川西亞高山天然次生林不同演替階段土壤-微生物生物量及其化學計量特征
2021-08-02 10:39:30胡宗達劉世榮劉興良羅明霞李亞非石松林吳德勇肖玖金
生態學報 2021年12期
胡宗達, 劉世榮,劉興良,胡 璟,羅明霞,李亞非,石松林,吳德勇,肖玖金
1 四川農業大學資源學院,成都 611130 2 中國林業科學研究院森林生態環境與保護研究所,國家林業和草原局森林生態環境重點實驗室,北京 100091 3 四川省林業科學研究院,成都 610081 4 湖南農業大學資源環境學院,長沙 410128 5 成都理工大學旅游與城鄉規劃學院,成都 610059 6 四川農業大學林學院,成都 611130
土壤微生物在土壤養分供給和轉換過程中扮演十分重要的角色,其生物量被認為是土壤養分循環過程中的源或匯[1-2];微生物生物量(Soil microbial biomass,SMB)含量高低可反映土壤有機質周轉率、土壤活力大小以及生態系統生產力水平[3-5],在維持土壤營養物質循環過程中具有十分重要的作用。土壤微生物的分解和礦化效率則主要取決于土壤SMB化學計量比[2]。森林植被恢復演替引起土壤微生境理化性質、物種組成結構、根際分泌物、枯落物數量和質量等生物或非生物因素變化,使其土壤微生物群落結構及功能發生改變[6-7],導致土壤SMB隨森林演替年限或林齡增加出現逐漸增加[8-9]、先升后降[10-11]或變化不顯著[12]等趨勢,其化學計量比亦出現類似變化趨勢,但二者協同變化趨勢不盡相同[2,13],例如:Song等[14]研究發現,土壤微生物生物量碳(Cmic)、氮(Nmic)和磷(Pmic)含量隨植被演替呈逐漸增加及先升后降的變化趨勢,而Cmic∶Nmic、Cmic∶Pmic和Nmic∶Pmic則分別呈降低、先升后降和逐漸增加趨勢;周正虎和王傳寬[15]整合了分析13個森林和6個草地生態系統土壤-微生物生態化學計量的時間動態發現,Cmic、Nmic和Pmic含量隨演替顯著增加,而Cmic∶Nmic∶Pmic則未表現出普適性的時間變化格局。可見,植被演替對SMB及其化學計量特征的影響源于主導因子不同而表現出相異的時空變化格局。
SMB化學計量比(Cmic∶Nmic∶Pmic)常被用作土壤肥力評價的關鍵指標[4,16],以反映土壤養分及微生物生長的限制狀況[15,17];土壤微生物在土壤生境中具有維持自身Cmic、Nmic、Pmic組成比處于相對穩定的能力[18],然Cmic∶Nmic∶Pmic變化的實質性原因主要來自微生物群落結構的變化[19],亦受到土壤養分影響,進而改變了Cmic∶Nmic∶Pmic與土壤Csoil∶Nsoil∶Psoil的協同變化關系。此外,土壤Csoil、Nsoil、Psoil的利用效率可用土壤微生物熵(qMB)表征;qMB受土壤有機質數量和質量以及微生物可利用底物有效性的影響較大[20-21],常被用于預測土壤養分庫的細微變化、土壤退化程度監測及其恢復效果評價[5,22]。雖有眾多研究探討了植被恢復演替不同階段的qMB變化的時空格局[10,21,23],但尚未得出一致性結論。因此,進一步探索森林演替進程中SMB及其化學計量比與qMB的變化特征,分析土壤及土壤微生物C∶N∶P與qMB的耦合作用,有益于更好地理解森林演替進程中SMB和qMB的動態變化特征,這對維持或改善森林土壤質量尤為必要。
川西亞高山米亞羅林區屬于岷江上游的生態屏障,20世紀50—80年代曾大規模采伐,后經人工恢復及實施封山育林后,現已形成多種不同演替階段的人工和天然次生林鑲嵌的森林景觀格局。該林區SMB研究主要集中在不同林型[24]、林窗[25]、林線交錯帶[26]、凍融[27]等方面,其研究結果顯示,SMB和土壤微生物活性對人為和自然因素影響的響應具有十分重要的表征功能。然因該區域植被類型及形成時間不同,SMB及其活性差異較大,為了更好地理解天然次生林(簡稱次生林)演替進程中SMB的影響因素,以及土壤-微生物化學計量比與qMB的關系,仍需要對次生林不同恢復演替階段SMB及其化學計量比的關系進行深入探究。