miR-214-3p對缺糖缺氧再灌注誘導PC12細胞凋亡的作用及機制*
2021-08-02 02:56:22郭冬芬任真奎胡玉梅吳昌學禹文峰
貴州醫科大學學報 2021年7期
關鍵詞:血清
郭冬芬, 任真奎, 胡玉梅, 吳昌學*, 禹文峰*
(1.貴州醫科大學 分子生物學重點實驗室, 貴州 貴陽 550004; 2.貴州醫科大學 地方病與少數民族疾病教育部重點實驗室, 貴州 貴陽 550004; 3.黔西南州人民醫院 檢驗科, 貴州 興義 562400)
腦卒中是一種急性腦血管疾病,分為出血性和缺血性腦卒中,其中急性缺血性腦卒中又稱為腦梗死,是由于腦組織局部區域血液供應障礙,導致腦組織缺血、缺氧而發生壞死[1-2]。缺血性腦卒中具有高發病率、高致殘及高死亡率的特點,嚴重威脅人類健康,是世界范圍內致殘和致死的第二大疾病[3-5]。溶栓是目前公認的臨床治療急性缺血性腦卒中的唯一有效的治療方案[6-7],然而靜脈溶栓增加了出血性轉化的風險,反而會造成患者病情惡化和死亡[8-9],因而尋找新的治療靶點是研究者們一直重點關注的方向。micro RNA(miRNA)是小非編碼RNA中的一類,長度為21個核苷酸,能在轉錄后參與基因表達的調控,在細胞信號通路中發揮著重要功能,亦可在許多生理或病理過程中發生表達異常[10]。近年研究發現,miRNA與急性腦卒中發生具有密切關系[11];腦卒中患者中出現miRNA表達異常,如與發育相關的miR-124在大鼠和小鼠腦卒中模型中表達均顯著上調[12-13],miR-210在腦卒中患者血液中表達下調[14]。B淋巴細胞瘤因子2樣蛋白2(B-cell lymphoma 2 like protein 2,BCL2L2)作為BCL2家族新成員之一,具有抗凋亡功能[15],但miR-214-3p是否能通過靶向BCL2L2而參與調控神經元凋亡尚未見相關報道。因此,本研究通過構建大鼠腎上腺髓質嗜鉻瘤細胞缺糖缺氧再灌注模型,檢測miR-214-3p表達的變化及其對細胞凋亡的影響,探討miR-214-3p與腦卒中的相關性。
1 材料與方法
1.1 實驗材料
1.1.1細胞來源 腎上腺嗜鉻神經細胞瘤12(adrenal pheochromocytoma 12,PC12)細胞、人胚腎細胞293T購于中國科學院上海細胞庫。
1.1.2主要試劑與儀器miR-214-3p逆轉錄引物、模擬物(miR-214-3pmimic)、抑制物(miR-214-3pinhibitor)及相對應的模擬物對照(miR-214-3p negative control,miR-214-3pNC,廣州銳博),實時熒光定量聚合酶鏈式反應(quantitative real-time polymerase chain reaction,real-time PCR)相關試劑(北京Aidlab),完全培養基(dulbecco’s modified eagle medium,DMEM)和無糖培養基(美國Gibico),胎牛血清(以色列Biological Industries),凋亡相關蛋白裂解半胱天冬酶(cleaved cysteine-dependent aspartate-specifc proteases,Cleaved-caspase3)、B淋巴細胞瘤-2基因(B-cell lymphoma-2,Bcl2)、Bcl2-Associated X的蛋白質(Bcl2-Associated X,Bax)及BCL2L2抗體(美國CST);-80 ℃冰箱(中國HAIER),金屬浴(上海其林貝爾),臺式冷凍離心機(德國EPPendorf),電泳儀(北京百晶),CO2培養箱(美國Thermor)。
1.2 研究方法
1.2.1生物信息學分析 利用數據庫Targetscan 7.2(http://www.targetscan.org/vert_72/)預測miR-214-3p和BCL2L2之間有無結合位點。
