極端氣候條件確立
依據(jù)美國(guó)國(guó)家海洋和大氣管理局定義, Nino 3.4區(qū)海表溫度距平指數(shù)連續(xù)3個(gè)月高于0.5 °C為一次厄爾尼諾事件, 連續(xù)3個(gè)月低于0.5 °C為一次拉尼娜事件(Chen, 2007b)。
2015年為罕見的厄爾尼諾年, 全年海表面溫度距平(sea surface temperature anomaly, SSTA)均高于0.5 °C。2016年7月至年末SSTA均低于0.5 °C, 為拉尼娜事件。2017年在捕撈月之前均為正常月份, 未發(fā)生厄爾尼諾事件或拉尼娜事件。
1.4 分析方法
(1) 對(duì)不同年份樣本的性別比例、胴長(zhǎng)和體重組成、胃飽滿度和性腺成熟度分布進(jìn)行頻度分析(粟麗等, 2016; 陸化杰等, 2018)。
(2) 采用不同函數(shù)模型擬合ML與BW的生長(zhǎng)方程, 最大似然法估計(jì)模型參數(shù), 赤池信息準(zhǔn)則(AIC, Akaike’s information criterion)選取最低值方程為最優(yōu)模型(Imai, 2002)。
(3) 采用Logistic曲線, 推算不同年間南海鳶烏賊的初次性成熟胴長(zhǎng)(Tafur, 2001; 李建華等, 2016)。
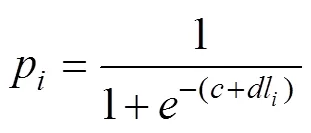
式中,p為組內(nèi)性成熟樣本比例;l為各組胴長(zhǎng)(mm); 初次性成熟胴長(zhǎng)ML50%=–。
2 結(jié)果
2.1 性別比例
研究表明, 不同年間鳶烏賊性別比例不同, 2017年最高, 2015年次之, 2016年最低(詳見表2)。
2.2 胴長(zhǎng)組成
不同氣候年間雌雄胴長(zhǎng)組成均不相同, 其中2015年雄性胴長(zhǎng)范圍84—250 mm, 優(yōu)勢(shì)胴長(zhǎng)組100—160 mm, 占比達(dá)到83.92%; 雌性胴長(zhǎng)范圍99—283 mm, 優(yōu)勢(shì)胴長(zhǎng)組100—160 mm, 達(dá)到78.33% (圖2a)。2016年雄性胴長(zhǎng)范圍56.5—236.5 mm, 優(yōu)勢(shì)胴長(zhǎng)組60—120 mm, 占比達(dá)到80.40%; 雌性胴長(zhǎng)范圍59.5—225 mm, 優(yōu)勢(shì)胴長(zhǎng)組60—140 mm, 達(dá)到75.97% (圖2b)。2017年雄性胴長(zhǎng)范圍59—154 mm, 優(yōu)勢(shì)胴長(zhǎng)組100—140 mm, 占比達(dá)到75.29%; 雌性胴長(zhǎng)范圍71—229 mm, 優(yōu)勢(shì)胴長(zhǎng)組80—140 mm, 達(dá)82.19% (圖2c)。
表1 鳶烏賊樣本采集

Tab.1 Sample collection of S. oualaniensis
表2 鳶烏賊性別比例

Tab.2 Female to male ratio of S. oualaniensis
2.3 體重組成
不同氣候年間雌雄樣本體重組成均不相同。其中2015年雄性體重范圍18—791 g, 優(yōu)勢(shì)體重組30—150 g, 占比達(dá)到77.60%; 雌性體重范圍21—941 g, 優(yōu)勢(shì)體重組30—180 g, 達(dá)到77.92% (圖3a)。2016年雄性體重范圍7—206 g, 優(yōu)勢(shì)體重組0—90 g, 占比達(dá)到77.02%; 雌性體重范圍7—387 g, 優(yōu)勢(shì)體重組0—90 g, 達(dá)到75.27% (圖3b)。2017年雄性體重范圍14—188g, 優(yōu)勢(shì)體重組30—90 g, 占比達(dá)到76.17%; 雌性體重范圍13—584 g, 優(yōu)勢(shì)體重組0—90 g, 達(dá)到77.82% (圖3c)。
2.4 胴長(zhǎng)與體重關(guān)系
