滇西北高原藏區小型獸類體表寄生蚤的生態位特征
2021-08-07 02:11:44劉正祥段興德邵宗體浦恩念杜春紅李玉瓊高子厚
生態學報 2021年13期
陳 星,劉正祥,段興德,邵宗體,浦恩念,蘇 超,杜春紅,李玉瓊,高子厚,*
1 中國鐵路成都局集團有限公司成都疾病預防控制中心,成都 610000 2 云南省地方病防治所,大理 671000
生態位指物種在生態系統中的位置,包括生境需求及其功能作用,既能反映該物種在各生態因子中的位置,又能反映其在物質循環與能量流動以及信息傳遞時所扮演的角色,一般主要運用生態位寬度和生態位重疊指數兩大指標來分析特定種群的生態位特征[1- 3]。生態位寬度可以解釋物種的豐度及其分布的特征與種間差異,生態位重疊則是度量物種間在生態因子聯系上的相似性。
寄生蚤作為蚤傳疾病、鼠傳疾病的主要傳播媒介,在鼠疫耶爾森菌等病菌的儲存及傳播過程中扮演重要角色,且多次從云南西北部高原藏區德欽縣鼠疫指示動物犬血清中檢測出F1抗體陽性,提示德欽縣可能存在鼠疫[4-5]。目前國內對植物、海洋生物、哺乳動物的生態學研究較多,小獸寄生蚤的生態學分析還存在很大的空白,因此明確德欽縣寄生蚤的生態學特征既對該地蚤傳疾病的防控至關重要,又是對該領域生態學研究的補充。生態位作為一個宏觀概念,在特定種群中,受多個生態因子的影響,故本文從海拔、生境和寄主3個資源軸對德欽縣小獸體表寄生蚤的生態位特征進行分析,了解不同生態因子中蚤種間分化、共存特征,為蚤傳疾病防控提供可靠生態學依據。
1 材料與方法
1.1 研究區域概況
德欽縣(98°3′56″—99°32′20″E,27°33′44″—29°15′2″N)位于云南省迪慶藏族自治州,地處滇西北橫斷山脈地區的滇川藏三省交界處,與鼠疫自然疫源地芒康縣、巴塘縣、德格縣、玉龍縣相鄰,為三江(瀾滄江、怒江、金沙江)并流核心區,是云南省地勢最高的縣城。境內最高海拔6740m,最低1464m,屬高山深谷地貌,立體氣候明顯,年均氣溫6.6℃,年均降水量631.7mm[6],干濕季分明,為蚤類等媒介生物的繁殖提供了有利條件[7]。2012年6月—2014年5月,依據海拔、生境等自然環境特征,在德欽縣境內選取42個調查點,進行群落生態學調查。
1.2 資源劃分
海拔范圍:2000—4500m,以500m為一個梯度,將其劃分為5個海拔梯度帶,依次以A—E表示;將生境劃分為4類:林地、耕地、灌叢、居民區,以I—IV表示。
1.3 樣本采集與處理
以鮮炸油條為誘餌,野外采用鋏夜法[8]布放鼠鋏,居民區采用籠日法[9]布放鼠籠。根據《中國動物志》和《中國脊椎動物紅色名錄》[10-11]依據外部特征對捕獲的小獸進行物種鑒定,梳撿其體表寄生蚤,保存于75%的乙醇溶液,依據《云南蚤類志》[12],在顯微鏡下進行形態學特征鑒定并記錄。
1.4 生態位指標測定
1.4.1生態位寬度測定
采用Shannon-Wiener 指數[13]來計算寄生蚤的生態位寬度,計算公式:
其中,Bi=i種的生態位寬度,Nij=i種利用j資源等級的數值,r=生態位的資源等級數。生態位寬度范圍:0—1,0表示沒有利用,1表示對所有等級都同樣的利用了。
根據李德志[14]等的研究,對生態位進行整合,將多個單一資源軸的生態位寬度值進行PCA(主成分分析)處理后整合成2個或單個新的資源軸:
