長期增溫對樹線交錯帶岷江冷杉幼苗異齡葉大小與出葉強度關系的影響
2021-09-01 10:55:18曾郅瑋趙世杰鮮駿仁陳國鵬王玉熒
生態學報 2021年14期
關鍵詞:生長
曾郅瑋,趙世杰,鮮駿仁,*,陳國鵬,王玉熒,秦 慧,馬 俊,符 鑫
1 四川農業大學環境學院, 成都 611130 2 多倫縣草原工作站, 錫林郭勒盟 027300 3 甘肅農業大學林學院, 蘭州 730070
為高效利用從外界獲得的資源, 植物個體各組成部分將以不同速率生長來呈現出相應的異速生長規律[1]。因此, 植物各組成部分的數量和大小之間的比例關系能夠反應個體發育過程中資源的分配策略, 并可通過冪指數異速生長機制模型進行研究[2]。葉片是植物進行光合作用的主要場所, 其生長狀況與形態結構決定了植物轉化光能的效率[3];在資源受限時, 植物的葉片大小與數量間存在著權衡關系[4]。小枝是葉片的載體, 其機械結構影響葉的大小與分布[5- 6], 同時小枝與葉之間存在資源分配策略, 枝葉大小關系被認為是植物生活史策略研究的一個重要維度[4]。因此, 葉的大小與數量的權衡與小枝的屬性密切相關。
Kleiman和Aarssen[7]以單位體積小枝上的葉片數量表征出葉強度, 探究單葉質量與出葉強度間的協變關系, 并認為這兩者間存在恒定的負等速生長, 提出了“出葉強度優勢”假說。越來越多的研究表明不同生活型、不同生境的植物葉大小-出葉強度間存在等速的權衡關系[8- 9], 但也有一些研究表明此關系并非均為等速: 劉長柱等[10]的研究表明在不同森林類型與森林群落間, 單葉質量-出葉強度的權衡關系存在差異;Fajardo[11]也發現南青岡(Nothofaguspumilio)的單葉面積-出葉強度間的斜率與-1.0有顯著區別。目前研究對于葉大小和出葉強度的定義并不一致,這必然影響對權衡關系普遍性的探討。葉大小一般使用單葉質量或單葉面積表征, 代謝生態學理論中, Price等對植物分型網絡模型(由West、Brown和Enquist提出, 簡稱WBE模型)進行了擴展, 并假設葉面積與葉生物量呈等速生長關系[1,12- 13], Niklas等提出的“收益遞減”假說則認為葉面積與葉生物量的異速生長指數顯著小于1.0[14- 15]。因此, 使用單葉質量與單葉面積表征葉大小可能使結果存在差異。