本研究采集米亞羅林區 3 種天然更新恢復演替階段的次生林和岷江冷杉(Abiesfaxoniana)原始林表層(0—20 cm)土樣,測定土壤有機碳(Csoil)、全氮(Nsoil)、全磷(Psoil)以及微生物生物量碳(Cmic)、氮(Nmic)、磷(Pmic)含量,計算它們的C∶N∶P和qMB,分析比較其差異,旨在探究:(1)不同恢復階段SMB以及其化學計量比的變化特征;(2)次生林恢復演替對SMB含量影響、qMB與C∶N∶P化學計量的耦合關系及其qMB對次生林演替的響應。研究結果可為有效經營和管理次生林提供科學依據,亦有助于加深對該區域土壤碳氮磷地球化學循環特征的認識。
1 材料和方法
1.1 研究區概況
理縣米亞羅林區位于青藏高原東部,受高山-亞高山高原地形的影響,具冬寒夏涼高山氣候特點,為半濕潤地區。年平均溫度6—12 ℃,1月平均氣溫 -8 ℃,7月平均氣溫12.6 ℃,年無霜期約200天,年均降雨量700—1400 mm,年蒸發量1000—1900 mm,冬季雪被期長達6—7個月;成土母巖主要為板巖、片巖、白云巖等為主,土壤類型為山地棕壤。該區域頂極群落以高山-亞高山暗針葉林為主,喬木層優勢物種有岷江冷杉、紫果云杉(Piceapurpurea)、粗枝云杉(P.asperata)、紅樺(Betulaalbo-sinensis)、白樺(B.platyphylla)、槭樹(Acerspp.)、皂莢(Gleditsiasinensis)、椴樹(Tiliatuan)等;灌木層優勢種分布有川滇高山櫟(Quercusaquifolioides)、陜甘花楸(Sorbuskoehneana)、野櫻桃(Prunstatsienensis)、紫花衛矛(Euonymusporphyreus)、高山柳(Salixcupularis)、康定柳(S.paraplesia)、紅刺懸鉤子(Rubusrubrisetulosus)、多種忍冬(Loniceraspp.)、莢蒾(Viburnumspp.)、華西箭竹(Fargesianitida)等;草本優勢種主要有珠芽蓼(Polygonumviviparum)、東方草莓(Fragariaorientalis)、糙野青茅(Deyeuxiascabrescens)、光柄野青茅(Deyeuxialevipes)、掌葉橐吾(Ligulariaprzewalskii)、嵩草(Kobresiaspp.)等植物。
1.2 土壤樣品采集和處理
2019年7月下旬,在米亞羅林區大郎壩、盡頭寨、十溝和十八拐溝上寨,按照立地條件相似原則,選擇20世紀60、70和80年代3種采伐跡地經自然恢復演替形成的天然次生林(簡化為SF60、SF70和SF80)和岷江冷杉原始林(簡化為PF,林齡為160年)為一個演替系列。每種次生林和原始林用測繪儀(博飛,DZS3-1,中國)隨機布設3個20 m×20 m間隔大于20 m的標準樣地作為試驗的3個重復和一個對照,并進行植被生態學常規調查(表1)。土壤采樣時,在已布設的標準樣地中,將其按照“田”字形再次分為4個10 m×10 m的樣方,在每個樣方中以“梅花5點”法用不銹鋼土鉆(Eijkelkamp, 型號 07.53.SC)采集0—20 cm的表層土樣,將20個取樣點的土壤充分混勻后,按照“四分”法取一個土樣裝入無菌塑料密封袋,置于有冰袋的冷藏箱運回實驗室,去除石礫、殘留根系及枯落物后過2 mm篩。將過篩樣品分為2份:一份新鮮樣品,用于測定土壤微生物生物量碳(Cmic)、氮(Nmic)、磷(Pmic)含量;一份經自然風干,用來測定土壤有機碳(Csoil)、全氮(Nsoil)和全磷(Psoil)含量等。
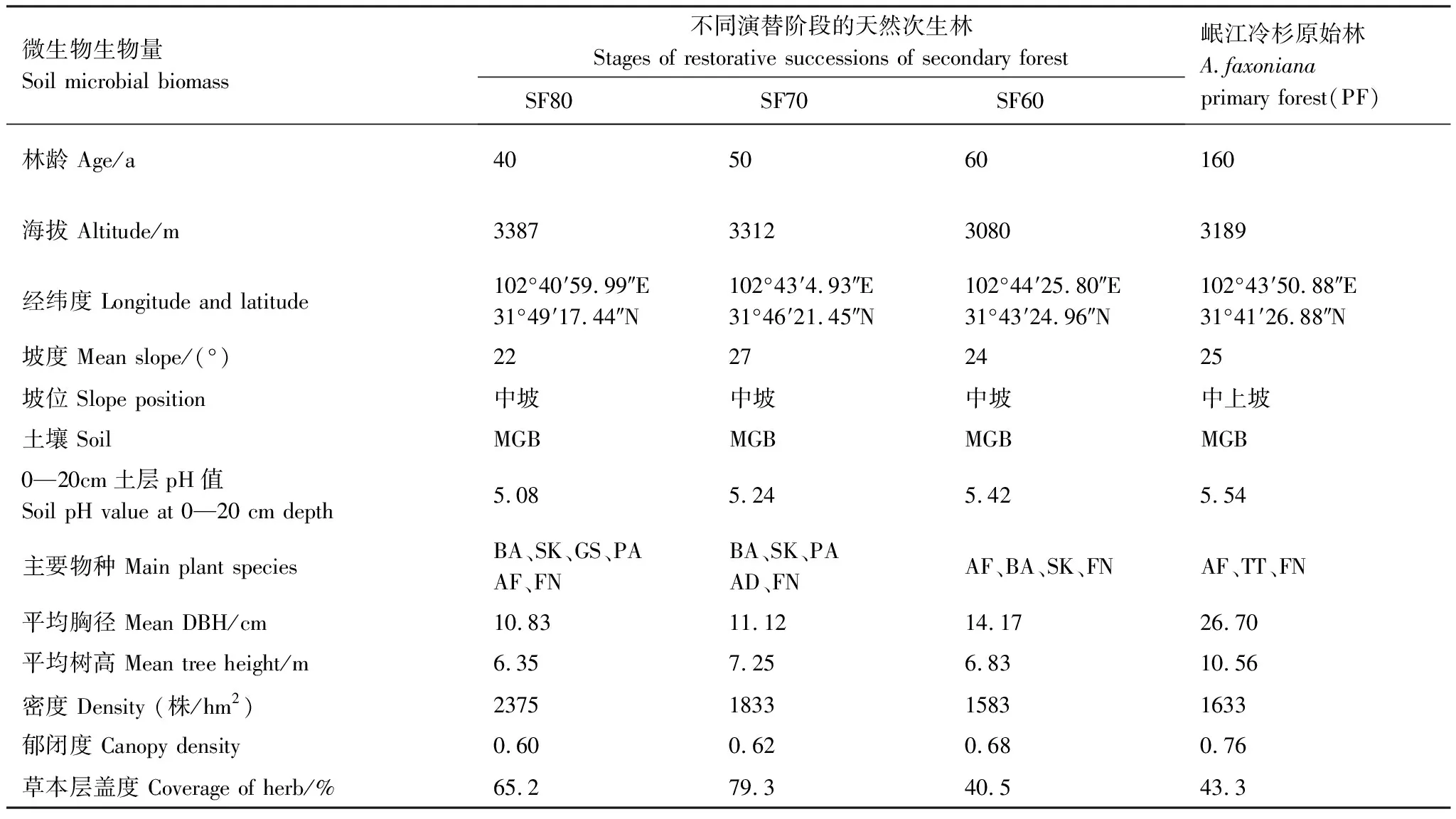
表1 不同更新階段天然次生林和岷江冷杉原始林樣地基本特征
1.3 測定方法
土壤 pH 用土水質量比1∶2.5混合攪拌后用 pH 計測定;土壤有機碳(Csoil)含量測定采用重鉻酸鉀外加熱法;全氮(Nsoil)含量測定用硫酸-催化劑消解-AA3連續流動分析儀法(Auto Analyzer 3, SEAL,德國);全磷(Psoil)含量測定用鉬銻抗比色法(島津UV-2450紫外分光光度計,日本島津)。
