Microbulbifer sp. A4B-17菌株的磷酸烯醇式丙酮酸羧激酶與烯醇化酶研究
2021-08-02 11:53:51劉曹彤陸依琳徐慧彭學
江蘇農業科學 2021年12期
劉曹彤 陸依琳 徐慧 彭學
摘要:對羥基苯甲酸(4-Hydroxybenzoate,4HBA)應用廣泛,在工業領域上被當作前體合成各種芳香族化合物,其中包括液晶材料、農藥等;在食品和化妝品等領域上被當作防腐劑。微泡菌屬(Microbulbifer sp.) A4B-17是一種能將葡萄糖合成4HBA的海洋細菌,為了提高4HBA的合成效率對合成莽草酸途徑前體的2個關鍵酶:GM004356編碼的磷酸烯醇式丙酮酸羧激酶(Phosphoenolpyruvate carboxykinase,PEPCK)和GM0031333編碼的烯醇化酶(Enolase)進行研究。通過構建蛋白系統進化樹發現,PEPCK與Microbulbifer sp. GL-2的PEPCK相似度為95.15%,Enolase與Microbulbifer variabilis的Enolase相似度為98.13%。利用低溫表達載體在大腸桿菌中對2個酶進行高效表達,提取和純化后,采用BCA法蛋白定量計算后得到純化后的PEPCK和Enolase分別占各自粗酶液的1.304%和1.123%。酶活采用PK/LDH(丙酮酸激酶/乳酸脫氫酶)偶聯檢測法檢測NADPH的減少量。根據米氏常數雙倒數法求得PEPCK和Enolase的Km值分別為0.041 mmol/L和0.056 mmol/L,PEPCK反應的最適溫度為30 ℃,最適pH值為7,Enolase的最適溫度為 30 ℃,最適pH值為7。以上結果為高效生產4HBA提供了理論依據。
關鍵詞:對羥基苯甲酸;磷酸烯醇式丙酮酸;烯醇化酶;磷酸烯醇式丙酮酸羧激酶
中圖分類號:S188+.3 文獻標志碼: A
文章編號:1002-1302(2021)12-0045-06
收稿日期:2020-11-04
基金項目:江蘇師范大學科研創新計劃校立項目(編號:2020XKT485)。
作者簡介:劉曹彤(1996—),女,江蘇淮安人,碩士研究生,主要從事環境微生物學研究。E-mail:3220859786@qq.com。
通信作者:彭學,博士,教授,主要從事環境微生物學研究。E-mail:pengxue@jsnu.edu.cn。