其中,Bin為多維空間生態位寬度指數,Bi為第i個生態因子軸上的生態位寬度值,n為生態因子軸個數。
1.4.2生態位重疊指數測定
采用Pianka. E.R生態位重疊指數[15]公式來計算蚤的生態位重疊指數,計算公式:
其中,NOij=i種和j種之間的生態位重疊指數,Pik=物種i種在k資源等級的數量與物種i在所有資源等級的總數量之比,Pjk=物種j種在k資源等級的數量與物種j在所有資源等級的總數量之比。生態位重疊指數的變化范圍:0—1,0表示完全不重疊,1表示百分之百重疊,NO>0.6為有意義的重疊,說明重疊較為嚴重[16]。
1.4.3統計分析
各種指標計算均在EXCEL、SPSS 21.0中完成。
2 結果
2.1 寄生蚤構成及數量特征
本次調查共捕獲小獸1371只,隸屬4目7科18屬28種,共梳撿蚤1275頭,隸屬于4科9亞科17屬33種。按照徐正會[17]將物種構成比大于10%定義為優勢種,1%—10%位常見種,小于1%為稀有種的定義,本次調查結果中迪慶額蚤(Frontopsylla(Frontopsylla)diqingensis)、云南櫛眼蚤(Ctenophthalmus(Sinoctenophthalmus)yunnanus)為優勢種,分別占所撿蚤匹的25.02%、22.82%;特新蚤德欽亞種(Neopsyllaspecialisdechingensis)、特新蚤指名亞種(Neopsyllaspecialisspecialis)、相關新蚤指名亞種(Neopsyllaaffinisaffinis)、喜山二刺蚤川滇亞種(Peromyscopsyllahimalaicasichuanoyunnana)、結實茸足蚤(Geusibia(Geusibia)torosa)、棕形額蚤(Frontopsylla(Frontopsylla)spadixspadix)、絨鼠怪蚤(Paradoxopsyllacustodies)、直緣雙蚤德欽亞種(Amphipsyllatutadeqinensis)、無值大錐蚤(Macrostylophoraeuteles)、二刺大錐蚤(Macrostylophorabispiniformabispiniforma)、卷帶倍蚤(Amphaliusspirataeniusspirataenius)以及細鉤蓋蚤(Callopsylla(Callopsylla)sparsilissparsilis)為常見種;其余均為稀有種。在海拔選擇上,蚤匹主要分布在中高海拔帶,3500—4000m海拔帶撿蚤最多。在生境選擇上,林地撿蚤數量、種類最多(表1)。在寄主選擇上,優勢蚤種云南櫛眼蚤主要寄生于西南絨鼠(Eothenomyscustos),迪慶額蚤則以姬鼠屬中的大耳姬鼠(Apodemuslatronum)和中華姬鼠(Apodemusdraco)為主要寄主。所有寄主中,西南絨鼠體外寄生蚤數量最多,達383頭,其次為大耳姬鼠、藏鼠兔(Ochotonathibetana)和珀氏長吻松鼠(Dremomyspernyi);大耳姬鼠體外寄生蚤種類最多,達15種,其次為西南絨鼠、中華姬鼠、白尾松田鼠(Phaiomysleucurus)(表2)。
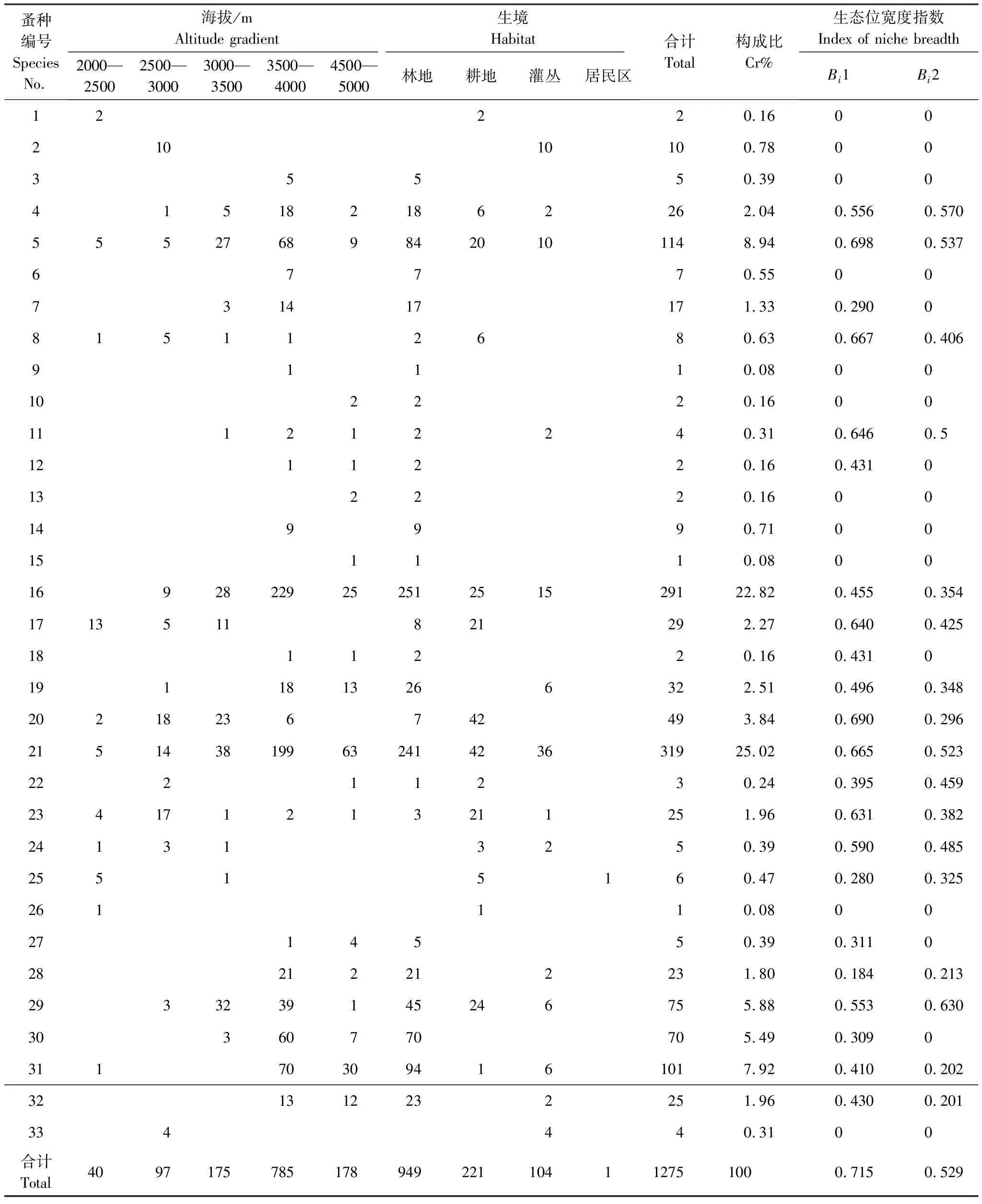
表1 德欽縣小獸體表寄生蚤群落結構及生境、海拔生態位寬度
2.2 寄生蚤生態位寬度指數