土壤微生物生物量碳(Cmic)、氮(Nmic)和磷(Pmic)含量測定采用氯仿熏蒸浸提法[28]:各取過2 mm篩新鮮土樣2份(Cmic和Nmic樣品均為7.5 g、Pmic為2.5 g),分別放置于有氯仿和無氯仿的真空干燥器,在暗箱內熏蒸24 h后取出,其中測定Cmic、Nmic樣品加入土液體積比為1∶4的 0.5 mol/L K2SO4水溶液,震蕩30 min后過濾,分別采用TOC-VCPH有機碳分析儀和AA3連續流動分析儀進行測定;測定Pmic則加入土液體積比為1∶20的 0.5 mol/L碳酸氫鈉溶液,震蕩30 min,吸取20 mL液體,加5 mL硫酸-鉬銻-抗混合顯色劑,定容至34 mL,用島津UV-2450紫外分光光度計比色測定。Cmic含量=(熏蒸土壤有機碳-未熏蒸土壤有機碳)/0.45,Nmic含量=(熏蒸土壤有機氮-未熏蒸土壤有機氮)/0.25,Pmic含量=(熏蒸土壤磷-未熏蒸磷)/0.40,式中0.45、0.25和0.40為轉換系數。
1.4 數據分析
根據Mooshammer等[29]以及周正虎和王傳寬[15]的方法,土壤碳、氮、磷及其微生物生物量化學計量特征采用質量比表示,即分別為Csoil∶Nsoil、Csoil∶Psoil、Nsoil∶Psoil和Cmic∶Nmic、Cmic∶Pmic、Nmic∶Pmic;土壤微生物熵qMBC、qMBN和qMBP分別用(Cmic/Csoil)× 100%、(Nmic/Nsoil)×100%和(Pmic/Psoil)×100%表示;土壤微生物化學計量不平衡性用Cimb∶Nimb、Cimb∶Pimb和Nimb∶Pimb表示,其比值越大說明資源質量越低,土壤微生物生長效率也越低[15],計算公式如下:
Cimb∶Nimb=[(Csoil÷Nsoil)/(Cmic÷Nmic)]
(1)
Cimb∶Pimb=[(Csoil÷Psoil)/(Cmic÷Pmic)]
(2)
Nimb∶Nimb=[(Nsoil÷Psoil)/(Nmic÷Pmic)]
(3)
采用單因素方差分析和最小顯著差異法(LSD)檢驗3種次生林以及次生林與岷江冷杉原始林之間的土壤-微生物生物量各指標的顯著性差異(水平為P<0.05)。用 Pearson相關法分析評價SMB含量及其化學計量比的影響因子。以上統計分析均由SPSS 26.0 軟件完成(顯著性水平均為0.05)。此外,利用Canoco 5.0軟件對微生物熵(qMB)與土壤-微生物生物量C∶N∶P化學計量之間的關系進行冗余分析(RDA);繪圖用Sigmaplot 14.0軟件完成;圖、表中數據均為平均值±標準偏差。
2 結果與分析
2.1 土壤碳、氮、磷含量及其化學計量特征
3種次生林土壤Csoil、Nsoil平均含量顯著低于PF(P<0.05),而Psoil的平均含量略低于PF (圖1)。Csoil和Nsoil含量隨次生林更新年限呈顯著下降趨勢,SF80、SF70和SF60較PF分別低21.15%和29.74%、26.61%和23.93%、61.98%和71.97%;Psoil含量則隨恢復演替進程呈先增后降的顯著變化趨勢,其中SF70最高,比PF高5.66%(P>0.05),SF80和SF60之間的Psoil含量差異不顯著(P>0.05)。
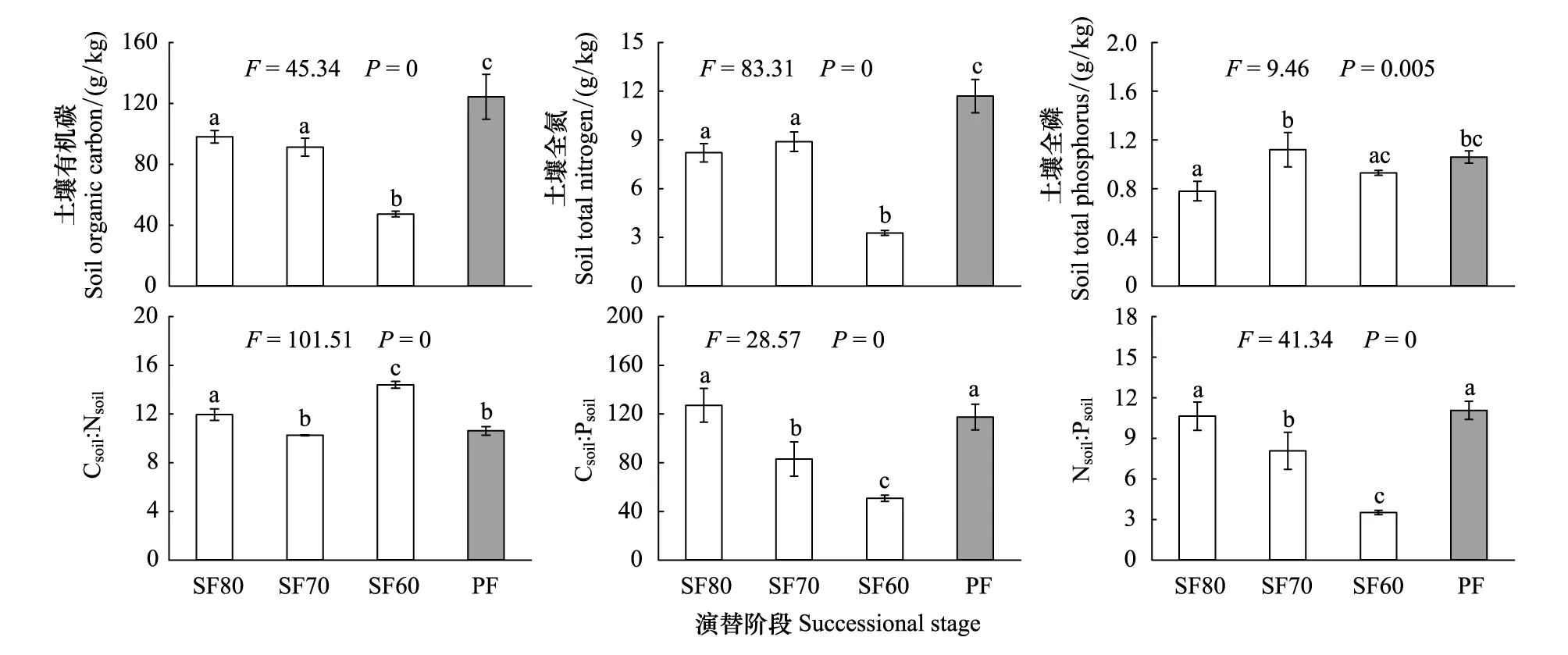
圖1 天然次生林不同演替階段及岷江冷杉原始林土壤C、N、P含量及化學計量變化特征 (n=3)Fig.1 Change characteristics of Csoil、Nsoil and Psoil content and its stoichiometric ratios of topsoil from sub-alpine natural secondary forests with different stages of restoration and A.faxoniana primary forest (n=3)SF80: 20世紀80年代砍伐跡地的天然次生林 Natural secondary forest from