對羥基苯甲酸(4-Hydroxybenzoate,4HBA)及其酯類物質,不僅能抑制真菌和細菌的生長,還能調節一些酶的代謝,應用非常廣泛[1],覆蓋了衣食住行各個方面,比如用于食品與化妝品的防腐劑、農業中的殺菌劑與化肥、手機電腦等的液晶制品等方面[2-6]。4HBA的毒性較低,抑菌效果也好,所以在2002年,對羥基苯甲酸酯類物質已經被我國批準作為食品防腐劑添加使用[7]。
目前大部分4HBA通過石油分餾[8]得到,也有報道利用植物[9]或者重組大腸桿菌[10]合成4HBA,但是這些方法存在很多的缺陷,植物合成得到的4HBA提純難度大[11]。微泡菌屬(Microbulbifer sp.) A4B-17菌株是能將葡萄糖合成4HBA及其酯類的海洋細菌,筆者所在課題組已經完成了A4B-17菌株的基因組測序和注釋[12]。合成4HBA的途徑有2條:葡萄糖降解途徑和莽草酸途徑,在后者途徑中磷酸烯醇式丙酮酸(PEP)和4-磷酸赤蘚糖縮合反應生成3-脫氧-D-阿拉伯庚酮糖酸-7-磷酸合成酶(DAHP),PEP的合成量較低成為莽草酸途徑的限速步驟。A4B-17菌株合成PEP的途徑有2條:2-酮-3-脫氧-6-磷酸葡糖酸途徑(ED途徑)和糖酵解途徑(EMP途徑)。在ED途徑中,磷酸烯醇式丙酮酸羧激酶(PEPCK)將草酰乙酸(OAA)脫羧生成PEP;在EMP途徑中烯醇化酶(Enolase)將2-磷酸甘油酸(OAA)脫水生成PEP。EMP途徑和ED途徑[13]的共同點是將合成PEP作為通路的最后一步反應。同時在之后的合成途徑中,PEP作為起始物質,4HBA及其酯類合成多少完全取決于它的積累量[14]。可以說要想大量合成4HBA及其酯類,就需要大量合成PEP。A4B-17菌株的PEPCK和Enolase這2個酶分別由編號為GM004356和GM001333的基因編碼,而且在該細菌體內,有且只有這2種酶能夠合成PEP,因此對這2個基因編碼的蛋白酶的研究是非常有必要的。對這2個酶的研究不僅能了解它們的理化性質,還能夠為提高PEP的產量提供理論基礎,從而為提高4HBA的產量打下基礎[15]。
1 材料與方法
1.1 試驗材料
Microbulbifer sp. A4B-17菌株(由筆者所在課題組從海水中分離得到,并保存于江蘇師范大學生命科學學院);大腸桿菌DH5α(筆者所在實驗室長期保存于 -80 ℃ 超低溫冰箱中);pET-21a(+)載體(購買自武漢淼靈生物科技有限公司)。
1.2 試驗試劑
