8個釀酒葡萄品種的抗寒性比較
2021-07-24 12:57:06賈金輝劉慧純蔡智軍田曉玲張海濤
中國果樹 2021年7期
賈金輝,徐 凌,劉慧純,蔡智軍,田曉玲,張海濤
(遼寧農業職業技術學院,營口115009)
隨著我國釀酒葡萄的發展,遼寧省也開始出現葡萄酒廠,比如本溪桓仁有五女山酒廠,也有張裕的分廠,但葡萄酒業在遼寧省發展較慢,最主要的原因是釀酒葡萄品種大多數從國外引進,抗寒性較差,很容易受到凍害,為了防止凍害,需要進行埋土防寒,增加成本,耗時耗力[1]。所以,對釀酒葡萄品種的抗寒性研究意義重大,本試驗以8 個釀酒葡萄品種作為材料,通過對枝條的相對電導率、芽段萌發率、可溶性糖含量、游離脯氨酸含量、丙二醛含量,根系的過冷能力、半致死溫度、平均隸屬度值等指標進行相關性分析[2],得出釀酒葡萄品種抗寒性的判斷依據,為東北寒冷地區的釀酒葡萄種植與發展提供一定的參考。
1 材料與方法
1.1 試驗材料
試驗材料取自遼寧農業職業技術學院釀酒葡萄園(位于遼寧省營口市熊岳鎮),8 個品種分別為‘雙豐’‘左優紅’‘北紅’‘北醇’‘雙紅’‘北冰紅’‘威代爾’‘公釀一號’。2018 年11 月剪取枝條和根系,每個品種隨機選15 株樹,每株隨機選2~3 根長勢良好的1 年生枝條和根系,枝條長度25~30 cm,根系直徑2~3 mm,純凈水清洗干凈,濾紙吸干枝條和根系的表面水分,將其封入樣本袋內待測[3]。
1.2 試驗方法
將供試枝條一部分置于4 ℃作為對照,其余分別在-10~-35 ℃每隔-5 ℃設置溫度梯度進行冷凍處理,并在達到設定溫度后保持15 h,再升至4 ℃保持1 d。然后隨機取樣進行相關指標的測定,相對電導率采用電導儀測定,芽段萌發率采用比值法測定,可溶性糖含量采用蒽酮法測定,丙二醛含量采用硫代巴比妥酸法測定,脯氨酸含量采用茚三酮比色法測定[4]。
將供試根系放入人工霜凍試驗箱內,并將溫度傳感器一端固定在根系表皮上,另一端與計算機和數據采集系統連接,設置計算機連續自動記錄數據,分析根系表面溫度變化。設置人工霜凍試驗箱從室溫降至-13 ℃,以3 ℃/h 進行降溫。繪制出溫度變化曲線,可確定根系的結冰點和過冷卻點。
根系的過冷能力=過冷卻點溫度-結冰點溫度[5]