clear-logging area during the 1980s; SF70: 20世紀70年代砍伐跡地的天然次生林 Natural secondary forest from clear-logging area during the 1970s; SF60: 20世紀60年代砍伐跡地的天然次生林 Natural secondary forest from clear-logging area during the 1960s; PF: 岷江冷杉原始林 Abies faxoniana primary forest; Csoil∶Nsoil: 土壤碳氮比 Soil C∶N ratio; Csoil∶Psoil: 土壤碳磷比 Soil C: P ratio; Nsoil∶Psoil: 土壤氮磷比 Soil N∶P ratio
土壤Csoil∶Nsoil隨次生林演替呈先降后升的顯著變化趨勢(圖1),在SF60中達到最大(14.40),其中SF70與PF差異不顯著(P>0.05);Csoil∶Psoil與 Nsoil∶Psoil高低均表現為SF80>SF70>SF60(P<0.05),與PF相比,SF80分別高了8.19%和低了3.88%;SF70、SF60分別低了29.47和27.01%、56.83和68.20%。從圖1可知,SF80、SF70、SF60和PF中的土壤Csoil∶Nsoil∶Psoil分別為126∶ 11∶1、82∶8∶1、51∶4∶1和118∶11∶1。
2.2 土壤微生物生物量及其化學計量特征
3種次生林土壤Cmic、Nmic、Pmic含量均低于PF(表2),其中Cmic和Pmic含量隨次生林演替呈先增后降的變化趨勢,其中SF60表層土壤Cmic含量比SF80和SF70分別下降了47.73%和52.51%(P<0.05),Pmic含量分別降低了15.48%(P>0.05)和38.29%(P<0.05);Nmic含量隨次生林演替年限呈顯著遞減趨勢,SF60較SF80和SF70分別下降了40.81%和28.31%。
3種次生林的Cmic∶Nmic高于PF(表2)。方差分析顯示,Cmic∶Nmic在SF80和SF70及SF70和SF60之間差異顯著(P<0.05),而在SF80和SF60間差異不顯著(P>0.05);Cmic∶Pmic在SF80和SF70及SF70和SF60之間差異不顯著(P>0.05);Nmic∶Pmic在不同次生演替階段無顯著性差異(P>0.05)。此外,從表2統計分析發現,SF80、SF70、SF60和PF中,土壤微生物生物量Cmic∶Nmic∶Pmic分別為23∶7∶1、19∶4∶1、14∶5∶1和18∶7∶1。

表2 天然次生林不同更新階段和岷江冷杉原始林土壤微生物生物量碳、氮和磷含量及其化學特征(n=3)
2.3 土壤微生物熵和微生物化學計量不平衡性特征
圖2可看出,次生林近60年的恢復演替對qMBC 和qMBP 未產生顯著影響(P>0.05),但對qMBN影響極顯著(P=0.003)。土壤微生物熵(qMB)隨次生林恢復演替年限增加呈現不同的變化趨勢:qMBC表現為先增后降的變化趨勢(P>0.05),到PF階段達到最低(0.97);qMBN表現為先降后升的極顯著變化趨勢(P<0.01),其中SF60顯著高于SF80、SF70和PF,分別高出49.5%、94.1%和34.29%;qMBP隨次生林演替呈逐漸降低趨勢(P>0.05)。
次生林恢復演替進程中,土壤Cimb∶Nimb、Cimb∶Pimb、Nimb∶Pimb變化趨勢差異較大(圖2)。Cimb∶Nimb隨次生林演替進程呈極顯著的先降后升變化趨勢(P=0.002),在SF60達到最高,但與PF差異不顯著(P>0.05);Cimb∶Pimb和Nimb∶Pimb表現出輕微的(P=0.175)或顯著的逐漸降低趨勢(P=0.039),其中SF60的Cimb∶Pimb顯著低于PF; SF80和SF70的Cimb∶Pimb和Nimb∶Pimb與PF相比,差異性不顯著(P>0.05)。
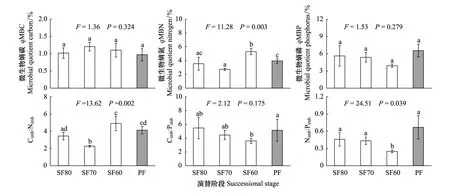