協(xié)方差分析表明, 不同氣候年間, 樣本的胴長(zhǎng)和體重的生長(zhǎng)均不存在性別間顯著性差異(2015年:=2.150,=0.143>0.05; 2016年:=0.02,=0.881> 0.05; 2017年:=2.232,=0.136>0.05)。通過模型優(yōu)化比較(表3), 線性函數(shù)為2015年(厄爾尼諾年)南海鳶烏賊胴長(zhǎng)和體重關(guān)系最適表達(dá)模型, 冪函數(shù)為2016年(拉尼娜發(fā)生期)最適表達(dá)模型, 指數(shù)函數(shù)為2017年(正常時(shí)期)最適表達(dá)模型(圖4):
胴長(zhǎng)與體重: BW=3.993 3 ML–427.249 (2015年); BW=2×10–4ML2.730 4(2016年); BW=4.511e0.022 8 ML(2017年)。
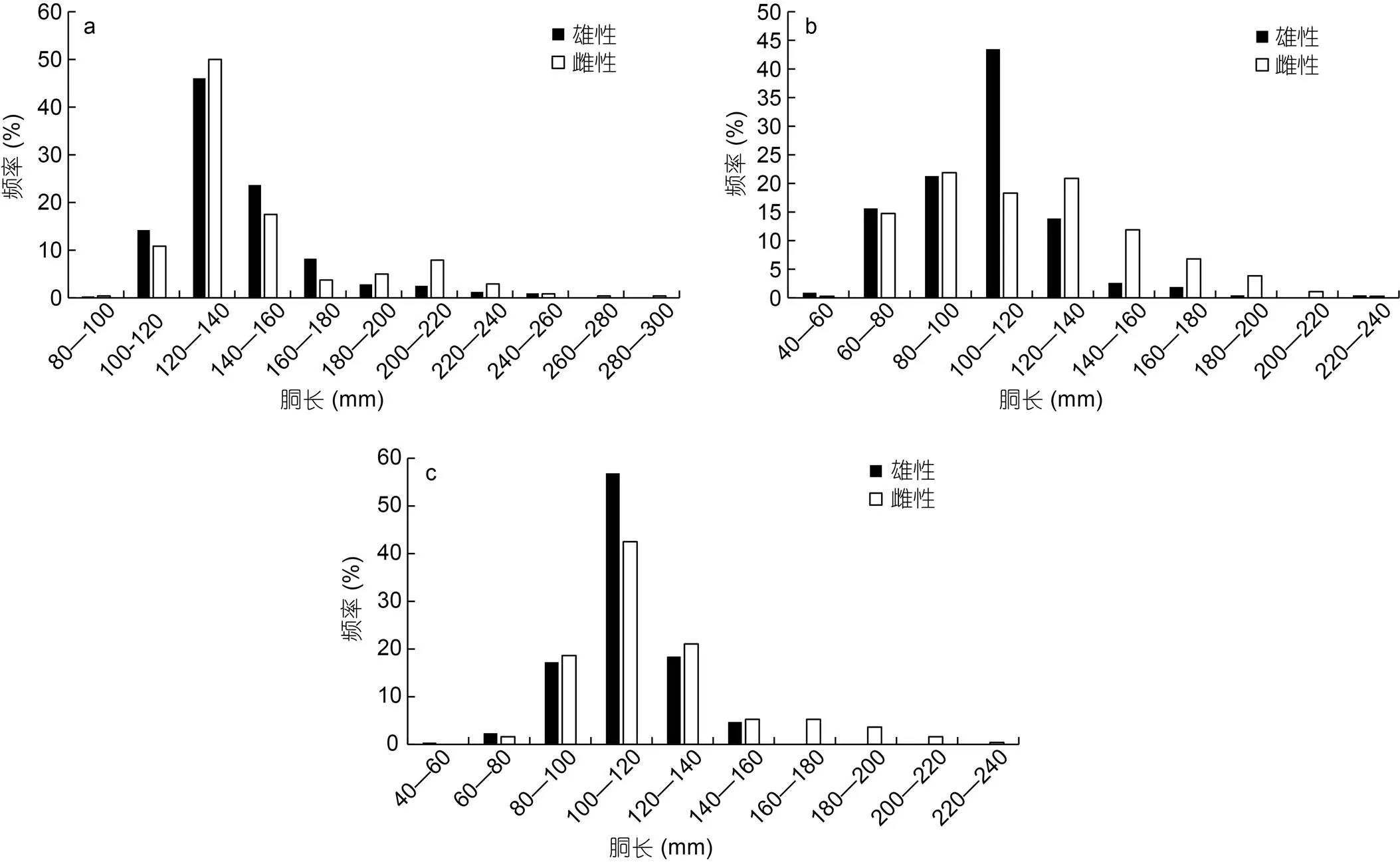
圖2 鳶烏賊胴長(zhǎng)組成
注: a. 2015年胴長(zhǎng)組成分布; b. 2016年胴長(zhǎng)組成分布; c. 2017年胴長(zhǎng)組成分布
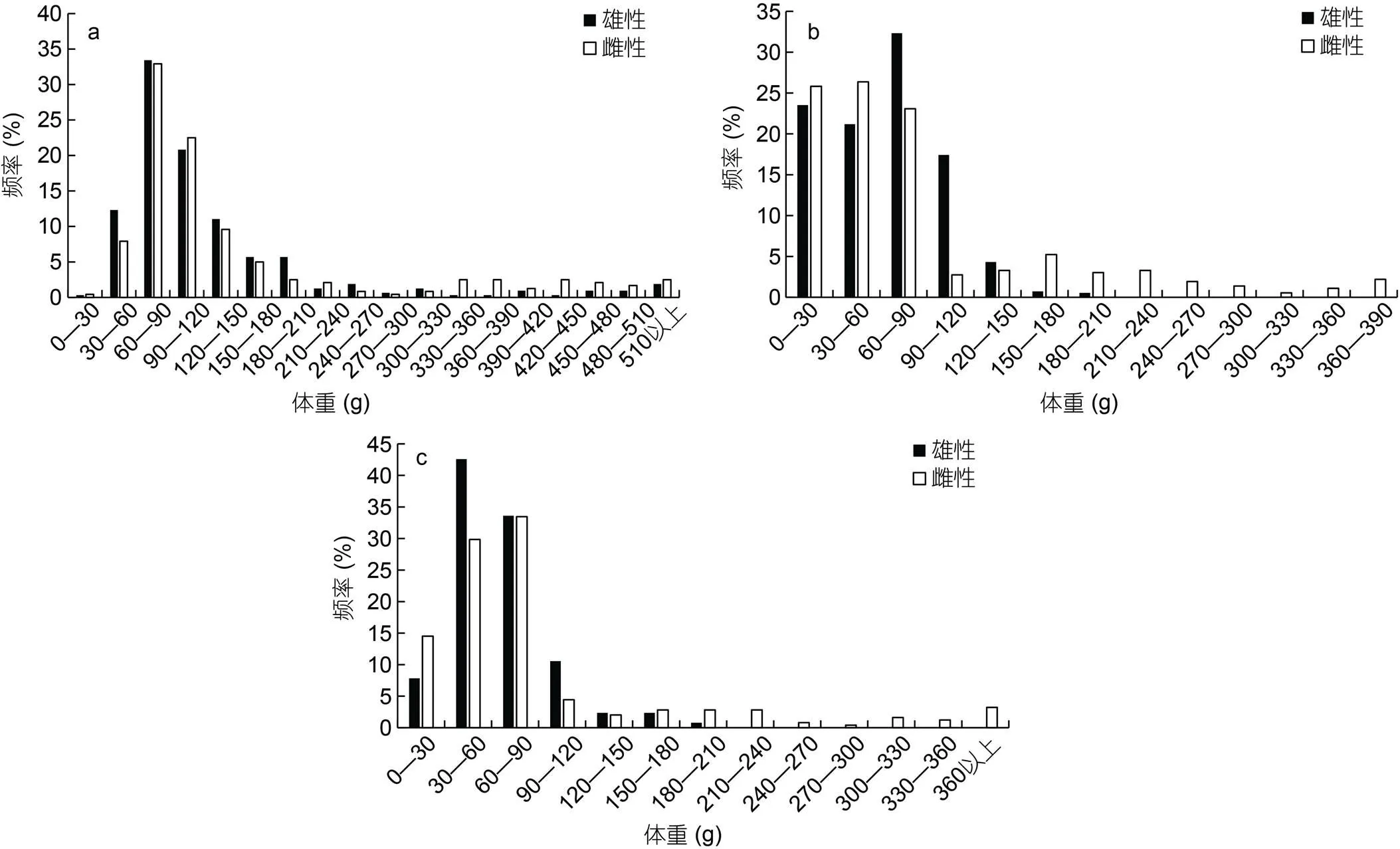
圖3 鳶烏賊體重組成
注: a. 2015年體重組成分布; b. 2016年體重組成分布; c. 2017年體重組成分布
表3 鳶烏賊胴長(zhǎng)與體重生長(zhǎng)模型的生長(zhǎng)參數(shù)與AIC值比較
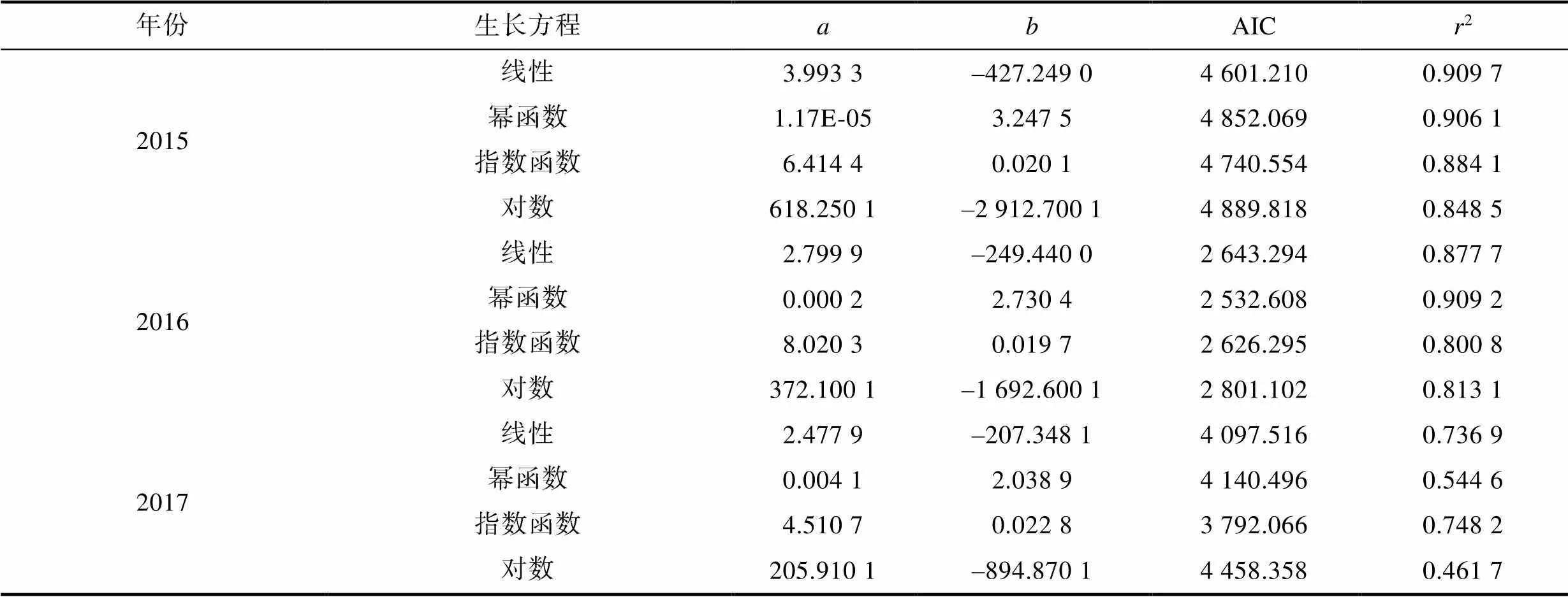
Tab.3 Comparison of parameters and AIC for mental length growth models of S. oualaniensis
2.5 胃飽滿度組成
研究表明, 不同氣候年間鳶烏賊胃飽滿度的組成不同。2015年(厄爾尼諾年)胃飽滿度主要為2—3級(jí), 占總樣本的72.17%, 雌雄胃飽滿度的組成不完全相同: 雄性以2級(jí)最多, 達(dá)到其總數(shù)的45.40%, 其他由高到低依次為3級(jí)、1級(jí)、4級(jí)和5級(jí), 分別占26.98%、20.32%、6.03%和1.27%; 雌性樣本中2級(jí)最多, 達(dá)到其總數(shù)的36.82%, 其他由高到低依次為3級(jí)、1級(jí)、4級(jí)和5級(jí), 分別占35.15%、17.57 %、10.04%和0.42% (圖5a)。2016年(拉尼娜發(fā)生期) 1級(jí)胃飽滿度占總樣本的85.98%, 雄性絕大多數(shù)為1級(jí), 占雄性總數(shù)的93.33%, 2級(jí)和3級(jí)分別為5.83%和0.83%, 無4級(jí)及以上的樣本; 雌性中也以1級(jí)胃飽滿度最多, 占雌性總數(shù)的78.63%, 其他由高到低依次為2級(jí)、3級(jí)和4級(jí), 占17.56%、3.05 %和0.76%, 無5級(jí)雌性樣本(圖5b)。2017年(正常時(shí)期)胃飽滿度2—4級(jí)為優(yōu)勢(shì)組成, 占樣本總數(shù)的76.52%, 雄性以4級(jí)最多, 占其總數(shù)的33.20%, 其他由高到低依次為3級(jí)、5級(jí)、2級(jí)和1級(jí), 分別占雄性總樣本21.09%、20.70%、17.19%和7.81%; 雌性中同樣以4級(jí)最多, 占雌性總樣本的30.89%, 其他由高到低依次為5級(jí)、3級(jí)、2級(jí)和1級(jí), 占27.64%、19.51 %、11.38%和10.57% (圖5c)。
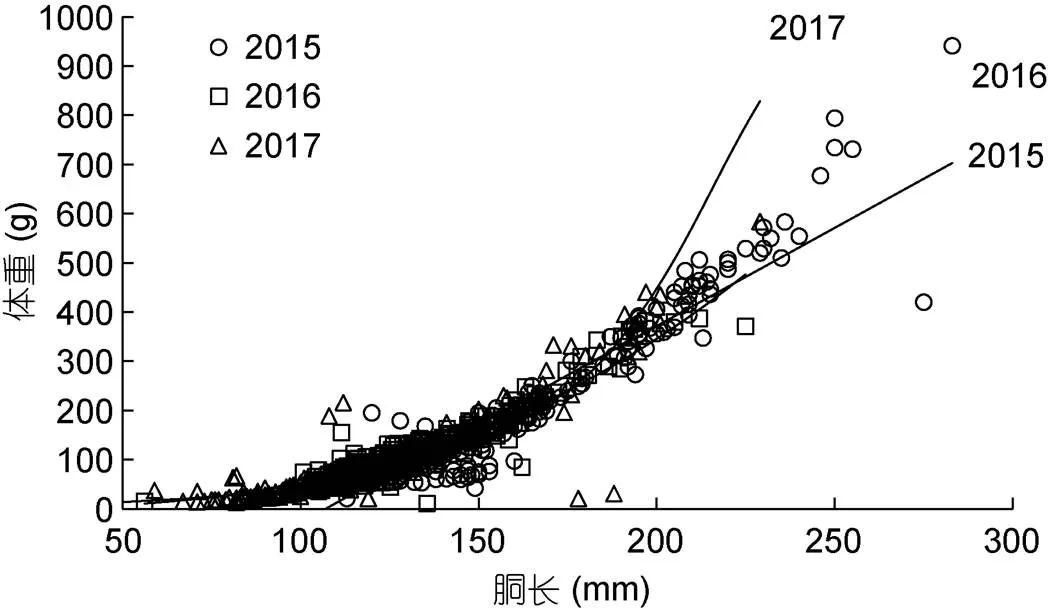
圖4 鳶烏賊胴長(zhǎng)與體重關(guān)系
2.6 性腺成熟度分布
研究表明, 不同氣候年間南海西沙海域鳶烏賊性成熟度分布不同。2015年鳶烏賊雄性個(gè)體Ⅰ期、Ⅱ期、Ⅲ期和Ⅳ期樣本數(shù)依次為149、86、68和12尾, 分別占雄性樣本的47.30%、27.31%、21.59%和3.81%; 雌性鳶烏賊個(gè)體Ⅰ期、Ⅱ期、Ⅲ期和Ⅳ期樣本數(shù)依次為115、50、52和22尾, 分別占雌性樣本的48.12%、20.92%、21.76%和9.20%。兩個(gè)性別均未出現(xiàn)Ⅴ期樣本, 性成熟樣本數(shù)(Ⅲ期以上)分別占雄性和雌性樣本總數(shù)的25.40%和30.96% (圖6a)。