1.3 數據處理和分析
試驗數據采用Excel 2016 和SPSS 15.10 軟件進行整理和分析。
2 結果與分析
2.1 枝條在低溫下的變化
2.1.1 枝條的相對電導率在低溫下的變化
從表1 可以看出,隨著溫度的降低8 個品種枝條的相對電導率都增加。-20 ℃時,‘公釀一號’的相對電導率大于40%,且極顯著高于其他品種,由此可知‘公釀一號’的抗寒性最弱[6]。-25 ℃時,‘北冰紅’‘威代爾’的相對電導率均大于50%,均極顯著高于除‘公釀一號’外的其他5 個品種,且其他5 個品種的相對電導率差異不顯著,說明‘北冰紅’‘威代爾’的抗寒性比除‘公釀一號’外的其他5 個品種弱。當溫度降至-30 ℃時,‘北醇’‘北紅’‘雙紅’‘左優紅’的相對電導率均大于50%,且均極顯著高于‘雙豐’,說明‘雙豐’的抗寒性最強[7]。
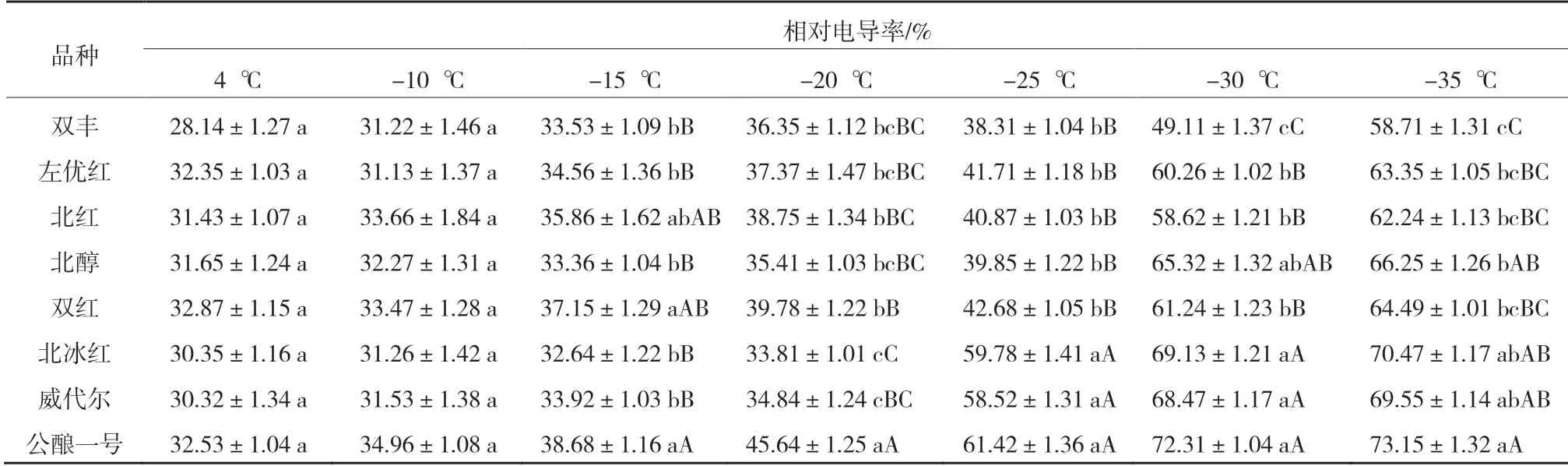
表1 8 個品種枝條在各低溫處理下的相對電導率
2.1.2 枝條的芽段萌發率在低溫下的變化
從表2 可以看出,隨著溫度的降低8 個品種枝條的芽段萌發率都降低,且差異顯著。4 ℃時,8個品種枝條的芽段萌發率都大于90%,且差異不顯著,說明8 個品種的枝條芽段健康良好[8]。當-10 ℃及更低溫度時,‘公釀一號’的芽段萌發率極顯著低于其他品種,故‘公釀一號’的抗寒性最弱。除‘公釀一號’外,‘北冰紅’和‘威代爾’的芽段萌發率均極顯著低于其他品種,且二者間差異不顯著,說明‘北冰紅’和‘威代爾’的抗寒性比‘公釀一號’強,比其他品種弱。在-15 ℃及更低溫度時,‘左優紅’‘北紅’‘北醇’‘雙紅’的芽段萌發率均極顯著低于‘雙豐’,說明‘雙豐’的抗寒性最強。

表2 8 個品種枝條在各低溫處理下的芽段萌發率
2.1.3 枝條的可溶性糖含量在低溫下的變化
由表3 可知,隨著溫度降低8 個品種枝條的可溶性糖含量都先升高后降低。‘公釀一號’在-10~-15 ℃的可溶性糖含量極顯著低于其他品種,但隨溫度降低‘公釀一號’受低溫凍害影響可溶性糖含量突然增加,其抗寒性最差[9]。在-10 ℃及更低溫度(-30 ℃除外)時,‘北醇’‘雙紅’‘北冰紅’‘威代爾’可溶性糖含量極顯著低于除‘公釀一號’外的其他品種,抗寒性居中。在-10~-25 ℃,‘北紅’和‘左優紅’的可溶性糖含量差異不顯著。