圖2 天然次生林不同演替階段和岷江冷杉原始林土壤微生物生物量和土壤微生物化學計量不平衡性的變化特征(n=3)Fig.2 Change characteristics of soil microbial biomass and soil microbial stoichiometry imbalance ratios of topsoil from sub-alpine natural secondary forests with different stages of restoration and A.faxoniana primary forest (n=3)PF: 岷江冷杉原始林 Abies faxoniana primary forest; Cimb∶Nimb: 微生物生物量碳氮化學計量不平衡性 Microbial biomass C∶N stoichiometric imbalance; Cimb∶Pimb: 微生物生物量碳磷化學計量不平衡性 Microbial biomass C∶P stoichiometric imbalance; Nimb∶Pimb: 微生物生物量氮磷化學計量不平衡性 Microbial biomass N∶P stoichiometric imbalance
2.4 土壤碳氮磷和土壤-微生物生物量化學計量之間的相互關系
Pearson相關分析(表3)顯示,Csoil與Nsoil含量極顯著正相關(P<0.01),Csoil含量和Nsoil與Psoil含量正相關(P>0.05);Csoil和Nsoil含量與Cmic、Nmic、Pmic含量極顯著(P<0.01)或顯著(P<0.05)正相關;Cmic、Nmic、Pmic含量兩兩之間呈極顯著或顯著正相關。Csoil∶Nsoil與Csoil、Nsoil和Cmic和Pmic極顯著負相關;Csoil∶Psoil和Nsoil∶Psoil與Csoil、Nsoil和Cmic和Nmic極顯著或顯著正相關;Nsoil∶Psoil與Csoil∶Nsoil和Csoil∶Psoil分別呈極顯著負相關和正相關;Nmic∶Pmic與Nmic和Cmic∶Pmic分別呈顯著和極顯著正相關。
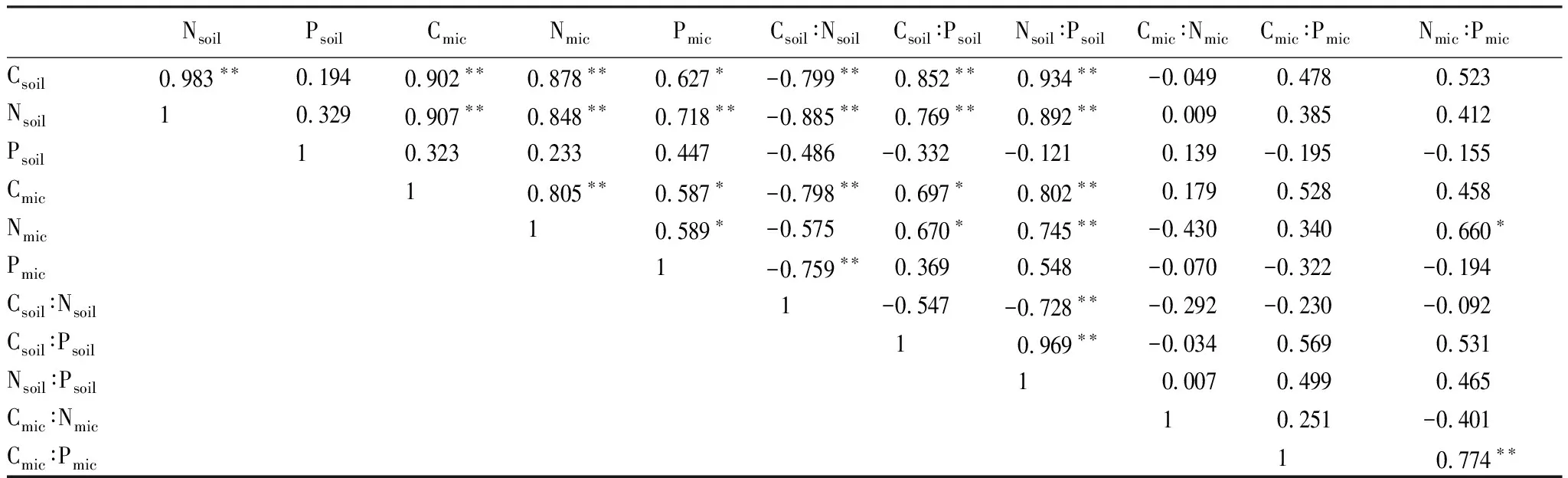
表3 土壤-微生物生物量碳氮磷及其化學計量比之間的相關系數(n=12)
SMB化學計量不平衡性與土壤-微生物生物量及其化學計量比的相關性分析(表4)顯示,Cimb∶Nimb與Cimb∶Pimb相關性不顯著(P=0.822);Nimb∶Pimb分別與Nsoil和Cimb∶Pimb呈顯著正相關(P=0.026),而與Cimb∶Nimb顯著負相關(P=0.013);Cimb∶Nimb分別與Csoil∶Nsoil和Cmic∶Nmic顯著正相關(P=0.020)和極顯著負相關(P=0.000);Cimb∶Pimb與Pmic和Nsoil∶Psoil極顯著(P=0.005)和顯著正相關(P=0.035);Nimb∶Pimb分別與Nsoil和Nsoil∶Psoil顯著正相關(P=0.025和0.016),而與Pmic和Csoil∶Nsoil極顯著正相關和負相關(P=0.004和0.003)。
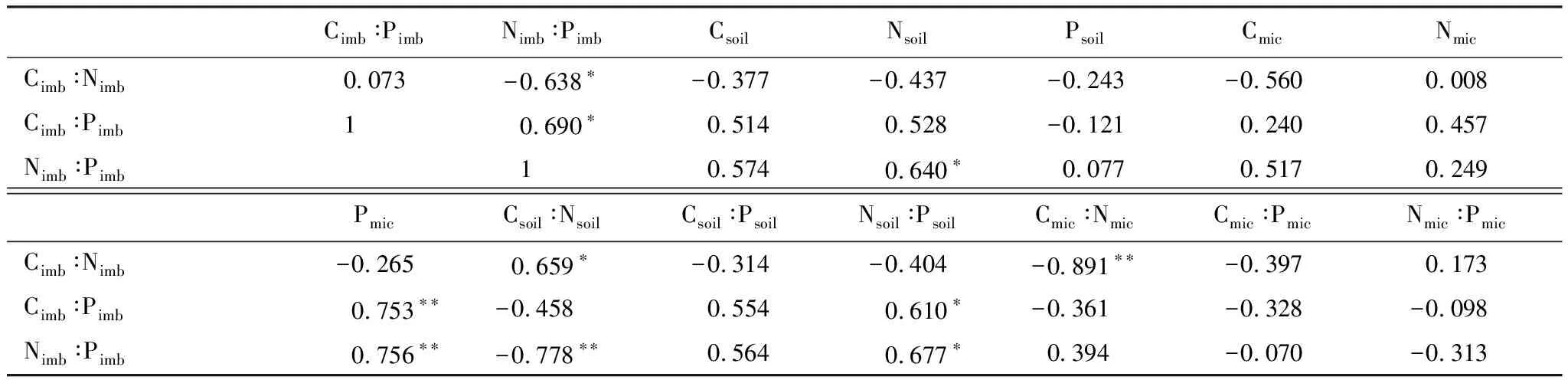
表4 土壤微生物生物量化學計量不平衡性與土壤微生物生物量化學計量比的相關性(n=12)
2.5 土壤微生物熵與土壤-微生物化學計量不平衡性的RDA分析
以qMB為響應變量,土壤-微生物化學計量比值為解釋變量進行冗余分析(圖3),結果顯示:Nimb∶Pimb分別與Cimb∶Nimb和Cimb∶Pimb負相關和正相關。qMBN與Csoil∶Nsoil、Cimb∶Nimb正相關,而與Cmic∶Nmic、Nimb∶Pimb負相關;qMBP與Csoil∶Psoil、Nsoil∶Psoil、Cimb∶Pimb、Nimb∶Pimb正相關。圖3可看出,第一軸和第二軸分別解釋了qMB變化的72.78%和24.51%,說明土壤-微生物化學計量比能很好地反映qMB變化。Nimb∶Pimb對qMB的影響最大,解釋了qMB變化的62.6%,其次為Cmic∶Nmic,解釋了qMB變化的21.7%。