1.2.2細胞培養 取PC12細胞培養于含10%胎牛血清的DMEM培養基中,培養條件為37 ℃、5%CO2,此方法下培養的即為PC12細胞的正常組(Control組),根據實驗情況選擇繼續傳代培養或用于實驗。取293T細胞培養于含10%胎牛血清的DMEM培養基中,培養條件為37 ℃、5%CO,根據實驗情況選擇繼續傳代培養或用于實驗。
1.2.3熒光素酶報告基因實驗 構建帶有熒光素酶報告基因的3′UTR野生型/突變型的BCL2L2質粒,合成miR-214-3pmimic/mimic NC質粒。取正常生長且細胞密度為50%時的293T細胞,PBS輕柔洗滌兩次,選擇一部分孔將野生型BCL2L2 3′UTR同miR-214-3pmimic共轉染(wt+mimic組),對照組為野生型BCL2L2 3′UTR同mimic NC共轉染(wt+NC組),另一部分孔則是將突變型BCL2L2 3′UTR同miR-214-3pmimic共轉染(mut+mimic組),對照組為突變型BCL2L2 3′UTR同mimic NC共轉染(mut+NC組)。轉染6h后更換培養基繼續培養,轉染48h后收集細胞,按照試劑盒說明書進行操作,最后使用熒光素酶活性發光計(promega corporation)分別測量四個組別的Renilla luciferase值,此即為報告基因發光值。
1.2.4OGD/R模型構建 選取生長狀態良好的PC12細胞,胰酶消化,1 000 r/min離心3~5 min,用含血清的完全培養基混成密度為5×109~5×1011個/L的懸液,吸取懸液100 μL加到含有培養基2 mL的6孔板,待孔內細胞密度為70%時,吸掉培養孔內的培養基,用磷酸鹽緩沖液輕柔洗滌2次,加無糖培養基2 mL,置于37 ℃的含5%CO2、94%N2及1%O2恒溫培養箱中培養4 h,更換培養基為含10%胎牛血清的DMEM完全培養基,更換培養條件37 ℃、5%CO2,繼續培養18 h,為OGD/R組。
1.2.5PC12細胞的轉染 選取正常生長的PC12細胞,用含血清的培養基混制成密度為5×108個/L的細胞懸液,接種到6孔板中,待密度達50%時,分別在相應的孔進行以下轉染操作:轉染miR-214-3pmimic(mimic組)、mimic NC(mimic NC組)、miR-214-3pinhibitor(inhibitor組)、inhibitor NC(inhibitor NC組),轉染后培養4~6 h,更換新鮮培養基繼續培養48 h。
1.2.6PC12細胞的OGD/R+miR-214-3pmimic模型 轉染miR-214-3pmimic到PC12細胞后,進行缺糖缺氧4 h再灌注18 h,為OGD/R+mimic組。
1.2.7miR-214-3p和BCL2L2 mRNA水平 采用實時熒光定量聚合酶鏈式反應(Real-time fluorescent quantitative PCR,Real-time PCR)檢測, 用TRIZOL法從Control組、OGD/R組PC12細胞的總RNA并測定濃度,取 0.5 μg進行逆轉錄獲取cDNA(TUREscript 1st Stand Cdna SYNTHESIS kit);取cDNA 1 μL同SYBR及相應的引物混合制成10 μL的體系,上機循環;miR-214-3p的循環條件為95 ℃ 3 min,95 ℃ 10 s、60 ℃ 30 s,39個循環,70 ℃ 15 s完成反應;BCL2L2的循環條件為95 ℃ 3 min,95 ℃ 10 s、60 ℃ 30 s,40個循環,70 ℃、15 s完成反應。引物序列見表1。miR-214-3p以U6作為內參照,BCL2L2以GAPDH作為內參照,分別計算miR-214-3p和BCL2L2 mRNA的相對表達水平。

表1 引物序列Tab.1 Primer sequence