表3 8 個品種枝條在各低溫處理下的可溶性糖含量
2.1.4 枝條的游離脯氨酸含量在低溫下的變化
由表4 可知,8 個品種枝條的游離脯氨酸含量除‘雙豐’隨溫度的降低持續升高外其他品種均先升高后降低,說明‘雙豐’抗寒性最強。-25 ℃時‘公釀一號’的游離脯氨酸含量最低,且極顯著低于其他品種,抗寒性最弱。-30 ℃時‘北冰紅’和‘威代爾’的游離脯氨酸含量極顯著低于除‘公釀一號’外的其他品種,說明‘北冰紅’和‘威代爾’的抗寒性較弱。在-10~-25 ℃,‘北紅’‘北醇’ ‘雙紅’的游離脯氨酸含量無顯著差異。

表4 8 個品種枝條在各低溫處理下的游離脯氨酸含量
2.1.5 枝條的丙二醛含量在低溫下的變化
從表5 可以看出,隨著溫度的降低8 個品種枝條的丙二醛含量除‘雙豐’持續升高外其他品種均先升高后降低。‘公釀一號’在-20 ℃時丙二醛含量最高,極顯著高于其他品種,抗寒性最弱[10]。在-25 ℃時‘北冰紅’和‘威代爾’的丙二醛含量較高,極顯著高于其他品種,抗寒性較弱。‘左優紅’‘北紅’‘北醇’‘雙紅’在-30 ℃時丙二醛含量均達到最高值,‘雙豐’則在-35 ℃時丙二醛含量達到最高值。
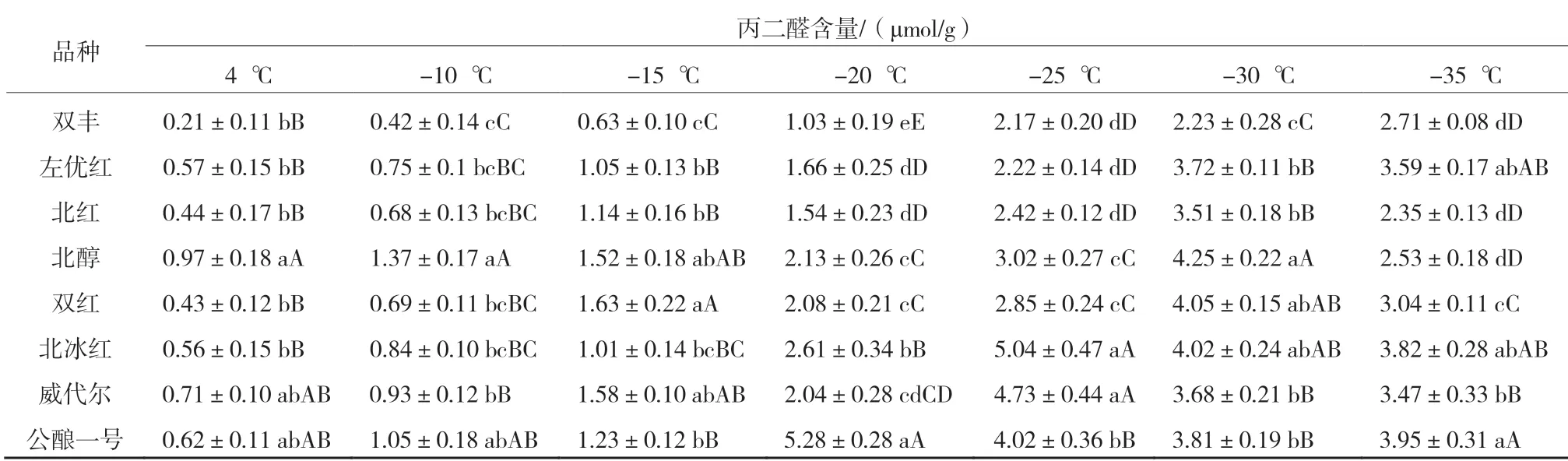
表5 8 個品種枝條在各低溫處理下的丙二醛含量
2.2 根系在低溫下的變化
2.2.1 根系的過冷卻點與結冰點
研究表明葡萄根系組織抗寒性與過冷卻點呈負相關[5]。0 ℃以下時,根系組織細胞為過冷卻的狀態,此狀態下的細胞內水分仍呈液態,不會受到冰凍傷害。所以,抗寒性越強的根系,其過冷卻點就越低。由表6 可知,‘雙豐’根系的過冷卻點最低,抗寒性最強,能在-8.5 ℃中存活,其次是‘北紅’‘左優紅’‘雙紅’‘北醇’根系能在-7.8~-7.1 ℃中存活,‘北冰紅’和‘威代爾’根系則能在-5.9~-5.8 ℃中存活,‘公釀一號’根系的過冷卻點最高,能在-5.1 ℃中存活。

表6 8 個品種根系的過冷卻點與結冰點
2.2.2 根系的過冷能力