2016年雄性Ⅰ期、Ⅱ期、Ⅲ期和Ⅳ期樣本數(shù)依次為292、202、60和2尾, 分別占雄性樣本的52.52%、36.33%、10.79%和0.36%;雌性鳶烏賊個(gè)體Ⅰ期、Ⅱ期、Ⅲ期、Ⅳ期和Ⅴ期樣本數(shù)依次為136、58、46、61和61尾, 分別占雌性樣本的37.57%、16.02%、12.71%、16.85%和16.85%。其中雄性未出現(xiàn)Ⅴ期樣本, 性成熟樣本數(shù)分別占雄性和雌性樣本總數(shù)的11.15%和46.41% (圖6b)。2017年雄性個(gè)體Ⅰ期、Ⅱ期、Ⅲ期和Ⅳ期樣本數(shù)依次為99、82、52和20尾, 分別占雄性樣本的39.13%、32.41%、20.55%和7.90%; 雌性鳶烏賊個(gè)體Ⅰ期、Ⅱ期、Ⅲ期和Ⅳ期樣本數(shù)依次為220、7、4和14尾, 分別占雌性樣本89.80%、2.86%、1.63%和5.71%。兩個(gè)性別均未出現(xiàn)Ⅴ期樣本, 性成熟樣本數(shù)分別占雄性和雌性樣本總數(shù)的28.46%和7.35% (圖6c)。
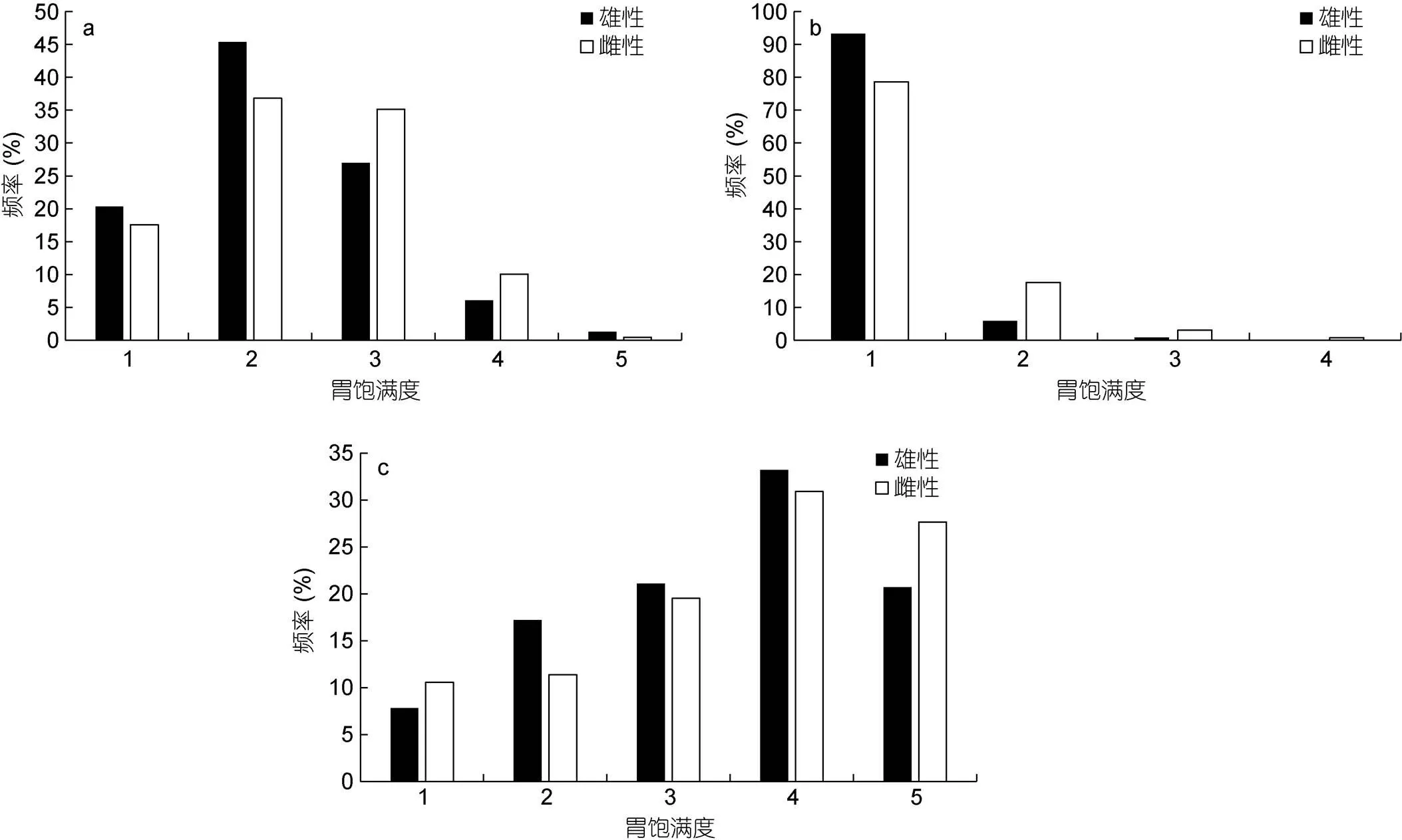
圖5 鳶烏賊胃飽滿度組成
注: a. 2015年胃飽滿度組成; b. 2016年胃飽滿度組成; c. 2017年胃飽滿度組成

圖6 鳶烏賊性腺成熟度組成
注: a. 2015年性腺成熟度組成; b. 2016年性腺成熟度組成; c. 2017年性腺成熟度組成
2.7 初次性成熟胴長(zhǎng)估算
研究表明, 不同氣候年間南海西沙海域鳶烏賊雌雄個(gè)體的初次性成熟胴長(zhǎng)不同: 2015年(厄爾尼諾年)雄性為184.12 mm (圖7a), 雌性為192.66 mm (圖7b); 2016年(拉尼娜發(fā)生期)雄性為144.41 mm (圖7c), 雌性為143.68 mm (圖7d); 2017年(正常時(shí)期)雄性為132.70 mm (圖7e), 雌性為188.52 mm (圖7f)。
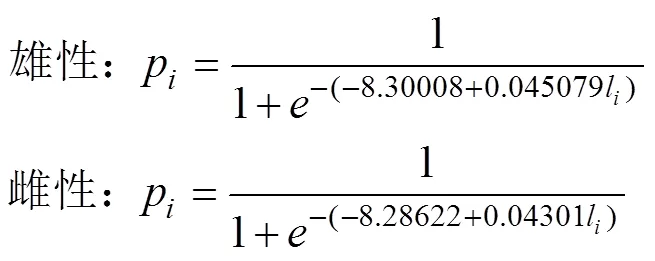
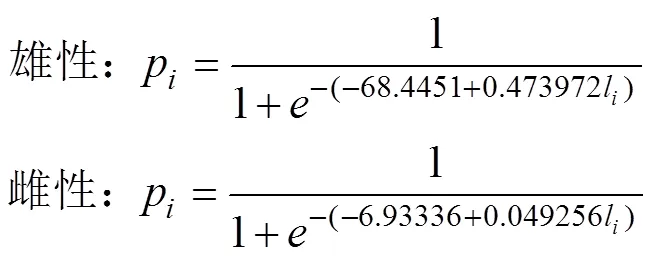

3 討論與分析
3.1 生長(zhǎng)特征
不同氣候條件下雌雄個(gè)體生長(zhǎng)存在差異, 但雌性胴長(zhǎng)與體重均大于雄性, 這與其他學(xué)者的結(jié)果相同(Bello, 2001)。胴長(zhǎng)相同個(gè)體, 雌性觸腕長(zhǎng)于雄性, 鰭寬也更寬, 因此一般認(rèn)為雌性比雄性捕食能力強(qiáng), 攝食量更大(Chembian, 2013), 本研究中不同年間雌雄胃飽滿度較高(3級(jí)以上)的個(gè)體占比分別為45.61%、34.29% (2015年); 3.82%、0.83% (2016年); 78.05%、75% (2017年), 雌性均高于雄性, 也印證這一觀點(diǎn), 即高效的營(yíng)養(yǎng)獲取使雌性生長(zhǎng)速度更快。
不同氣候年間胴長(zhǎng)與體重組成存在差異, 2015年間(厄爾尼諾)鳶烏賊優(yōu)勢(shì)胴長(zhǎng)組最大, 2017年(正常時(shí)期)次之, 2016年(拉尼娜)最小。分析認(rèn)為, 厄爾尼諾期間南海西沙海域鳶烏賊索餌場(chǎng)溫度偏低, 拉尼娜期間溫度偏高(陸化杰等, 2018), 溫度降低, 鳶烏賊個(gè)體增大, 這與其他學(xué)者研究結(jié)論一致, 如加利福尼亞灣莖柔魚()在2009—2010年厄爾尼諾期間個(gè)體偏小(Hoving, 2013); Forsythe等(1989)發(fā)現(xiàn)福氏槍烏賊()在水溫相差1 °C的水域中飼養(yǎng), 個(gè)體大小可以相差三倍; 柔魚()處在生產(chǎn)力水平較高亞熱帶鋒的秋季產(chǎn)卵群個(gè)體明顯大于冬春群個(gè)體(Ichii, 2011)。另有學(xué)者研究發(fā)現(xiàn), 頭足類成體個(gè)體大小受卵粒大小影響, 兩者呈正相關(guān)(Segawa, 1990), 雌性鳶烏賊9—10個(gè)月左右成熟, 多次產(chǎn)卵且產(chǎn)卵期長(zhǎng)(可持續(xù)4個(gè)月), 期間需大量攝食保證產(chǎn)卵, 因此鳶烏賊繁殖行為易受環(huán)境影響, 進(jìn)而影響性成熟個(gè)體大小, 也為本結(jié)果提供了佐證(German, 2002; Laptikhovsky, 2005)。