Q5 High-Fidelity DNA Polymerase,5×Q5 High GC Enhancer(optional),5×Q5 Reaction Buffer,dNTP Mix,dd Water,LB肉湯,100 mg/mL Ampicillin(Amp)母液,50×TAE Buffer(pH值8.5),1%瓊脂糖凝膠,DNA marker,DNA loading buffer,GoldView核酸染料,質粒提取相關試劑[16],磷酸緩沖液PBS母液(0.2 mol/L),0.1 mol/L異丙基-β-D-硫代半乳糖苷(IPTG),30% (質量濃度)丙烯酰胺溶液(Acr-Bis),10%(質量濃度)過硫酸銨,5×Tris-Glycine Buffer(電泳緩沖液),1 mol/L Tris-HCl(pH值為6.8),1.5 mol/L Tris-HCl(pH值為8.8),10% SDS考馬斯亮藍R-250染色液,考馬斯亮藍脫色液,5×SDS-PAGE loading Buffer,β-巰基乙醇(β-ME),蛋白純化試劑盒等。
1.3 GM004356和GM001333編碼基因的克隆與鑒定
1.3.1 目的片段的擴增
首先分別設計GM004356和GM001333這2個目的基因的引物,引物設計見表1,引物序列交由上海捷瑞生物工程有限公司合成。設計PCR反應體系,加入PCR管中,混勻,放入PCR儀,設置好反應條件,運行程序。35次循環。
1.3.2 載體酶切
選擇pET-21a(+)載體,大小為5 443 bp,酶切位點為BamHⅠ和XhoⅠ,采用雙酶切,載體酶切后,需要將目的基因與載體進行連接,形成重組質粒。選擇的酶連試劑盒為Ready-to-Use Seamless Cloning Kit。總反應體系為 50 μL,混合均勻后于37 ℃水浴5 min。
1.3.3 陽性轉化子質粒提取
將重組質粒轉入大腸桿菌,培養結束后,挑取部分單菌落進行液體培養,然后利用十二烷基硫酸鈉(SDS)堿裂法提取質粒。
1.3.4 陽性轉化子酶切鑒定
將酶切體系加到PCR管中,混合均勻,放入37 ℃水浴鍋中孵育5~10 min,然后瓊脂糖凝膠電泳驗證結果。
1.4 PEPCK和Enolase的表達與酶活探究
1.4.1 粗酶提取