葡萄根系組織的過冷能力可用結冰點與過冷卻點溫度差來表示,過冷能力越強,溫度差越大[5]。由圖1 可知,‘雙豐’的過冷能力最高,抗寒性最強;‘公釀一號’的過冷能力最低,抗寒性最弱;其他品種的過冷能力由高到低依次為‘北紅’‘左優紅’‘雙紅’‘北醇’‘威代爾’‘北冰紅’。此抗寒能力強弱排序基本與枝條、葉片得出的結果一致。
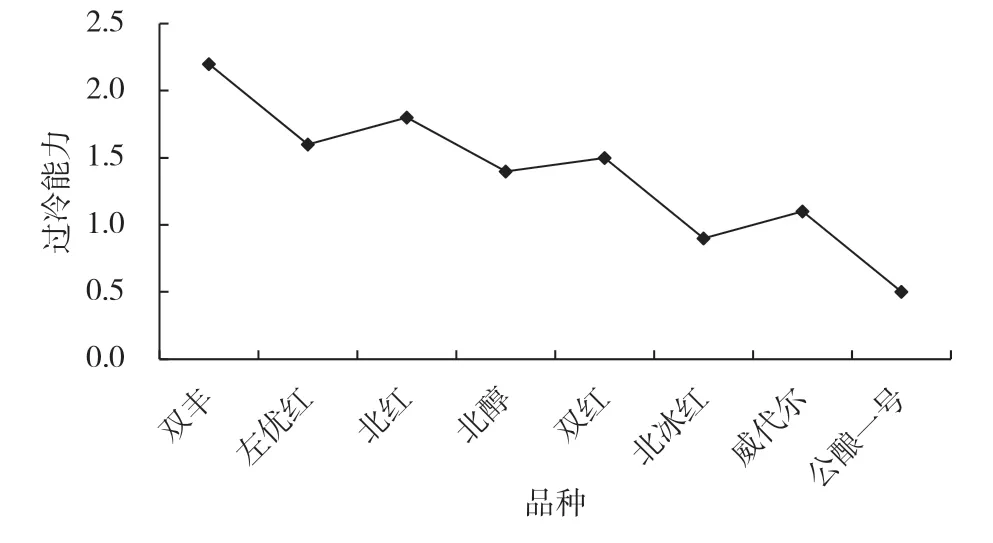
圖1 8 個品種根系的過冷能力
2.3 抗寒性的綜合評價
2.3.1 建立Logistic 方程和求解半致死溫度
研究表明溫度與相對電導率之間的關系為曲線[7],Logistic 方程也是曲線方程,故能用Logistic方程進行擬合。y=K/(1+ae-bx)為Logistic 方程,LT50=-lna/b 為半致死溫度即曲線的拐點[11-12]。
根據8 個葡萄品種枝條在預設溫度下的相對電導率建立Logistic 方程、求解半致死溫度LT50和擬合度(R2),見表7。從表7 可知,‘雙豐’半致死溫度約為-32.9 ℃,極顯著低于其他品種,抗寒性最強。‘左優紅’‘北紅’‘北醇’‘雙紅’4 個品種的半致死溫度在-27.5~-25.4 ℃。‘北冰紅’和‘威代爾’的半致死溫度在-22.0 ℃左右。‘公釀一號’的半致死溫度約為-18.4 ℃,極顯著高于其他品種,抗寒性最弱。

表7 8 個品種的半致死溫度
2.3.2 利用隸屬函數法求解抗寒性強弱
研究表明平均隸屬度與抗寒性有相關性[13],抗寒能力越弱的平均隸屬度越小。隸屬函數方程為X(u)=(X-Xmin)/(Xmax-Xmin)或X(u)=1-[(X-Xmin)/(Xmax-Xmin)]。利用此方程求解8 個品種的抗寒性位次。由表8 可知,8 個品種抗寒性由弱到強排序為‘公釀一號’<‘北冰紅’<‘威代爾’<‘北醇’<‘雙紅’<‘左優紅’<‘北紅’<‘雙豐’。抗寒性結果與半致死溫度一致。

表8 8 個品種的隸屬度值
2.3.3 抗寒各指標的相關性分析
為了研究抗寒性指標間的關聯程度,對8 個品種的枝條、根系各指標、半致死溫度和平均隸屬度值進行相關性分析[14]。由表9 可知,與其他各指標的相關系數均達到極顯著水平的有平均隸屬度值、半致死溫度和根系的過冷能力,而其他各指標之間的相關性較差,因此可以用平均隸屬度值、半致死溫度和根系的過冷能力評價葡萄抗寒性的強弱[15]。