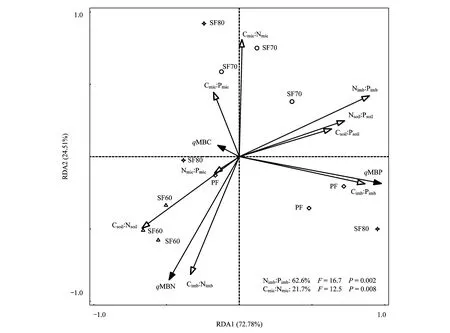
圖3 土壤微生物熵和土壤-微生物C∶N∶P化學計量的RDA排序圖Fig.3 Redundancy analysis of soil microbial entropy and soil-microbial C∶N∶P stoichiometryqMBC: 微生物熵碳 Microbial entropy carbon; qMBN: 微生物熵氮 Microbial entropy nitrogen; qMBP: 微生物熵磷 Microbial entropy phosphorus; Cmic∶Nmic: 微生物生物量碳氮比 C-to-N ratio of microbial biomass; Cmic∶Pmic: 微生物生物量碳磷比 C-to-P ratio of microbial biomass; Nmic∶Pmic: 微生物生物量氮磷比 N-to-P ratio of microbial biomass
3 討論
3.1 次生林恢復演替對土壤微生物生物量及其化學計量比的影響
3.1.1對土壤微生物生物量碳氮磷的影響
植被演替通過改變林分結構而影響到土壤底物有效性、微生物群落結構、土壤酶活性等[30-32],進而使其SMB具有不同的時空變化特征。本研究結果顯示,以闊葉樹種為主的SF80和SF70土壤SMB含量高于針葉樹種為主的SF60,與已有研究結果一致[33]。土壤Cmic和Pmic含量排序為SF60 本研究中,SMB含量在PF的達到最大值,說明在岷江冷杉原始林群落中,林木不斷生長、生物量逐漸積累、地表枯落物分解量以及根系分泌物增加,為土壤微生物提供了更多的食物來源,有利于土壤微生物的生長,土壤底物利用率高,如需明晰上述因素驅動SMB含量變化的機制,需進一步深入研究。相關分析表明,Cmic和Nmic含量與Csoil和Nsoil含量相互之間呈極顯著或顯著正相關,與已有研究結果一致[17,44],說明Csoil和Nsoil含量增加可提升土壤碳礦化水平及土壤酶活性[45],從而提高Cmic和Nmic含量,反之亦然;Pmic與Psoil關系不顯著,表明次生林短期恢復演替對Pmic的改善不如Cmic和Nmic明顯,究其原因:(1)次生林早期恢復演替階段進程中,SMB含量變化主要受受控于Csoil和Nsoil含量的變化[46];(2)土壤中Psoil含量高低與成土母質關系較大,且Psoil在土壤中轉移和代謝速率速率較慢[47],該區域中磷代謝速率、途徑和來源需要進一步探究。綜上,恢復提升采伐跡地天然次生林(尤其是1960s次生林)的土壤有效養分含量,需要采取合理營林措施及長期有效的管護監控。 3.1.2對土壤碳氮磷化學計量比的影響 森林植被恢復演替顯著影響到土壤Csoil、Nsoil和Psoil含量變化,進而導致土壤養分化學計量比的變化[15,48]。Csoil∶Nsoil可反映森林土壤底物被土壤微生物利用的有效性狀況,即Csoil∶Nsoil比值越高,土壤有機質礦化分解越慢,土壤質量越差,反之亦然[49]。本研究中,Csoil隨次生林演替呈顯著降低趨勢,Nsoil表現為先增后減的變化趨勢,但Csoil∶Nsoil則呈現出先減后增的顯著變化趨勢(圖1);3種不同演替階段的次生林中,Csoil∶Nsoil變幅為10.25—14.40(均值12.2),略高于中國亞熱帶地區平均水平(12.1)[48],其原因可能與次生林Csoil含量隨演替進程的降幅不及Nsoil含量的降幅有關(表2),說明次生演替導致林分結構及其物種組成的改變使其地表枯落物和地下根系中碳氮含量的時間耦合不同[50-51],進而影響到微生物群落組成發生變化,導致土壤有機質分解礦化及其利用程度不盡相同[52]。與PF相比,SF60中土壤碳氮礦化作用較慢,氮釋放少而使Csoil∶Nsoil顯著升高。 Csoil∶Psoil常被用作土壤磷礦化和固持能力的重要指標,以指示植物生長過程中土壤營養成分的供給狀況以及養分限制的閾值[48]。