將活化好的菌落接入含氨芐青霉素(Amp)的LB三角瓶中,37 ℃,180 r/min,培養至吸光度為0.4~0.6,再加入異丙基-β-D-硫代半乳糖苷(IPTG)低溫低速誘導培養12~24 h,使目的蛋白充分表達。將培養好的菌液5 000 r/min,低溫離心10 min,棄上清,用pH值為7.4的PBS緩沖液(磷酸緩沖鹽溶液)重懸細胞,再離心,再重懸,用超聲波破碎儀破碎細胞,4 ℃ 離心,上清即為粗酶混合液。
1.4.2 蛋白純化
提取出的粗酶中,絕大部分是不需要的雜蛋白,想要研究目的蛋白的性質,需要將目的蛋白提取出來,選擇蛋白純化試劑盒進行純化。為了給后續試驗提供一個定量結果,需要計算出粗酶中PEPCK和Enolase的含量。采用BCA(聚氰基丙烯酸正丁酯)蛋白定量法,根據吸光度與蛋白濃度成正比,繪制蛋白濃度標準曲線,從而能夠計算出目的蛋白占粗酶的百分比。
1.4.3 PEPCK和Enolase酶活檢測方法
PEPCK和Enolase酶活檢測方法采用PK(丙酮酸激酶)/LDH(乳酸脫氫酶)偶聯檢測法,即將不方便檢測的磷酸烯醇式丙酮酸,經PK和LDH催化后,生成丙酮酸,然后與還原型輔酶Ⅱ(NADPH)反應生成NADP+,而NADPH容易被檢測,NADPH濃度的減少量與PEPCK和Enolase的活性成正相關,通過測定在340 nm處NADPH吸光度的減少,來反映酶的活性。
根據PK/LDH偶聯檢測法探究2個酶的最適溫度和最適pH值。研究PEPCK的最適溫度,要控制pH值、反應時間、調控劑不變。設置不同反應溫度,固定pH值不變;研究PEPCK最適pH值時,要控制溫度、反應時間、調控劑不變,各反應成分與研究最適溫度時一致,只是改變緩沖液的pH值不同。用同樣的方法來探究Enolase的最適溫度和最適pH值。
2 結果與分析
2.1 系統進化樹分析
在Microbulbifer sp. A4B-17菌株中找到編號為GM004356和GM001333的全部堿基序列,經NCBI數據庫比對分析,確定這2個基因分別編碼PEPCK和Enolase。GM004356編碼的PEPCK與Microbulbifer sp. GL-2、Microbulbifer sp. THAF38和Microbulbifer variabilis編碼的PEPCK相似度較高,相似度都為95.15%,系統進化樹見圖1。GL-2菌株分離自硬骨魚腸道中[17],是一個新的降解纖維素的菌株。Microbulbifer variabilis分離自太平洋海藻,能夠產生抗癌吩嗪抗生素[18]。關于THAF38菌株還沒有相關報道。GM001333編碼的Enolase與Microbulbifer variabilis和Microbulbifer sp. THAF38的Enolase相似度較高,相似度都為98.13%,系統進化樹見圖2。