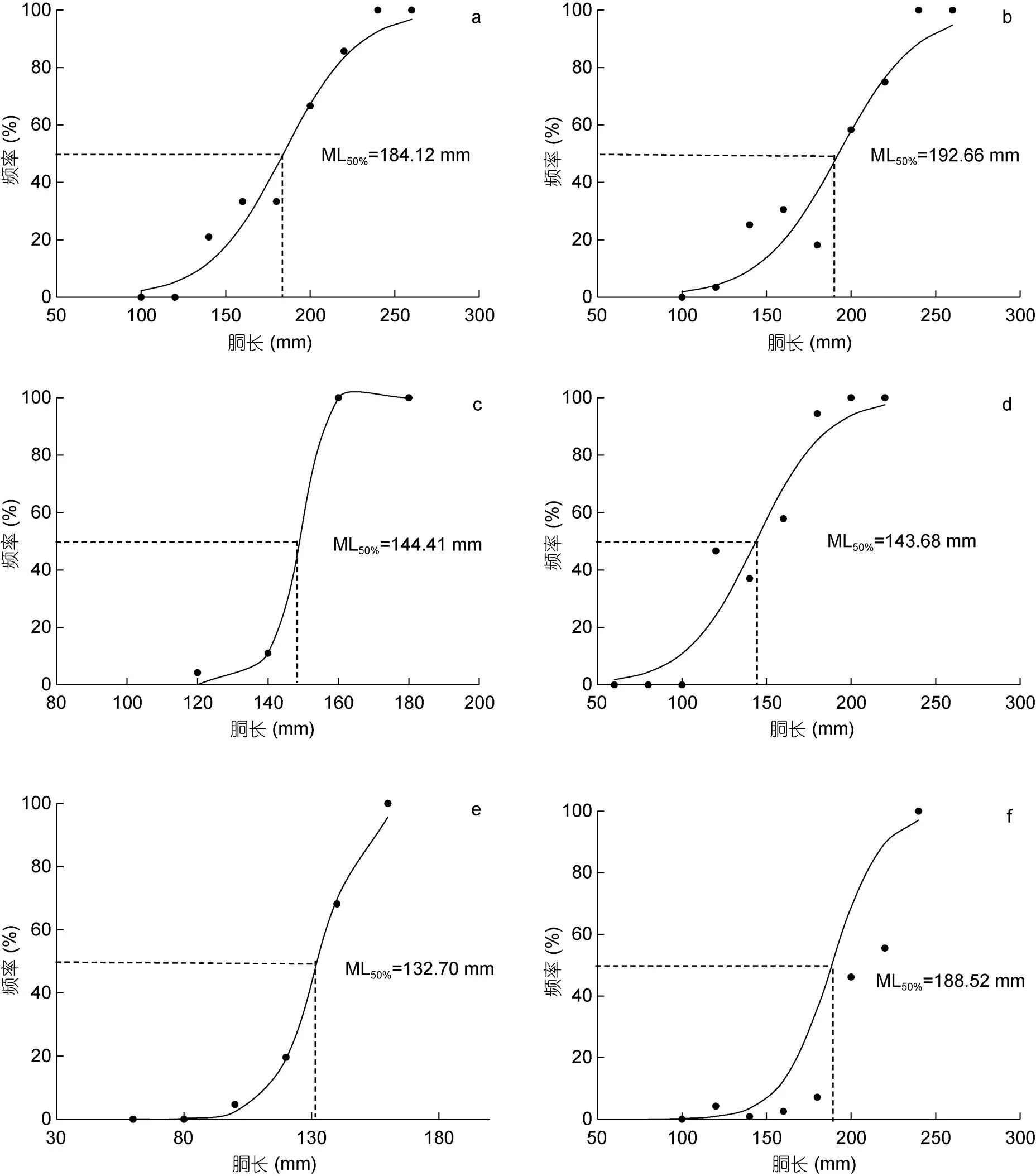
圖7 鳶烏賊初次性成熟胴長(zhǎng)
注: a. 2015年雄性初次性成熟胴長(zhǎng); b. 2015年雌性初次性成熟胴長(zhǎng); c. 2016年雄性初次性成熟胴長(zhǎng); d. 2016年雌性初次性成熟胴長(zhǎng) e. 2017年雄性初次性成熟胴長(zhǎng); f. 2017年雌性初次性成熟胴長(zhǎng)
不同氣候年間胴長(zhǎng)與體重關(guān)系也不相同, 2015年(厄爾尼諾)、2016年(拉尼娜)和2017年(正常時(shí)期)南海西沙鳶烏賊胴長(zhǎng)與體重關(guān)系分別最適用線性函數(shù)、冪函數(shù)和指數(shù)函數(shù)表示, 且三年均不存在性別間顯著性差異。江艷娥等(2019)對(duì)2012年(拉尼娜)南海鳶烏賊研究發(fā)現(xiàn)雌雄個(gè)體之間不存在性別間顯著性差異, 胴長(zhǎng)體重關(guān)系最適合用冪函數(shù)表示, 這與本研究結(jié)論相同。陸化杰等(2018)、李建華等(2016)對(duì)2016年、2014年(正常時(shí)期)南海群島海域中沙、南沙鳶烏賊研究發(fā)現(xiàn)其胴長(zhǎng)與體重關(guān)系存在性別間顯著性差異, 且胴長(zhǎng)與體重關(guān)系最適用冪函數(shù)表示; Chen等(2007a)對(duì)2004年(厄爾尼諾)西北印度洋鳶烏賊的研究認(rèn)為胴長(zhǎng)與體重關(guān)系最適用冪函數(shù)表示, 這與本研究結(jié)果不同。不同海域生長(zhǎng)方程不同, 可能是捕撈海域不同, 受索餌對(duì)象和海洋環(huán)境因素影響導(dǎo)致。
3.2 繁殖特征
不同氣候條件下南海西沙群島鳶烏賊雌雄組成存在差異, 2017年(正常時(shí)期)雌雄比接近于1, 這與李建華等(2016)對(duì)2014年(正常時(shí)期)中西太平洋鳶烏賊研究結(jié)果相同, 但在2015年(厄爾尼諾)、2016年(拉尼娜)雄性個(gè)體數(shù)量均多于雌性。初步分析認(rèn)為, 雌性鳶烏賊承擔(dān)產(chǎn)卵的任務(wù), 性成熟時(shí)性腺大且重。有研究發(fā)現(xiàn), 在不考慮溫度和營(yíng)養(yǎng)等外界因素條件下, 性腺重量與胴體肌肉重量呈反比(Gabr, 1999)。相關(guān)研究還表明, 雌性產(chǎn)卵時(shí)會(huì)將胴體肌肉作為能量?jī)?chǔ)備, 導(dǎo)致雌雄個(gè)體在生殖方面的投入不同, 而溫度變化一般不會(huì)導(dǎo)致雄性頭足類精子活度變化(Naud, 2006), 但會(huì)使雌性頭足類改變卵粒的大小與產(chǎn)卵次數(shù)(Pecl, 2008), 這一現(xiàn)象也可能會(huì)加劇雌雄數(shù)量差異性, 雌性鳶烏賊生長(zhǎng)過程中更高的生殖投入可能導(dǎo)致雌性個(gè)體數(shù)量較少。
本研究表明, 2015年、2017年8月捕撈鳶烏賊性成熟個(gè)體占比存在顯著差異, 2015年性成熟鳶烏賊數(shù)量(27.79%)多于2016年(25.05%)和2017年(18.07%), 且2015年雌、雄鳶烏賊性成熟個(gè)體占比(30.96%、25.40%)差異較2016年(41.41%、11.15%)和2017年(7.35%、28.46%)小, 原因可能是厄爾尼諾期間, 南海溫度偏低(劉成程等, 2017), 造成性腺投入量少(Pecl, 2004), 生長(zhǎng)相同重量性腺對(duì)營(yíng)養(yǎng)物質(zhì)與氧氣等養(yǎng)料需求減少, 性成熟較往年提前, Hoving等(2013)對(duì)莖柔魚的研究中也發(fā)現(xiàn)性成熟提前的現(xiàn)象。