出葉強度基于生物量還是體積因研究對象而異, Kleiman和Aarssen采用單位體積莖的葉數計算出葉強度, 被認為未考慮不同樹種的木材密度[7,16];而Huang等[17]認為單葉質量與基于體積的出葉強度間的權衡取決于生物量投資與莖組織密度,而與基于質量的出葉強度間的權衡僅取決于生物量投資。對不同參數表征葉大小與出葉強度的差異進行比較, 可以明確權衡關系的判斷是否受定義影響, 促使一致定義的形成, 有助于探討此權衡關系的普遍性。
個體發育過程與其所處環境密切相關, 在全球變暖的影響下, 植物的個體形態、生長發育過程都會發生明顯的變化[18- 20]。環境溫度的升高將使個體的代謝速率提高[21- 22], 不僅引起葉片大小的改變, 還將影響枝葉的生長關系[23- 24], 葉大小-出葉強度的權衡關系可能因此發生變化。同時, 植物個體的生長是一個長期的過程, 常綠樹種多年生葉片和枝條對植物的生活史策略具有重要影響。然而, 為避開枝條次生生長對研究的影響,以往的研究多數只利用當年生的小枝開展枝葉關系分析[10,25- 26], 對多年生枝條的枝葉關系研究較少;確定當年生小枝上所發現的權衡關系是否可以推及多年生枝條, 有助于進一步探討植物功能性狀關系隨年齡的協變規律[27]。不同年齡的葉片的物質分配與環境敏感性已被證實存在差異[23,28],那么在植物生長過程中, 葉大小與出葉強度的關系是否存在變化, 全球變暖如何影響葉大小與出葉強度的關系。對這些問題進行探究, 能夠解釋全球變暖模式下植物不同發育階段的葉大小-出葉強度的權衡變化。
川西亞高山樹線交錯帶地處青藏高原東部, 因相對簡單的群落結構和樹線附近的邊緣效應而成為了研究高寒生態系統對全球變化響應的天然平臺[29], 而岷江冷杉(Abiesfaxoniana)是川西北亞高山針葉林的建群種之一。前期研究表明增溫將使岷江冷杉芽提前開放, 也會限制葉面積的擴大而促進葉片厚度的增加[23,29], 但并未涉及葉大小與出葉強度的權衡關系研究。因此, 本研究以岷江冷杉為研究對象, 利用前期設置的開頂箱(Open-top chamber, OTC)模擬全球變暖, 以單葉質量、單葉面積表示葉大小, 分別以莖生物量、莖體積為載體計算出葉強度, 對葉大小-出葉強度進行了異速生長分析, 研究長期增溫對岷江冷杉葉大小與出葉強度關系的影響, 擬回答以下科學問題:使用不同參數表征葉大小與出葉強度的異同?葉大小-出葉強度權衡關系是否存在于多年生小枝中?長期增溫對葉大小-出葉強度權衡關系具有何種影響?