2.2 高效表達質粒的構建
2個目的基因GM004356和GM001333分別進行PCR,瓊脂糖凝膠成像分析(圖3)。選用pET-21a(+)載體,采用雙酶切,與目的基因相連,構成重組質粒,導入大腸桿菌體內。通過驗證,獲得陽性轉化子,并測序,與原始目的基因比對,發現重組質粒中的目的基因未發生突變或缺失,與原始基因完全一致,可以利用該菌落進行后續的蛋白研究。
2.3 蛋白質的提取和純化
低溫低速誘導目的蛋白表達,提取粗酶,電泳初步判斷轉入大腸桿菌的目的基因成功表達,接著對粗酶進行純化,電泳驗證純化結果,發現目的蛋白PEPCK和Enolase均能被純化,且效果明顯,可以進行后續的定性分析,具體結果見圖4和圖5。
將粗酶液稀釋10倍后用BCA法測得樣品吸光度為2.526 4,通過標準曲線, 計算出粗酶液中蛋白總濃度為3.298 mg/mL。純化之后目的蛋白PEPCK吸光度為0.337,根據標準曲線,計算出純化后的PEPCK濃度為0.043 mg/mL,占粗酶液的1.304%。同樣的方法,得到Enolase占粗酶液的1.123%。
2.4 PEPCK和Enolase米式常數的研究
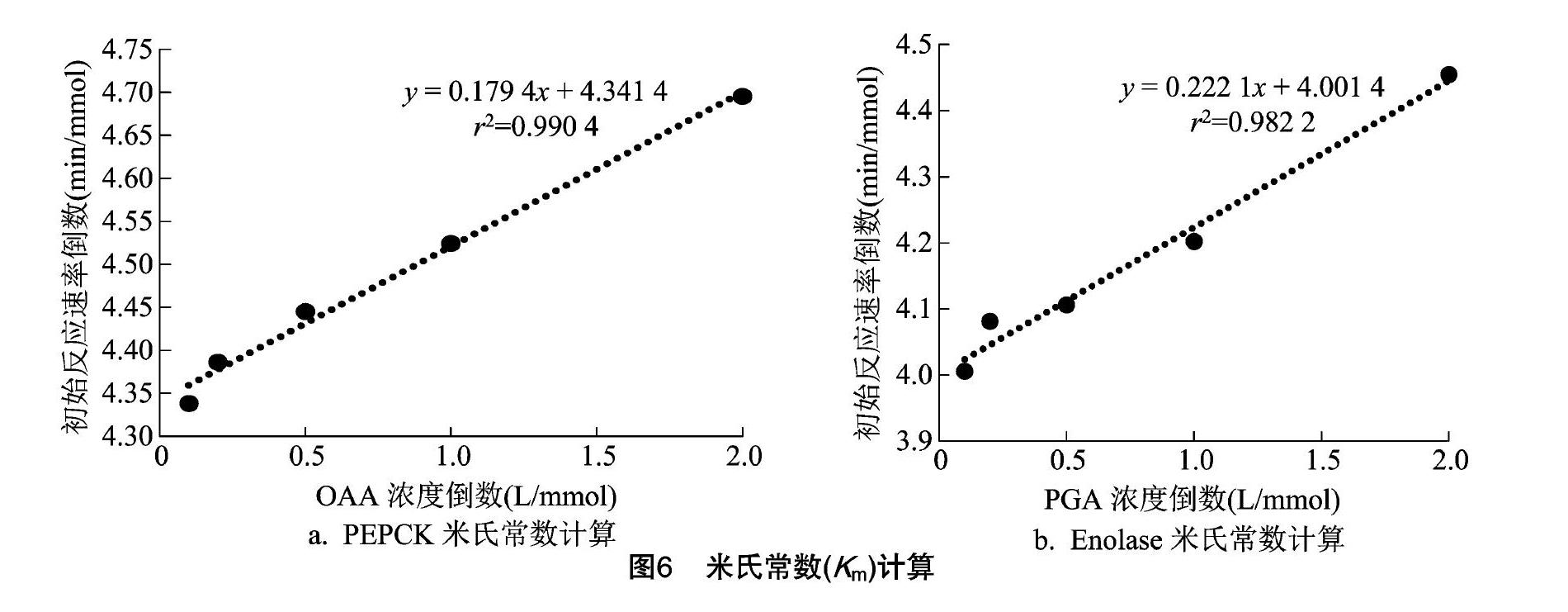
米氏常數(Km)定義為當酶促反應達最大速度(vm)一半時底物(S)的濃度。對Km的研究,能揭示其在參加反應時的重要性質,為該酶應用到工業生產中提供重要的理論依據。根據米氏常數雙倒數求法的計算公式:1/v=Km/(vmax[S])+1/vmax,可以看成y=ax+b的形式,需要做出初始反應速率的倒數,以及反應底物濃度的倒數,設定反應時間為 2 min,底物濃度足夠大。因為當y=0時,橫截距即為-1/Km,得到PEPCK的Km值為0.041 mmol/L,如圖6-A所示。Enolase的Km值為0.056 mmol/L,如圖6-B所示。2個Km值都較低,說明PEPCK與OAA,Enolase與底物2-磷酸甘油(PGA)的酶促反應很容易進行,從而轉化生產磷酸烯醇式丙酮酸(PEP),因此更有利于重要中間產物PEP在菌體中的積累。