不同氣候條件鳶烏賊初次性成熟胴長(zhǎng)存在差異, 正常時(shí)期(雄性132.70 mm, 雌性188.52 mm)雌雄間初次性成熟胴長(zhǎng)差異較大, 可能是正常時(shí)期較厄爾尼諾期間海水表面溫度偏高, 溫度升高使雌雄間個(gè)體大小差異更大, Pecl(2004)等學(xué)者對(duì)澳大利亞擬烏賊()的研究與本結(jié)論一致, 厄爾尼諾期年間初次性成熟胴長(zhǎng)(雄性184.12 mm, 雌性192.66 mm)明顯長(zhǎng)于拉尼娜年間(雄性144.41 mm, 雌性143.68 mm), 聯(lián)系胃飽滿度分布探究, 2015年全部樣本胃飽滿度以2—3級(jí)為主, 占樣本總數(shù)的72.17%, 而2016年鳶烏賊樣本中胃飽滿度1級(jí)占優(yōu)勢(shì)地位, 達(dá)樣本總數(shù)的85.98%, 可能因?yàn)槔崮绕陂g, 南海鳶烏賊索餌場(chǎng)受海表面信風(fēng)與海流影響, 海水上層水體營(yíng)養(yǎng)不足, 導(dǎo)致鳶烏賊初次性成熟胴長(zhǎng)明顯下降, 營(yíng)養(yǎng)物質(zhì)的豐富程度可能是比溫度更重要的影響因素。
4 結(jié)論與展望
本研究通過厄爾尼諾(2015年)、拉尼娜(2016年)、正常時(shí)期(2017年)鳶烏賊漁業(yè)生物學(xué)資料對(duì)不同海洋環(huán)境下鳶烏賊的個(gè)體差異進(jìn)行了探究, 發(fā)現(xiàn)其對(duì)海洋環(huán)境變化響應(yīng)劇烈, 漁業(yè)生物學(xué)特性差異明顯: 2015年南海鳶烏賊雄性數(shù)量多于雌性, 整體樣本個(gè)體較大, 性成熟時(shí)間提前; 2016年雄性數(shù)量也多于雌性且雌雄性別比例差異最大, 海域營(yíng)養(yǎng)水平較低, 初次性成熟胴長(zhǎng)小; 2017年雌雄數(shù)量較為接近且雌雄間初次性成熟胴長(zhǎng)差異大。鳶烏賊生長(zhǎng)速度快, 捕撈量大, 在南海漁業(yè)中處于重要地位(張宇美等, 2013), 今后應(yīng)對(duì)海水表面溫度、海水表面鹽度、海水表面風(fēng)場(chǎng)、葉綠素濃度等海洋環(huán)境因子如何具體影響鳶烏賊生長(zhǎng)進(jìn)行更深層的研究, 從而為鳶烏賊的開發(fā)提供更充分、更充實(shí)的依據(jù)。
王堯耕, 陳新軍, 2005. 世界大洋性經(jīng)濟(jì)柔魚類資源及其漁業(yè). 北京: 海洋出版社, 78—93
劉成程, 張文超, 晏 宏, 2017. 南海西沙海域溫度異常對(duì)厄爾尼諾-南方濤動(dòng)活動(dòng)的響應(yīng). 地球環(huán)境學(xué)報(bào), 8(2): 148—156
李建華, 陳新軍, 方 舟等, 2016. 3個(gè)不同海區(qū)鳶烏賊漁業(yè)生物學(xué)的初步比較. 海洋漁業(yè), 38(6): 561—569
朱益民, 楊修群, 陳曉穎等, 2007. ENSO與中國(guó)夏季年際氣候異常關(guān)系的年代際變化. 熱帶氣象學(xué)報(bào), 23(2): 105—116
江艷娥, 陳作志, 林昭進(jìn)等, 2019. 南海海域鳶烏賊中型群與微型群漁業(yè)生物學(xué)比較. 水產(chǎn)學(xué)報(bào), 43(2): 454—466
張 鵬, 楊 吝, 張旭豐等, 2010. 南海金槍魚和鳶烏賊資源開發(fā)現(xiàn)狀及前景. 南方水產(chǎn), 6(1): 68—74
張宇美, 顏云榕, 盧伙勝等, 2013. 西沙群島海域鳶烏賊攝食與繁殖生物學(xué)初步研究. 廣東海洋大學(xué)學(xué)報(bào), 33(3): 56—64
陸化杰, 王從軍, 陳新軍, 2014. 4—6月東太平洋赤道公海鳶烏賊生物學(xué)特性初步研究. 上海海洋大學(xué)學(xué)報(bào), 23(3): 441—447
陸化杰, 童玉和, 劉 維等, 2018. 厄爾尼諾年春季中國(guó)南海中沙群島海域鳶烏賊的漁業(yè)生物學(xué)特性. 水產(chǎn)學(xué)報(bào), 42(6): 912—921
陳新軍, 劉必林, 王堯耕, 2009. 世界頭足類. 北京: 海洋出版社, 1065—1066
陳新軍, 邵 峰, 2006. 印度洋西北部公海鳶烏賊資源特征及其與海況的關(guān)系. 中國(guó)海洋大學(xué)學(xué)報(bào), 36(4): 611—616
粟 麗, 陳作志, 張 鵬, 2016. 南海中南部海域春秋季鳶烏賊繁殖生物學(xué)特征研究. 南方水產(chǎn)科學(xué), 12(4): 96—102