2.2.1海拔生態位寬度(Bi1)
在海拔資源軸上,總體生態位寬度值為0.715,蚤類生態位寬度范圍為0—0.698,其中生態位寬度值最大的是特新蚤德欽亞種,為0.698;其次是棕形額蚤、斯氏新蚤川滇亞種、迪慶額蚤、高山狹臀蚤(Stenischiamontanismontanis)、喜山二刺蚤川滇亞種和絨鼠怪蚤,生態位寬度值分別為0.690、0.667、0.665、0.646、0.640、0.631;而人蚤(Pulexirritans)等11種蚤類在該資源軸上的生態位寬度值為0,僅分布在單一海拔帶,未利用海拔等級資源(表1)。
2.2.2生境生態位寬度(Bi2)
在生境資源軸上,總體生態位寬度值為0.529,相對海拔資源軸的總體生態位寬度較窄。蚤類生態位寬度范圍:0—0.630,其中無值大錐蚤的生態位最寬,為0.630;其次是特新蚤指名亞種、特新蚤德欽亞種和迪慶額蚤,生態位寬度分別為0.570、0.537、0.523;而人蚤等16種蚤的生態位寬度為0,僅分布在單一生境,未利用生境等級資源(表1)。
2.2.3寄主生態位寬度(Bi3)
在寄主資源軸上,總體生態位寬度為0.749,是三種資源軸的總體生態位寬度中的最大值。蚤類的寄生生態位寬度值范圍為0—0.584,其中絨鼠怪蚤的生態位較寬,寬度值為0.584;其次是迪慶額蚤、棕形額蚤、喜山二刺蚤川滇亞種、特新蚤德欽亞種和斯氏新蚤川滇亞種(Neopsyllastevensisichuanyunnana),其生態位寬度值分別為:0.567、0.553、0.550、0.537和0.530;而鼠兔角頭蚤(Echidnophagaochotona)和圓凹多毛蚤(Hystrichopsylla(Hystroceras)zii)等9種蚤類的生態位寬度為0,僅寄生在單一宿主,未利用小獸等級資源(表2)。
2.2.4多維生態位寬度(Bin)
利用李德志對多個單維生態位寬度值整合的方法將三個生態因子進行整合,得到多維生態位寬度值Bin,為寄生蚤在海拔、生境、寄主3個資源軸上的整體生態位寬度。結果顯示多維生態位寬度值范圍為0—1.031,特新蚤德欽亞種和迪慶額蚤生態位寬度值最大,分別為1.031、1.019,在所有蚤種中占據相對最優的分布形式;其次為喜山二刺蚤川滇亞種、斯氏新蚤川滇亞種、絨鼠怪蚤、棕形額蚤,分別為0.945、0943、0.941、0.932,也顯示出較優分布。鼠兔角頭蚤、圓凹多毛蚤、后棘新蚤(Neopsyllahonorahonora)、紀氏叉蚤(Doratopsyllajii)、瀘水櫛眼蚤(Ctenophthalmus(Sinoctenophthalmus)lushuiensis)、長突怪蚤(Paradoxopsyllalongiprojectus)、昌都蓋蚤(Callopsylla(callopsylla)changduensis)7種蚤類生態位寬度值為0,在三種生態資源軸中均為單一分布形式(表2)。
2.3 蚤類生態位重疊指數