1 材料和方法
1.1 研究區概況
本研究位于四川省平武縣王朗國家級自然保護區(32°59′ N, 104°01′ E, 海拔2300—4980 m):地處青藏高原東緣, 屬山地季風性氣候, 年平均氣溫1.5—2.9℃, 7月平均氣溫12.7℃, 1月平均氣溫-6.1℃。年降水量在801—825 mm之間, 雨季多集中在5月到8月。岷江冷杉林(2600—3500 m)是保護區主要森林群系, 土壤為暗棕壤。
1.2 樣地設置
實驗設置已有詳盡描述[23,30]:2005年9月,在大窩凼外側坡樹線交錯帶的平緩灌叢設置8個1.5 m×1.5 m×2.6 m、頂端開口面積1.2 m×1.2 m的OTC,OTC由聚碳酸酯制作,在OTC附近設置對照樣方(CK),每個OTC和對照樣方間距在5 m以上;10月從樣地附近移植長勢良好岷江冷杉(12—15 a,高約50—60 cm)和糙皮樺(Betulautilis,5—6 a,高約50—60 cm)幼苗到OTC及CK內,幼苗等距相間40 cm呈“田”排列(4或5株),幼苗距OTC均30 cm,移植中不破壞原有植物;2006年生長季,定期對OTC及CK內物種澆水,以確保物種存活;在生長季對OTC及CK的監測表明:OTC中空氣溫度(1.2 m)平均較CK高2.9℃,土壤溫度(5 cm)增加0.4℃。增溫全年進行,試驗地周圍用網圍欄封育,避免動物干擾,OTC和CK均不再人工施肥和澆水,OTC外膜保持潔凈,期間不定期對OTC裝置進行維護;2009年項目完成后未繼續監測環境因子, 僅對OTC進行維護。
1.3 植物采樣與測量
本實驗的研究對象為4年生一級枝條上當年 1年生、2年生和3年生的小枝, 小枝包括莖及其上著生的葉(4年生及以上的枝條有葉片脫落)。2016年8月下旬, 自幼苗梢部向下的4年生一級側上隨機采集小枝, 用枝剪從OTC及對照樣地內逐株個體的不同方向采集生長良好、葉片完整的枝條(目標枝均有二級枝、無三級枝), 根據枝條上芽鱗痕形成的環節判斷枝段年齡, 每個OTC、CK內各齡級保證4個有效樣本; 分別裝入自封袋記錄編號帶回實驗室, OTC和CK內各齡級保證30個有效樣本。將小枝上的完整葉片與莖分開并記錄小枝上的葉數, 用數顯卡尺(±0.01 mm)測量莖長度與直徑(中部), 使用葉面積儀(CI- 203, CID, USA)測定單葉面積。莖和葉片于105℃下殺青15 min后于85℃烘至恒量并稱量。