2.5 最適溫度和最適pH值
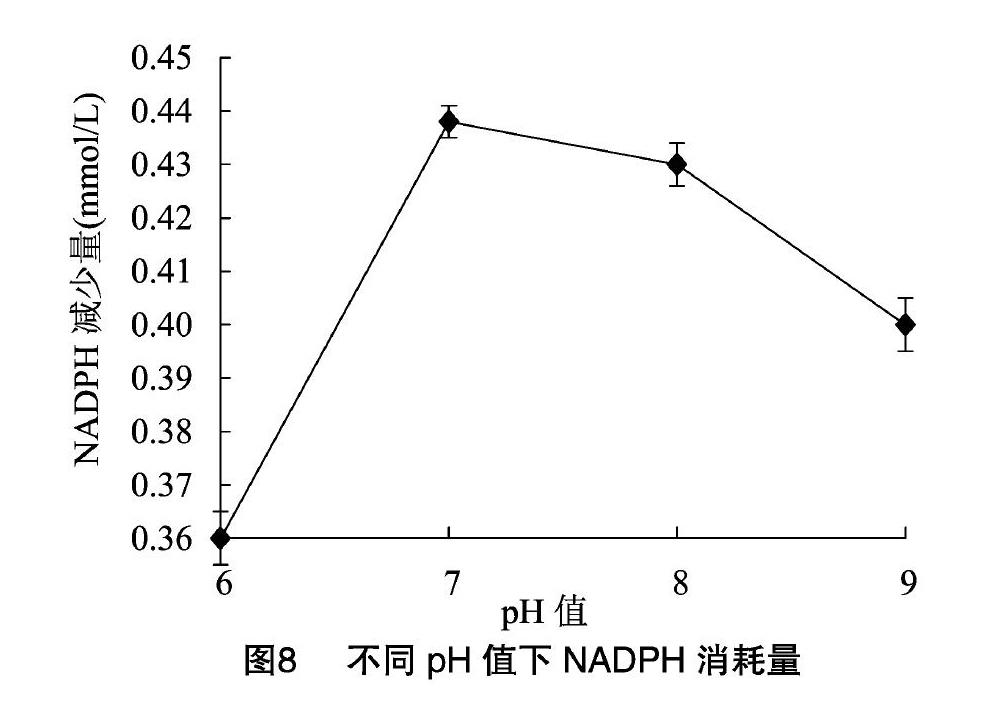
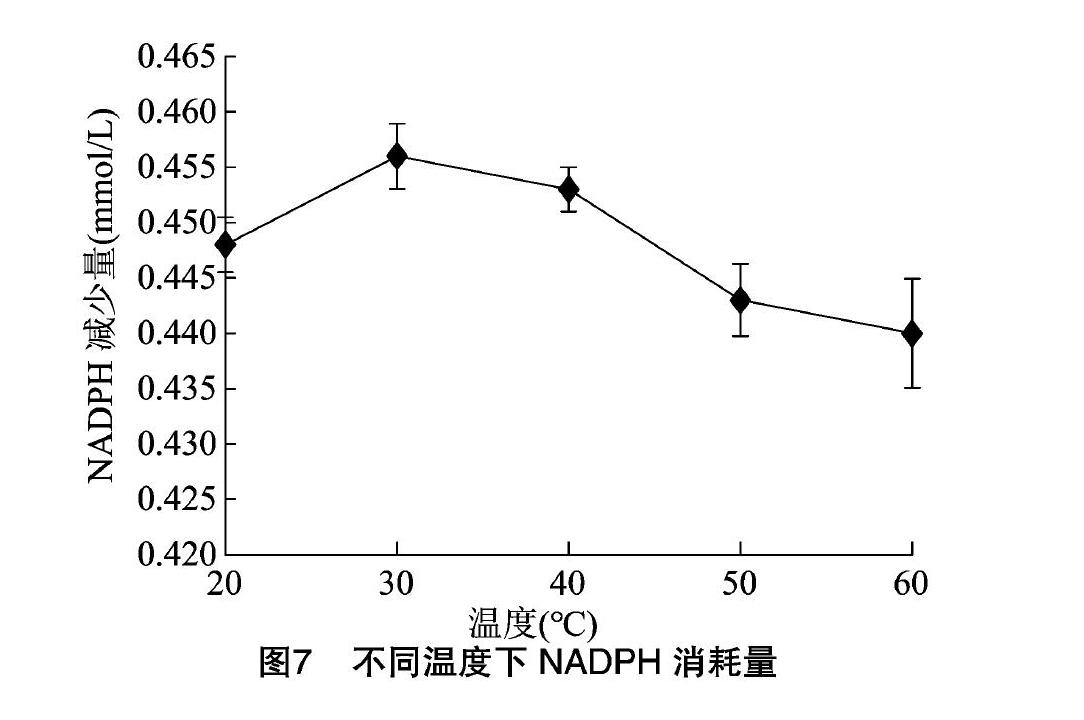
探究PEPCK的最適溫度,設置不同反應溫度,固定pH值保持不變,反應結束后測定NADPH的吸光度,結果見圖7,發現在30 ℃左右時,PEPCK活性最高。探究最適pH值時,設置不同pH值,保持溫度為30 ℃,發現pH值為7時酶活最高(圖8)。
同樣的方法,分別設置Enolase的反應溫度和緩沖液pH值,發現Enolase的最適反應溫度為30 ℃,最適pH值為7,結果如圖9和圖10所示。
3 討論與結論
Microbulbifer sp. A4B-17菌株的特點是本身就可以合成對羥基苯甲酸及其酯類,而且產量較高,不會產生對人體有毒有害的物質。本研究對細菌細胞內編號為GM004356和GM001333的2個目的基因進行轉化克隆,高效表達PEPCK和Enolase,通過提取純化計算出純化之后目的蛋白PEPCK占粗酶液的1.304%,純化后的Enolase占粗酶液的1.123%。PEPCK酶反應的最適溫度為30 ℃。最適pH值為7;Enolase的最適溫度為30 ℃,最適pH值為7。PEPCK和Enolase的米氏常數分別是0.041、0.056 mmol/L,較低的Km值更有利于酶促反應的快速進行。通過蛋白系統進化樹分析,PEPCK與Microbulbifer sp. GL-2的PEPCK相似度為95.15%;Enolase與Microbulbifer variabilis的Enolase相似度為98.13%。本研究對PEPCK和Enolase 2種酶進行深入研究,為將來2個酶的優化研究建立標準。筆者所在課題組期望通過對該菌株的深入研究,改變傳統的生產方式,利用可再生資源合成4HBA等工業原料。
本試驗所采用的酶活性研究方法均為PK/LDH酶偶聯法,該方法雖操作簡單,思路清晰,但是在偶聯法中,PK以及LDH的活性會對整個試驗結果產生較大影響,而且最終得到的米氏常數其實是目的蛋白酶與PK、LDH共同作用的結果。目前對PEP的檢測暫無有效可行的方法,開辟一種新的檢測方法還需要大量工作,后續可以在本研究的基礎上進行拓展研究。
參考文獻:
[1]郭 陽,臧 埔,郜 玉,等. 化妝品防腐劑的使用現狀及進展[J]. 中南藥學,2018,16(9):1258-1263.