有研究指出,Csoil∶Psoil比值越低,越有利于微生物對土壤有機質分解,促進土壤有效磷的增加,反之存在Psoil受限[53];也有研究認為,Csoil∶Psoil比值高說明土壤微生物對土壤有效磷分解利用表現出同化趨勢,容易出現土壤微生物與地上植被競爭性吸收利用有效磷的情況,存在較強的Psoil固持潛力[49]。本研究中,Csoil∶Psoil和Nsoil∶Psoil在SF60達到最低值(圖1),而Csoil∶Nsoil在SF60含量最高,且Csoil∶Psoil和N∶Psoil與土壤Csoil和Nsoil極顯著正相關,與Psoil相關性不顯著(表3),究其原因:一方面,該研究區域的林下棕壤土壤磷素限制狀況低于其它土壤類型,有機磷凈礦化率較高,微生物分解受磷素影響較小[54];另一方面,隨著次生林的恢復演替,Csoil和Nsoil有效補給不足,地上植被生長對Csoil和Nsoil需求漸增使其Csoil和Nsoil含量處于較低水平(圖1)。另外,SF70、SF80和PF的Csoil∶Psoil和Nsoil∶Psoil高于中國亞熱帶的平均水平(78和6.4),而SF60則低于其平均水平[48],與已有研究結果相似[55]。綜合分析可知,Csoil和Nsoil是影響研究區土壤Csoil∶Psoil和Nsoil∶Psoil的關鍵因素。 3.1.3對土壤微生物生物量碳氮磷化學計量比的影響 Cmic∶Nmic∶Pmic是判定微生物群落結構變化的重要指標[18,29,56]。已有研究顯示,森林主要樹種組成及物種數量以及生物量、枯落物、根系等的數量和質量不同,其土壤微生物種類結構亦不同[57,58],致使SMB含量高低變化趨勢各異[29],進而影響到土壤微生物生物量的化學計量比。本研究結果顯示,次生林恢復演替對Cmic∶Nmic和Cmic∶Pmic產生顯著影響,對Nmic∶Pmic影響未達到顯著性水平,與已有研究結論相同[59]。隨著次生林的恢復演替,Cmic∶Nmic表現出先增后減的顯著變化趨勢;Cmic∶Pmic和Nmic∶Pmic則表現為隨演替進程呈逐漸降低趨勢,原因在于隨著次生林恢復演替,林分的主要樹種組成從闊葉樹種占優勢演變到針葉樹種占優勢(表1),使其地表枯落物、根系分泌物等的數量和質量不同,導致土壤微生物活性產生差異而使土壤SMB含量變化不同所致(表2)。有研究認為,土壤Cmic∶Nmic在3—6之間,表明土壤細菌占優勢,反之以真菌為主導[10]。研究區不同演替階段次生林土壤Cmic∶Nmic在3.00—4.53(均值3.68)和PF(2.60),低于全球和中國森林土壤的平均水平(8.2和8.25)[18,60],說明研究區土壤微生物以細菌為主;Nmic∶Pmic遠低于全球(6.9)平均水平[18],說明研究區域森林土壤微生物在一定程度上受到了磷素的限制(表2),此外,相關分析發現,Nmic∶Pmic與Nmic和Cmic∶Pmic極顯著或顯著正相關,不同于我國亞熱帶的研究結論[56],一方面表明SMB的化學計量特征隨生境變化而變化;另一方面可能是源于Nmic釋放受到Nsoil限制,進而修正了Nmic∶Pmic。雖然本研究結果分析了次生林演替對土壤Cmic∶Nmic∶Pmic的變化特征,但要明晰上述因素對土壤微生物生物量生態化學計量比的影響機理,仍需進一步針對Cmic∶Nmic∶Pmic與土壤微生物群落物種組成耦合關系作深入研究。 土壤微生物熵(qMB)是衡量土壤有機碳累積或損失的重要指標[15],比值越高表示土壤碳積累多[23]。有研究顯示,隨著植被演替時間尺度的延伸,qMB出現增加或降低趨勢[15,61],主要源于土壤底物有效性高低以及異養微生物群落的組成結構。研究發現,隨著森林的恢復演替,qMBC變化趨勢不顯著,說明研究區域森林土壤Csoil向Cmic轉化的效率相對穩定,其土壤碳庫穩定性較為良好;不同演替階段qMBC變幅為0.97%—1.21%(均值為1.08%),與其它區域的亞高山云杉(Piceamorrisonicola)森林(0.4%—1.2%)[62]和亞熱帶濕潤森林(0.7%—1.7%)[63]相近,但低于中國森林的平均水平(1.92%)[60];qMBN在SF60中顯著高于其他演替階段的林分類型,其變幅為2.74%—5.31%,高于亞高山云杉林(1.4%—2.3%)[62],而在SF60中(5.31%)明顯高于國森林平均水平(3.43%)[60],說明SF60土壤氮素活性較大,土壤氮庫穩定性較差,其養分含量較低;qMBP隨次生林恢復演替進程的降低趨勢不顯著,其變幅為3.92%—6.54%,具有相對較高的穩定性(圖2)。