1.4 數據處理
將直接測量所獲得的數據錄入Excel表格中并進行計算:單葉質量(ILM)由葉生物量(LM)和葉數量(LN)計算得到(ILM=LM/LN);單葉面積由葉面積儀(CI- 203, CID, USA)直接測定;出葉強度分別基于莖生物量與莖體積, 即單位莖生物量上的葉片數量與單位莖體積上的葉片數量, 其中, 莖體積(SV)由莖長度(SL)與莖直徑(SD)計算得到(SV=(SD/2)2·π·SL)。進行異速生長分析前, 將各參數進行以10為底的對數轉換, 使之符合正態分布。所收集到的數據小枝屬性包括:葉生物量、葉片數量、單葉質量、單葉面積、莖生物量、莖直徑、莖長度、莖體積、基于莖生物量的出葉強度、基于莖體積的出葉強度。
葉大小與出葉強度間的權衡關系通過異速生長方程y=bxa來描述, 并轉換為log(y)=log(b)+a×log(x), 其中x與y分別表示出葉強度和葉大小, 斜率a表示異速生長指數(a=±1為等速生長,a≠±1為異速生長), 截距b表示異速生長常數。使用軟件SMATR(Standardised Major Axis Tests & Routines, v2.0)采用標準化主軸估計(SMA)方法[31-32]計算葉大小和數量間的異速生長指數和常數, 檢驗不同處理組間SMA斜率a的異質性,當斜率同質時給出共同斜率并檢驗y軸截距和SMA斜率方向上的變化,并進行多重比較(先對六組數據進行斜率異質性檢驗, 若整體上不存在共同斜率, 再分為對照組和升溫組, 進行各年齡段的斜率異質性檢驗)。小枝性狀的統計分析在SPSS 25.0 (SPSS Inc., Chicago, IL, USA) 上進行,顯著性為P<0.05。
2 結果分析
2.1 單葉質量與出葉強度的關系
岷江冷杉小枝單葉質量和出葉強度間存在顯著的負相關關系(圖1; 表1), 增溫和年齡未顯著影響異速生長關系的斜率。當出葉強度基于莖生物量時(圖1), 各組斜率與-1.0無顯著差異(表1), 具有共同斜率-1.018(P=0.415, 95%置信區間為-1.138到-0.909), 與-1.0無顯著差異(P=0.760)。在共同斜率下, 各齡級間截距隨小枝年齡的增大而減小(表1)。增溫對共有斜率下1年生組的截距影響極顯著差異(P<0.01), 對2年生組與3年生組的截距影響不顯著(P值分別為0.773、0.140), 即在相同出葉強度下, OTC中1年生小枝相較對照具有更小的單葉質量, 或相同單葉質量下, OTC中1年生小枝的出葉強度更低。