海拔生態位重疊指數中,0.6 生境生態位重疊指數中,有意義的重疊有208對,占比39.39%,多為特新蚤指名亞種、特新蚤德欽亞種、云南櫛眼蚤、迪慶額蚤、直緣雙蚤、卷帶倍蚤、細溝蓋蚤與其他蚤種的重疊。NO=0的有98對,占比18.56%,多為稀有種之間的重疊,最多的是與昌都蓋蚤的重疊,其次是與鼠兔角頭蚤、介中怪蚤(Paradoxopsyllaintermedius)、金沙江怪蚤(Paradoxopsyllajinshajiangensisjinshajiangjinshajiangensis)和長突怪蚤的重疊;NO=1的有70對,占比13.26%,最多的是與圓凹多毛蚤的重疊,其次是二刺大錐蚤、直緣雙蚤察里亞種(Amphipsyllatutachaliensis)和二毫新蚤指名亞種(Neopsyllabisetabiseta)的重疊,除相關新蚤指名亞種和二次大錐蚤為常見種外,其他均為稀有種。(表3) 寄主生態位重疊指數中,有意義的重疊有49對,占比9.28%,NO=0的有270對,占比51.14%,最多的是與昌都蓋蚤的種對,其次是與鼠兔角頭蚤和二刺大錐蚤的種對;NO=1的僅有4個種對,占比0.19%,分別是鼠兔角頭蚤和昌都蓋蚤、圓凹多毛蚤和紀氏叉蚤、瀘水櫛眼蚤和云南櫛眼蚤、云南茸足蚤(Geusibia(Geusibia)yunnanensis)和卷帶倍蚤的生態位重疊。(表4) 生態位(Niche)理論作為探討群落演變的經典理論,在種間關系、群落結構、物種多樣性及種群進化等研究領域被廣泛應用[18]。生態位寬度和種間生態位重疊被認為是物種多樣性及群落結構的決定因素,也反映了所在群落的穩定性[19]。 本次調查結果顯示,迪慶額蚤和云南櫛眼蚤為優勢種,但在三種資源軸中的生態位寬度值均不是最大,這是因為生態位寬度值除了與數量有關之外,更重要的是與分布有關,某些具有數量優勢的蚤種在分布中并不具有分配優勢,如云南櫛眼蚤98.3%寄生于單一宿主西南絨鼠。三種資源軸中,具有較大生態位寬度值的寄生蚤均為常見蚤種,如特新蚤德欽亞種,棕形額蚤等,這些蚤種除了在數量上具有相對優勢外,在資源軸內部分布也具有優勢,為廣生態位物種,跨境分布廣,生態幅寬,數量相對均勻,具有很強的競爭優勢。而生態位寬度值較小甚至為0的蚤種,如人蚤、鼠兔角頭蚤等蚤類,僅在單一生境、海拔、寄主中有分布,為狹生態位物種,生態幅窄,適應能力弱,未利用等級資源,在資源競爭中不具備競爭優勢,生態脆性增加[20],容易因外部環境變化導致物種滅絕。 生境因子、海拔因子與資源的利用、種間種內的競爭、捕食與寄生、及生物的繁衍等息息相關[21-22]。本次調查中,小獸體表寄生蚤生境選擇以灌木叢為主,這可能是因為該生境中水和食物資源充足,環境隱蔽,適宜小獸生存,而寄生蚤的分布很大程度上依賴于小獸的分布[23]。在海拔選擇上,集中在3500m以上的中、高海拔帶,多為云南櫛眼蚤和迪慶額蚤,這與其主要宿主大耳姬鼠、西南絨鼠分布一致,也是依賴小獸分布的另一種表現形式。在寄主資源軸上絨鼠怪蚤、棕形額蚤、迪慶額蚤占優勢,分別寄生于7、9、12種小獸,是具有最多寄主的蚤種。而在寄主資源軸上,蚤的生態位寬度值越大,其生態幅越寬,對寄主選擇的特異性越低,可寄生的宿主越多,越傾向于是一個泛化種。生態位越窄,對寄主選擇性越高,特化程度越高,更傾向于是一個特化種,甚至專一寄生。該資源軸與寄生蚤本身的特性,如寄生方式、對寄主的選擇性等高度相關。多維生態位寬度顯示特新蚤德欽亞種和迪慶額蚤生態位寬度值最大,而特新蚤作為云南省齊氏姬鼠-大絨鼠鼠疫疫源地的媒介蚤,其在鼠疫傳播中發揮著重要作用,應重點關注,加強防控。 生態位重疊指數反映的不僅是不同蚤種對于資源利用的相似度,更表明了潛在的競爭程度,重疊指數越高,其選擇越近似[24]。