[2]石金娥,劉靜秋,尚淑霞,等. 對羥基苯甲酸酯類防腐劑在醬油、食醋中應用狀況分析[J]. 中國調味品,2011,36(7):11-12.
[3]李欣航,張金龍,畢永賢,等. 無尼泊金酯類化妝品防腐體系的應用研究[J]. 日用化學品科學,2018,41(8):14-21.
[4]陳建文,厲華明,周榮榮. 食品中對羥基苯甲酸酯類的應用現狀與檢測方法[J]. 中國釀造,2008(8):4-5.
[5]胡 梅. 化妝品防腐及防腐體系的構建[C]//第九屆中國化妝品學術研討會論文集. 北京:品觀網,2012:105-107.
[6]朱 莉,許超艷,李晶晶,等. 生物合成對羥基苯甲酸的研究進展[J]. 生物工程學報,2015,31(3):328-337.
[7]林忠洋,馬萬里,齊 跡,等. 對羥基苯甲酸酯類防腐劑的人體暴露[J]. 化學進展,2015,27(5):614-622.
[8]Okai N,Miyoshi T,Takeshima Y,et al. Production of protocatechuic acid by Corynebacterium glutamicum expressing chorismate-pyruvate lyase from Escherichia coli[J]. Applied Microbiology and Biotechnology,2016,100(1):135-145.
[9]董秀梅,晁 青,王柏臣. 植物磷酸烯醇式丙酮酸羧激酶(PEPCK)研究進展[J]. 植物學報,2013,48(3):320-328.
[10]Zhang H R,Pereira B,Li Z J,et al. Engineering Escherichia coli coculture systems for the production of biochemical products[J]. Proceedings of the National Academy of Sciences of the United States of America,2015,112(27):8266-8271.
[11]Sommer S,Heide L. Expression of bacterial chorismate pyruvate-lyase in tobacco:evidence for the presence of chorismate in the plant cytosol[J]. Plant & Cell Physiology,1998,39(11):1240-1244.
[12]Tian J,Zhu L,Wang W J,et al. Genomic analysis of Microbulbifer sp. strain a4B-17 and the characterization of its metabolic pathways for 4-hydroxybenzoic acid synthesis[J]. Frontiers in Microbiology,2018,9:3115.
[13]張麗平,汪文君,李 智,等. 對羥基苯甲酸甲酯降解菌的初步研究[J]. 江蘇農業科學,2018,46(20):337-340.
[14]郭 晶,高菊芳,唐振華. 羧酸酯酶及其在含酯類化合物代謝中的作用[J]. 農藥,2007,46(6):365-368.
[15]Fiske M J,Whitaker R J,Jensen R A. Hidden overflow pathway to L-phenylalanine in Pseudomonas aeruginosa[J]. Journal of Bacteriology,1983,154(2):623-631.
[16]李載平. 分子克隆實驗指南[J]. 科學通報,2002,47(24):1888.
[17]Sugimoto Y,Ohnishi K I,Suzuki S. Complete genome sequence of cellulase-producing Microbulbifer sp. strain GL-2,isolated from marine fish intestine[J]. Microbiology Resource Announcements,2020,9(32):720-746.
[18]Nishijima M,Takadera T,Imamura N,et al. Microbulbifer variabilis sp. nov. and Microbulbifer epialgicus sp. nov.,isolated from Pacific Marine algae,possess a rod-coccus cell cycle in association with the growth phase[J]. International Journal of Systematic and Evolutionary Microbiology,2009,59(7):1696-1707.