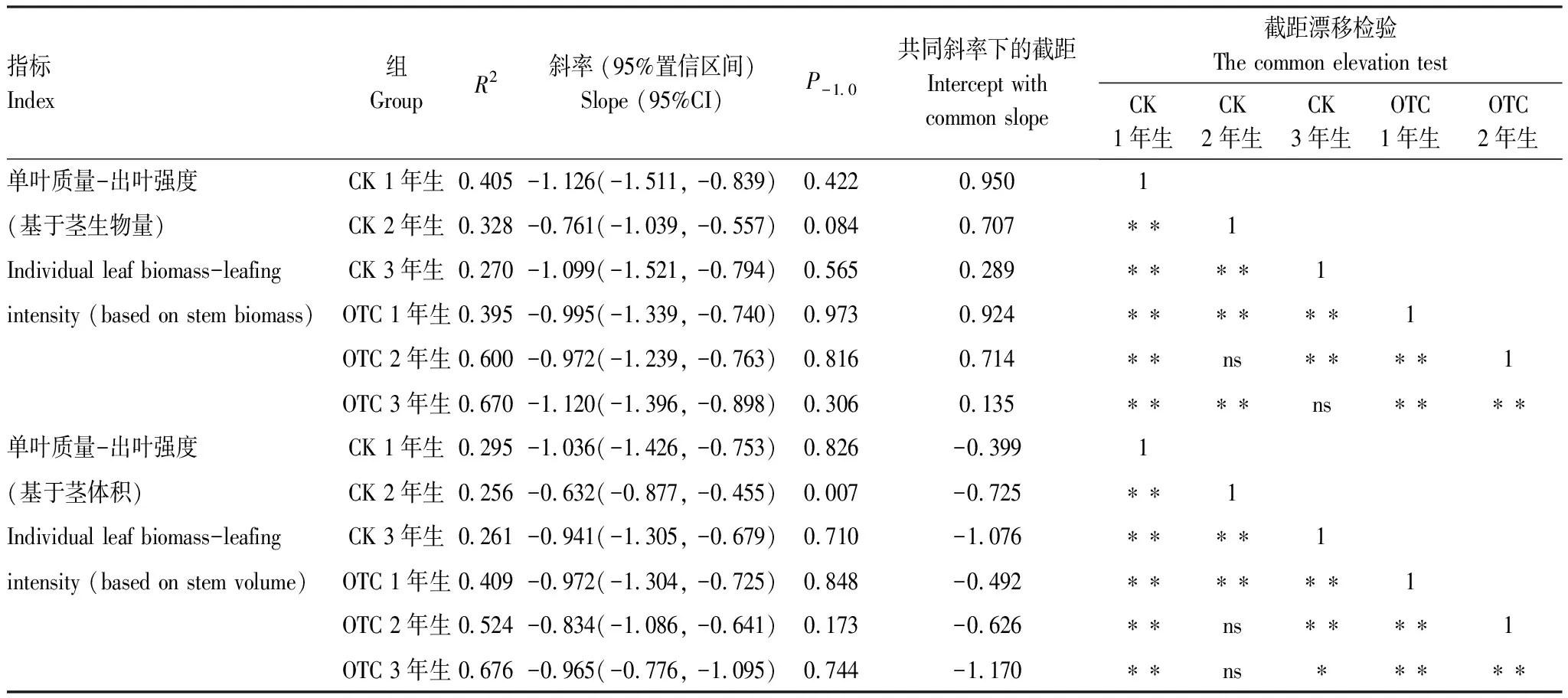
表1 岷江冷杉異齡葉單葉質量與出葉強度的SMA分析
當出葉強度基于莖體積時(圖1), 除對照2年生小枝外, 各組斜率與-1.0無顯著差異(表1), 且共同斜率-0.900(P=0.276, 95%置信區間為-1.011到-0.801)與-1.0無顯著差異(P=0.075), 但與基于莖生物量時的共同斜率存在顯著差異(P<0.05)。在共同斜率下, 截距在齡級間的變化與基于莖生物量時一致, 當小枝年齡相同時, 對照1年生小枝極顯著大于升溫(P<0.01), 對照2年生小枝與OTC無顯著差異(P=0.173), 對照3年生小枝顯著大于OTC(P<0.05)。無論出葉強度基于莖生物量還是莖體積, 六組數據的共同斜率與-1.0均無顯著差異, 單葉質量與出葉強度間存在恒定的負等速生長關系, 截距隨小枝年齡增加而減小; 當年生小枝的截距受增溫影響而顯著減小, 其共同主軸向下漂移。

圖1 岷江冷杉異齡葉單葉質量與出葉強度之間的關系Fig.1 Relationships between individual leaf biomass and leafing intensity at different ages of A. faxonianaCK和OTC分別表示對照(control check)和開頂箱(Open-top chamber)
2.2 單葉面積與出葉強度的關系
岷江冷杉小枝單葉面積大小與出葉強度間存在顯著負相關關系(圖2,表2)。當出葉強度基于莖生物量時(圖2), 整體上并不存在共同斜率(P=0.008)。相同年齡小枝對照組與增溫組的斜率間無顯著差異(P值均大于0.05), 增溫并未造成異速生長關系斜率的改變。在各齡級間, 對照2年生小枝斜率的絕對值極顯著小于對照1年生和3年生小枝, 異速生長關系發生改變;升溫組中存在共同斜率-0.740 (P=0.450, 95%置信區間為-0.879到-0.623), 極顯著小于-1.0 (P<0.01), 即出葉強度變化時單葉面積并非等比改變。
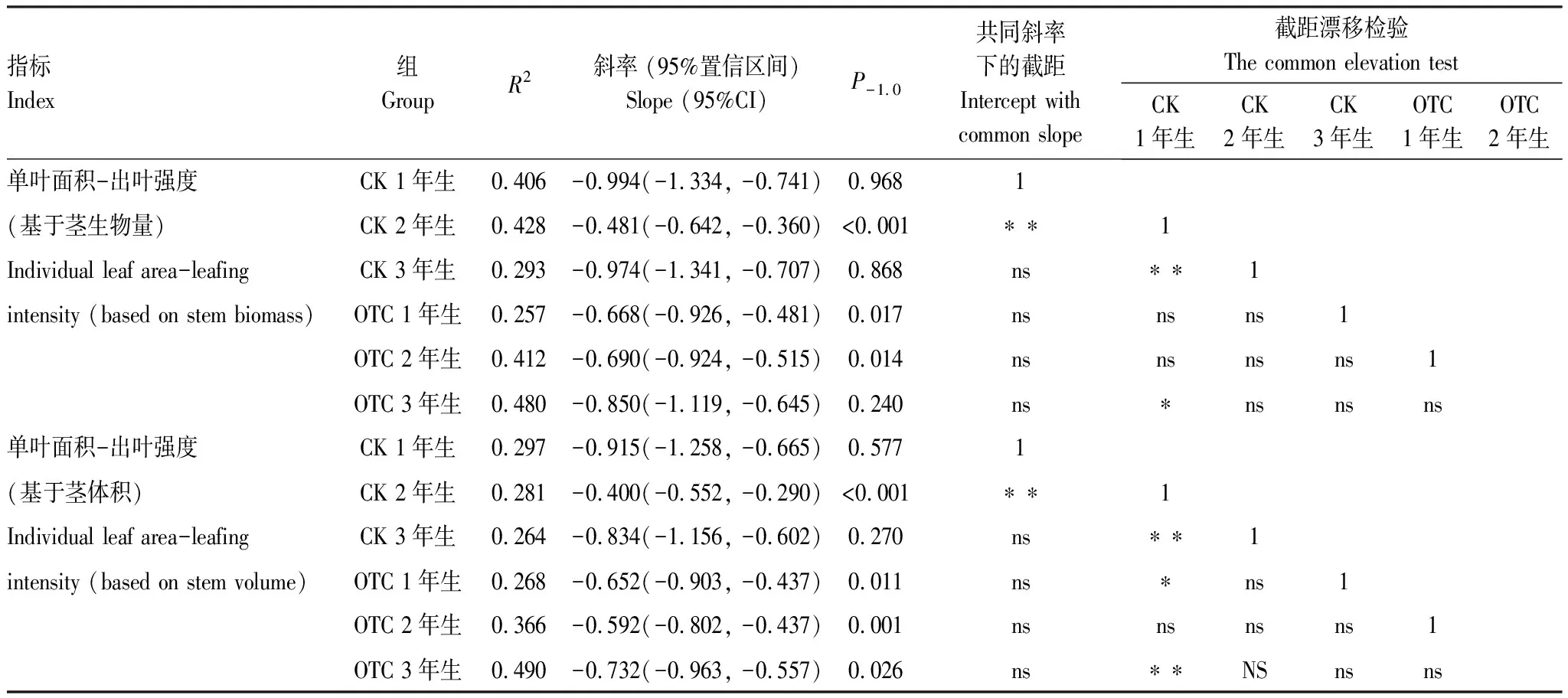