總之,不同演替階段qMB變化趨勢不同,主要在于次生林不同演替階段土壤微生物對C、N和P的積累和消耗取決于土壤微生物對有機質的分解[64]。根據土壤微生物化學計量比限制理論,微生物會首選吸收利用最適宜元素組分來維持自身化學計量比的平衡關系。從研究結果看,qMB比Csoil∶Nsoil∶Psoil和Cmic∶Nmic∶Pmic表現出更平穩的變化趨勢,能否用qMB作為衡量該研究區次生林土壤有機碳變化指標,值得商酌。 有研究顯示,qMB與Csoil∶Nsoil∶Psoil及其不平衡性指標之間存在顯著的正負相關關系[15,60],原因在于土壤微生物的生長代謝需要Csoil、Nsoil、Psoil等養分的綜合協調供給。本研究結果顯示,土壤-微生物生物量化學計量不平衡性指標中,Nimb∶Pimb分別與Cimb∶Pimb和Cimb∶Nimb顯著正相關和負相關,與Zhou等[60]的結果相似。從冗余分析結果看,Nimb∶Pimb和Cmic∶Nmic是影響qMB變化的關鍵因素,其中Nimb∶Pimb(解釋量為62.6%)起著主導作用,進一步證實了次生林演替進程中土壤微生物生長繁殖主要受到氮素和磷素的綜合協調影響,同時也表明土壤-微生物化學計量不平衡性在一定程度上能夠體現土壤微生物對土壤養分變化的適應性及其協同調節著森林生態系統養分動態的平衡關系[29]。依據生長速率理論,土壤微生物的高生長速率需要大量的蛋白質酶及核糖體RNA的投資,意味著土壤N∶P值的減小[65]。本研究中,土壤微生物Cmic∶Nmic以及Nimb∶Pimb與qMBN呈負相關關系;Csoil∶Psoil、Nsoil∶Psoil其化學計量不平衡性與qMBP正相關,說明Cmic∶Nmic和Nimb∶Pimb比值高意味著微生物生長受到N和P資源供給限制[15],導致土壤微生物對N和P元素分解效率降低而使Csoil∶Psoil、Nsoil∶Psoil及其化學不平衡性比值降低。綜上可知,森林植被演替僅對qMBN產生顯著影響,據此說明該研究區主要受到N資源的供給限制。此外,隨著全球氣候變暖、N沉降的加劇可能會影響次生林的生態功能,其地上植被和地下元素的平衡狀況及生態化學計量比如何響應,需要對植物-枯落物-土壤元素含量及其化學計量特征的耦合機理展開深入研究。為此,下一步研究重點應集中探究該生態系統元素間的耦合關系及其化學計量的季節動態變化對升溫、N沉降等全球變化的響應機理。 總體而言,隨著天然次生林恢復演替年限的逐漸增加,土壤有機碳、氮含量總體上呈顯著降低趨勢,土壤全磷含量呈增加趨勢;土壤微生物生物量生長更新到60年階段其含量最低;土壤及微生物生物量氮、磷的化學計量比值隨著恢復演替時間尺度延長而降低,土壤碳氮比呈先降后升的顯著變化趨勢;微生物生物量碳氮比與土壤碳氮比呈相反的變化趨勢;土壤-微生物生物量化學計量的不平衡性變化特征基本上能直接或間接地反映次生林演替進程中的土壤質量狀況。此外,該研究區主要受到氮素供給缺乏的限制,添加氮素或氮磷復合元素的供給可能會促進其次生林林木生長。3.2 次生林恢復演替對土壤微生物熵的影響
3.3 土壤-微生物C∶N∶P化學計量比對土壤微生物熵的影響
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
科技知識動漫(2017年7期)2017-08-09 19:52:45
汽車工程學報(2017年2期)2017-07-05 08:13:02
科技知識動漫(2017年5期)2017-05-11 21:34:16
科技知識動漫(2017年4期)2017-04-15 22:24:55
科技知識動漫(2017年2期)2017-02-06 20:59:46