Bello G, 2001. Dimorphic growth in male and female cuttlefish(Cephalopoda: Sepiidae) from the Adriatic Sea. Helgoland Marine Research, 55: 124—127
Chembian A J, 2013. Studies on the Biology, Morphometrics and Biochemical composition of the Ommastrephid squid,(Lesson, 1830) of the south west coast of India. Cochin: Doctor Dissertation of Cochin University of Science and Technology, 46
Chen X J, Liu B L, Tian S Q, 2007a. Fishery biology of purpleback squid,, in the northwest Indian Ocean. Fisheries Research, 83(1): 98—104
Chen X J, Zhao X H, Chen Y, 2007b. Influence ofon the western winter-spring cohort of neon flying squid () in the northwestern Pacific Ocean. ICES Journal of Marine Science, 64(6): 1152—1160
Forsythe J W, Hanlon R T, 1989. Growth of the Eastern Atlantic squid,Steenstrup (Mollusca: Cephalopoda). Aquaculture Research, 20(1): 1—14
Gabr H R, Hanlon R T, Hanafy M H, 1999. Reproductive versus somatic tissue allocation in the cuttlefish Sepia dollfusi Adam (1941). Bulletin of Marine Science, 65(1): 159—173
German Z, Chesalin N, Michael C, 2002. Main results of long-term worldwide studies on tropical nektonic oceanic squid genus: An overview of the soviet investigations. Bulletin of Marine Science, 71(2): 1019—1060
Hoving H J T, Gilly W F, Markaida U, 2013. Extreme plasticity in life-history strategy allows a migratory predator (jumbo squid) to cope with a changing climate. Global Change Biology, 19(7): 2089—2103
Ichii T, Mahapatra K, Sakai M, 2011. Changes in abundance of the neon flying squidin relation to climate change in the central North Pacific Ocean. Marine Ecology Progress Series, 441: 151—164
Imai C, Sakai H, Katsura K, 2002. Growth model for the endangered cyprinid fishbased on otolith analyses. Fisheries Science, 68(4): 843—848
Laptikhovsky V V, Nigmatullin C M, 2005. Aspects of female reproductive biology of the orange-back squid,(Steenstup) (Oegopsina: Ommastrephidae) in the eastern tropical Atlantic. Scientia Marina, 69(3): 383—390