表2 岷江冷杉異齡葉單葉面積與出葉強度的SMA分析
當出葉強度基于莖體積時(圖2), 整體上不存在共同斜率(P=0.01), 同齡級間對照與增溫斜率無顯著差異, 各齡級間, 對照2年生斜率絕對值顯著小于1年生與3年生(P值均為0.001), 在增溫組中存在共同斜率-0.662(P=0.570, 95%置信區間為-0.786到-0.557), 與-1.0存在顯著差異(P<0.001)。這些結果與基于莖生物量的結果一致。
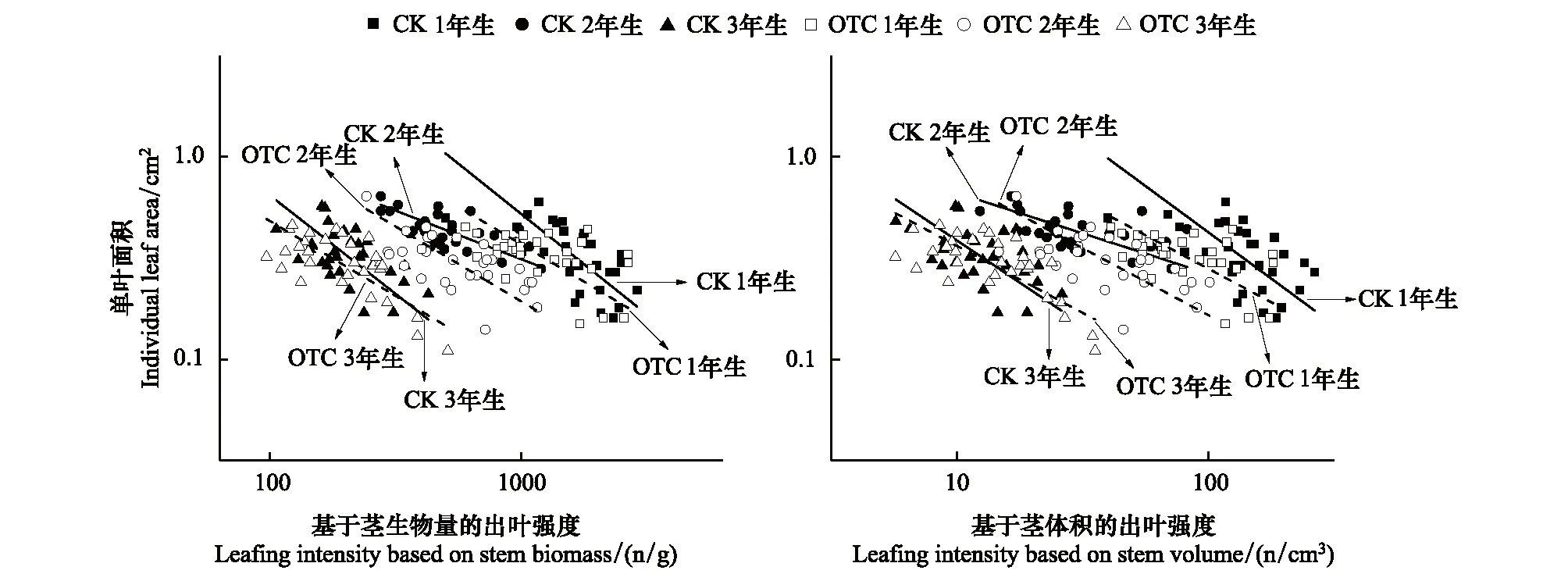
圖2 岷江冷杉異齡葉單葉面積與出葉強度之間的關系Fig.2 Relationships between individual leaf area and leafing intensity at different ages of A. faxoniana
3 討論
3.1 基于參數表征導致的差異
當用單葉質量表示葉大小時, 各組具有共同斜率且與-1.0無顯著差異(出葉強度分別基于莖生物量、莖體積,P分別為0.415、0.276), 而用單葉面積表征時, 對照組間的斜率具有顯著差異, 升溫組中存在的共同斜率顯著大于-1.0。這表明葉大小與數量的權衡關系會因為葉大小的表征方式不同而存在差異, 符合收益遞減假說[14- 15], 即隨著葉生物量的增加, 葉面積的增加量小于葉生物量的增加量, 葉面積與葉生物量間存在斜率顯著小于1.0的異速生長關系, 因此, 當單葉質量與出葉強度間存在恒定的負等速生長關系時, 可以推斷單葉面積與出葉強度間存在斜率顯著大于-1.0的異速生長關系。這種現象與跨物種的研究有別, 如楊冬梅等跨61個物種的研究發現, 不同物種葉面積和葉質量收益遞減的差異可能是造成岷江冷杉單物種葉大小與數量權衡有別于跨物種關系的關鍵[33]。此外, 以單葉面積表征葉大小時, 2年生小枝組的權衡關系發生了顯著或極顯著的改變(與1年生組和3年生組的斜率差異, 圖2), 這可能與年際間的溫度與降水變化有關[23]。前期研究中(2009年項目結束而未進行溫度的連續監測), 2007年OTC增溫2.2℃且降水較多, 促進了綿穗柳(Salixeriostachya)單葉面積的增加, 而2009年OTC增溫2.9℃而降水減少, 綿穗柳葉面積顯著降低[29,34]。與單葉質量相比, 單葉面積僅是一個二維變量, 并未考慮到葉片厚度的變化, 將來的研究中可考慮結合葉片厚度進行比較, 在此基礎上分析葉面積、體積等之間是否存在等速/異速生長關系, 以全面揭示這些性狀間的權衡。