1.2.8BCL2L2、Cleaved-caspase3、Bax、Bcl2蛋白表達 采用蛋白免疫印跡(Western blot)進行檢測,提取mimic組、mimic NC組、inhibitor組、inhibitor NC組總蛋白,檢測BCL2L2表達;提取Control組、OGD/R組及OGD/R+mimic組總蛋白,檢測Cleaved-caspase3、Bax、Bcl2表達。各組別用蛋白裂解液充分裂解總蛋白后,收集總蛋白并利用BCA定量試劑盒對蛋白質進行定量,根據定量結果將不同組蛋白按相同的量與5×SDS上樣緩沖液按4 ∶1混合,100 ℃煮沸10 min變性蛋白;SDS-PAGE電泳、轉膜2 h、5%牛血清白蛋白(bovine serum albumin,BSA)封閉液封閉2 h,BCL2L2、Cleaved-caspase3、Bax、Bcl2及內參一抗4 ℃孵育過夜,相應二抗室溫孵育2 h,曝光。
1.3 統計學分析
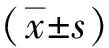
2 結果
2.1 miR-214-3p靶向調控BCL2L2的表達
選用數據庫Targetscan 7.2進行預測,結果顯示miR-214-3p與野生型BCL2L2的3′UTR端存在結合位點,而與3′UTR發生突變的BCL2L2之間未發現結合位點;在293T細胞中,熒光素酶報告基因結果顯示,與wt + NC組比較,wt + mimic組中熒光素酶的活性降低(P<0.05),而mut + NC組與mut + mimic組熒光素酶活性未發生明顯變化(P>0.05);在PC12細胞中,與mimic NC組比較,mimic組BCL2L2蛋白表達下調,與inhibitor NC組比較,inhibitor組BCL2L2蛋白表達上調(P<0.05)。見圖1。

注:A為數據庫預測結合位點(加粗字母表示預測結合序列位點),B為293T細胞雙熒光素酶報告基因結果,C、D分別為PC12細胞中BCL2L2的蛋白表達和定量結果;(1)與wt+NC組或mimic-NC組比較,P<0.05;(2)與inhibitor-NC組比較,P<0.05。圖1 miR-214-3p靶向調控BCL2L2蛋白表達Fig.1 miR-214-3p inhibited BCL2L2 expression via sponging BCL2L2
2.2 miR-214-3p與BCL2L2的表達
與Control組比較,OGD/R組PC12細胞中BCL2L2的mRNA和蛋白表達水平降低、miR-214-3pmRNA表達升高,差異均有統計學意義(P<0.05)。見圖2。

注:A、B分別為BCL2L2蛋白表達和定量結果,C、D分別為BCL2L2和miR-214-3p mRNA表達量;(1)與Control組比較,P<0.05。圖2 PC12細胞miR-214-3p與BCL2L2的表達Fig.2 The expression levels of miR-214-3p and BCL2L2 in PC12 cell
2.3 相關凋亡蛋白的表達
免疫印跡結果顯示,與Control組相比,OGD/R組PC12細胞中凋亡蛋白Cleaved-caspase3和Bax的表達增加、抗凋亡蛋白Bcl2的表達降低,差異均有統計學意義(P<0.05);與OGD/R組比較,OGD/R+mimic組PC12細胞中Cleaved-caspase3和Bax的表達增加、Bcl2的表達降低(P<0.05)。見圖3。

注:A為Cleaved-caspase3、Bax及Bcl2蛋白的電泳結果,B、C及D分別為Cleaved-caspase3、Bax及Bcl2蛋白表達的定量結果;(1)與Control組比較,P<0.05; (2)與OGD/R組比較,P<0.05。圖3 miR-214-3p通過靶向BCL2L2調控凋亡蛋白表達Fig.3 miR-214-3p regulated the expression levels of apoptosis-related proteins by targeting BCL2L2