Naud M J, Havenhand J N, 2006. Sperm motility and longevity in the giant cuttlefish,(Mollusca: Cephalopoda). Marine Biology, 148(3): 559—566
Pecl G, 2004. Therelationships between season of hatching, growth and condition in the southern calamary,. Marine and Freshwater Research, 55(4): 429—438
Pecl G, Jackson G D, 2008. The potential impacts of climate change on inshore squid: biology, ecology and fisheries. Reviews in Fish Biology and Fisheries, 18(4): 373—385
Pecl G, Moltschaniwskyj N, Tracey S R, 2004. Inter-annual plasticity of squid life history and population structure: ecological and management implications. Oecologia, 139(4): 515—524
Segawa S, 1990. Food consumption, food conversion and growth rates of the oval squidby laboratory experiments. Nippon Suisan Gakkaishi, 56(2): 217—222
Tafur R, Villegas P, Rabí M, 2001. Dynamics of maturation, seasonality of reproduction and spawning grounds of the jumbo squid(Cephalopoda: Ommastrephidae) in Peruvian waters. Fisheries Research, 54(1): 33—50
Turner R, McConney P, Monnereau I, 2020. Climate change adaptation and extreme weather in the small-scale fisheries of Dominica. Coastal Management, 48(5): 436—455
COMPARISON IN FISHERY BIOLOGY OFIN DIFFERENT CLIMATE EVENTS IN THE SOUTH CHINA SEA
LU Hua-Jie1, NING Xin1, LIU Wei2, ZHANG Yu-Xiang2, CHEN Zi-Yue1, CHEN Xin-Jun1, 3, 4, 5, 6
(1. College of Marine Sciences of Shanghai Ocean University, Shanghai 201306, China; 2. Hainan Academy of Ocean and Fishery Sciences, Haikou 570100, China; 3. National Distant-water Fisheries Engineering Research Center, Shanghai Ocean University, Shanghai 201306, China; 4. The Key Laboratory of Sustainable Exploitation of Oceanic Fisheries Resources, Shanghai Ocean University, Ministry of Education, Shanghai 201306, China; 5. Key Laboratory of Oceanic Fisheries Exploration, Ministry of Agriculture and Rural Affairs, Shanghai 201306, China; 6. Scientific Observing and Experimental Station of Oceanic Fishery Resources, Ministry of Agriculture and Rural Affairs, Shanghai 201306, China)