以海拔和生境為資源軸計算的生態位重疊指數顯示:完全重疊和完全不重疊的種對,多為稀有種與稀有種之間,而有意義的重疊多為優勢種、常見種、稀有種兩兩之間交叉重疊。這是因為優勢蚤種和常見蚤種依賴于小獸的分布,除了具有數量優勢外,還具有分布優勢,較寬的生態幅和較強的跨帶分布能力讓其競爭優勢明顯,因此和其他蚤種重疊指數較高。而稀有種數量較少,寄主相對單一,甚至只寄生于某一種小獸,導致選擇同一小獸作為寄主的蚤種之間重疊指數高,甚至完全重疊,而選擇不同小獸作為寄主的蚤種間重疊指數低,甚至完全不重疊。在以寄主為資源軸的重疊指數表明:完全重疊的種對很少,完全不重疊的種對很多,更進一步表明蚤類寄生方式、對宿主的差異性選擇對其生態位重疊指數的的影響較大。 重疊指數較高甚至完全重疊的物種能夠在同一資源軸上同時存在是因為在其他資源軸上存在性狀替代或分離,如取食時間不同、活動時間不同、食性偏好不同[25-27]等。這種通過生態位分化減少種間競爭,更合理的利用生態資源,實現共存而沒有導致相互驅離的現象[28]在物種間普遍存在。某一蚤種能與其他多個蚤種重疊,說明其競爭能力強,在生態競爭中更容易存活,在外力導致群落重構過程中,具有更大的競爭優勢[29]。這種不同物種的適應性差異構成的群落物種的生態位分化,是不同物種對環境適應性的結果[30]。隨著資源軸的增加,物種的生態位分化會逐漸增大,以達到共存[31]。 在任一生態系統中,物種間的相互作用都始終處于動態變化中,通過不同功能生物間激烈的競爭或共生驅動形成不同資源軸上的生態位分化,減少種間競爭,進而實現共存,形成相對穩定的群落結構[29]。但當環境中原有優勢種因外部原因突然銳減,其占位種會快速占據優勢地位,成為新的優勢種,如果其為重要的疾病傳播媒介蚤種時,可以導致疾病的爆發甚至快速流行。因此,相對穩定的蚤類群落結構和其生態位格局對于蚤傳疾病的防控相當重要。 本文運用生態位寬度值和生態位重疊指數分析了德欽縣小型獸類體表寄生蚤的生態學特征。結果表明不同資源軸上寄生蚤的生態位寬度值不同,重疊指數也不同,這除了與調查環境本身的海拔、生境相關外,還與寄生蚤的寄生方式和對寄主的差異性選擇有關。該結果揭示了寄生蚤在不同生態因子中的生態位格局,為鼠蚤監測提供指導,對于有效防控蚤傳疾病具有重大現實意義。 致謝:云南省地方病防治所和德欽縣疾病預防控制中心幫助調查,特此致謝。3 討論


4 結論
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
吉林廣播電視大學學報(2021年4期)2022-01-14 02:35:48
保健醫苑(2021年7期)2021-08-13 08:48:02
作文成功之路·小學版(2020年5期)2020-06-11 12:48:26
學生天地(2020年36期)2020-06-09 03:12:30
小學科學(學生版)(2020年5期)2020-05-25 07:11:32
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
小天使·一年級語數英綜合(2018年11期)2018-11-23 09:47:26
汽車觀察(2018年10期)2018-11-06 07:05:26
資源再生(2017年3期)2017-06-01 12:20:59