3 討論
腦卒中的高發病率、高致殘及高死亡率給世界醫療衛生和經濟造成重大負擔,尋找有效治療腦卒中的方案依舊亟待解決。近年研究發現,miRNA在腦卒中的發生發展中發揮一定作用,miRNA不僅能作為一種潛在的疾病治療靶標,其異常表達也可作為疾病早期的診斷指標[16-17]。而miR-214-3p主要與細胞的增殖、遷移及生存等相關,如miR-214-3p通過靶向細胞黏附分子促進骨肉瘤的增殖、遷移及浸潤[18-20]。近年來還發現miR-214-3p能參與細胞凋亡的調控,神經元凋亡是腦卒中的重要病理特征之一[21-24]。考慮到miRNA可參與基因表達的調控,從而參與細胞多種生理或病理過程,因此本研究旨在探究miR-214-3p是否能調控凋亡相關蛋白的表達,從而參與調控神經元的凋亡。
本研究首先利用Targetscan 7.2在線數據庫進行預測,發現miR-214-3p與野生型的BCL2L2的3′UTR區之間存在結合位點;接著在293T細胞上進行熒光素酶基因報告實驗,實驗結果顯示,與wt+mimic NC組相比,wt+mimic組熒光素酶活性下降,而當BCL2L2 3′UTR位點發生突變時,mut+mimic NC組和mut+mimic 組熒光素酶活性均未發生明顯變化。這近一步說明miR-214-3p能靶向結合并下調BCL2L2表達。
為了進一步驗證設想,本研究在PC12細胞上進行以下實驗。首先對PC12細胞進行缺糖缺氧再灌注處理,成功構建腦卒中細胞模型后,實時熒光定量結果顯示,與Control組比較,經OGD/R處理后的PC12細胞中miR-214-3p表達上調,而抗凋亡基因BCL2L2 mRNA表達下調,同時免疫印跡結果顯示BCL2L2蛋白表達均下調。BCL2L2作為Bcl2的家族成員之一,同樣具有抗凋亡的作用[25],當BCL2L2的表達受到抑制后,抗凋亡作用減弱,細胞凋亡增加。因此推測,miR-214-3p的異常表達可能與腦卒中的發生相關,且miR-214-3p很有可能是通過靶向調控BCL2L2來參與疾病的發生。
Fan等[26]研究表明,下調miR-214-3p表達能減輕脊髓神經元的凋亡。本研究結果顯示,與Control組相比,OGD/R組PC12細胞的BCL2L2表達下調,同時還檢測到抗凋亡蛋白Bcl2的表達也發生下調,凋亡蛋白Bax和Cleaved-caspase3的表達增加;在PC12細胞中轉染miR-214-3pmimic再進行OGD/R處理后,與OGD/R組相比,BCL2L2和Bcl2的表達下調,凋亡蛋白Bax和Cleaved-caspase3的表達增加。這一結果表明,過表達miR-214-3p,可以減少抗凋亡蛋白的表達,上調凋亡蛋白的表達,從而促進神經元的凋亡,這與前述報道一致。
綜上所述,本研究表明miR-214-3p可能通過靶向抑制抗凋亡蛋白BCL2L2的表達,繼而上調凋亡蛋白的表達,使神經元的凋亡加重,從而參與腦卒中的發生。雖然本研究僅在細胞層面進行,未深入到動物層面,但這一探討為后續的研究提供了新思路。
猜你喜歡
中老年保健(2021年3期)2021-08-22 06:50:04
天津醫科大學學報(2021年2期)2021-03-29 05:31:08
昆明醫科大學學報(2021年1期)2021-02-07 01:06:36
現代臨床醫學(2021年1期)2021-01-26 00:56:02
昆明醫科大學學報(2020年12期)2021-01-26 00:44:04
中華養生保健(2020年4期)2020-11-16 01:31:40
中西醫結合肝病雜志(2020年2期)2020-10-27 02:18:50
豬業科學(2018年8期)2018-09-28 01:27:38
中成藥(2017年8期)2017-11-22 03:18:47
川北醫學院學報(2015年5期)2015-12-05 08:22:29