The effect of different climate events on the fishery biology of the purple back flying squid() was studied based on 1 970 samples collected during El Ni?o year (August 2015), La Ni?a year (August 2016), and normal year (August 2017) in the South China Sea. The result indicated that the ratio of females to males was different in the different years, the largest in 2017, the second in 2015, and the smallest in 2016. The mantle length (ML) and body weight (BW) composition was also different in those years with a different high concentration (the largest in 2015, then in 2017, and smallest in 2016). The relationship between the ML and BW was different and there was no significant difference in gender among those years, and the relationship could be best described by linear (2015), power (2016), and exponent (2017) functions. The matured individuals were different in those years, accounting for 25.40%, 11.15%, and 28.46% for males, and 30.96%, 46.41%, 7.35% for females, respectively. In addition, the feeding stages were different. The first mature mantle length was different in those years, being 184.12 mm, 144.41 mm, and 132.70 mm for males; and 192.66 mm, 143.68 mm, and 188.52 mm for females in 2015, 2016, and 2017, respectively. Warmer seawater could resulted in smaller body size, shorter fist mature ML, and enlarged individual difference between male and female.
; fisheries biological characteristic; gonad maturity stage; the El Ni?o year; the La Ni?a year; the South China Sea
* 國(guó)家重點(diǎn)研發(fā)計(jì)劃項(xiàng)目, 2019YFD0901404號(hào), 2019YFD090402號(hào); 國(guó)家自然科學(xué)青年基金項(xiàng)目, NSFC 4150618號(hào)。陸化杰, 博士, 副教授, 碩導(dǎo), E-mail: hjlu@shou.edu.cn
2020-11-02,
2021-03-25
S931; S932; Q958
10.11693/hyhz20201100304