表9 各指標間的相關性分析
3 討論與結論
植物細胞膜的透性與環境溫度緊密相關,溫度較低時,膜透性將隨之改變,功能逐漸降低甚至完全喪失,這會導致細胞內的溶液向外滲出。滲出的溶液越多,電導率就越高。抗寒性和細胞膜的透性存在負相關性,通過測定枝條的電導率來判斷枝條的抗寒性是一種經典的試驗方法[16-18]。Logistic 方程是典型的S 曲線方程,相對電導率與溫度之間的關系曲線能較好地用Logistic 方程進行擬合[19-20]。曲線的拐點即為半致死溫度[21]。半致死溫度可以作為評價抗寒性的一項重要指標[22-23]。研究表明,植物的抗寒能力與枝條的芽段萌發率呈正相關性,植物的抗寒能力越強,其在低溫下的芽段萌發率越高。許多研究認為,植物受到低溫脅迫時,體內的可溶性糖會發生變化,隨之改變細胞的滲透濃度,溫度越低,可溶性糖含量會越高,使滲透濃度增加,導致冰點隨之降低[24-25]。艾琳、王敏等研究表明,在常見氨基酸中游離脯氨酸的溶解度通常比較大,游離脯氨酸又是水溶性良好的氨基酸,所以它可以改變細胞的滲透壓,來保護原生質體[26-27]。當植物受到低溫凍害時,枝條上的游離脯氨酸含量會隨之上升,增大細胞內蛋白質膠體親水面積,增加植物的抗寒能力[28-29]。尤超等研究表明,植物在低溫環境下過多的活性氧容易產生膜脂過氧化,引發過氧化產物丙二醛的大量積累,因此可以通過測定丙二醛含量來判斷植物的抗寒能力[30]。
本試驗通過測定一些抗寒性生理指標,分析各項指標與抗寒性的相關性得知:枝條的芽段萌發率、游離脯氨酸含量、可溶性糖含量以及根系的過冷能力與抗寒性呈正相關;枝條的丙二醛含量和相對電導率與抗寒性呈負相關。枝條的芽段萌發率、相對電導率、可溶性糖含量、丙二醛含量、游離脯氨酸含量與其他抗寒指標的相關性呈顯著水平,平均隸屬度值、半致死溫度和根系的過冷能力與其他抗寒指標的相關性呈極顯著水平,因此可以用后3項指標來評價抗寒性強弱。利用隸屬函數法、半致死溫度和根系的過冷能力比較8 個葡萄品種的抗寒性,抗寒能力由弱到強一致表現為‘公釀一號’<‘北冰紅’<‘威代爾’<‘北醇’<‘雙紅’<‘左優紅’<‘北紅’<‘雙豐’。