使用單葉質量表征葉大小時, 基于莖生物量的共同斜率-1.018顯著小于基于莖體積的共同斜率-0.900, 但兩者共同斜率都與-1.0無顯著差異, 均符合Kleiman和Aarssen的“出葉強度”假說[7]; 單葉面積年際差異較大, 在表征葉大小時權衡關系發生了變化, 但基于莖生物量和莖體積的出葉強度的變化趨勢是一致的。這些結果符合Huang等的推測[17], 但又存在一定差異。Huang等[17]通過公式推導后認為基于質量的出葉強度間的權衡僅取決于生物量投資, 而基于體積則取決于生物量投資與莖組織密度,并推測兩者存在差異; 而在本研究中, 以單葉質量表征葉大小為例, 一方面, 基于莖生物量的共同斜率顯著小于基于莖體積的共同斜率, 兩者存在差異; 另一方面, 兩者的共同斜率均與-1.0無顯著差異, 在判斷是否為負等速權衡時結果一致。值得注意的是, 僅在判斷對照2年生的權衡關系時, 基于莖生物量和莖體積的結果是不同的(以單葉質量表征葉大小; 前者P-1.0= 0.084, 后者P-1.0= 0.007; 表1), 原因可能是兩種參數下的斜率差約0.1, 這一斜率差使顯著性的判斷不同。本研究結果表明基于莖生物量的斜率小于莖體積(斜率差約為0.1), 但一般情況下兩者對權衡關系的判定結果相同。就本研究而言, 單葉質量與基于莖生物量的出葉強度間存在恒定的負等速生長關系, 此關系并不隨小枝年齡而改變, 亦不受模擬增溫的影響, 可更好地體現葉大小與出葉強度的異速關系共享一個權度指數(Scaling exponent)。
3.2 葉大小-數量權衡關系隨小枝年齡的變化
在本研究中, 當用單葉質量表征葉大小時(出葉強度基于莖生物量, 下同), 無論是對照組還是增溫組, 異速生長指數在齡級間均無顯著差異; 但隨著小枝年齡的增加, 其異速生長常數顯著減小。這一結果表明在2年生、3年生的小枝中也存在單葉質量-出葉強度的負等速權衡。但若忽略小枝年齡, 將前三年的異齡小枝看成同一樣本進行研究, 將無法得出此負等速關系(本研究中, CK組整體斜率為-0.485,P<0.001, 95%置信區間為-0.567到-0.416;OTC組整體斜率為-0.532,P=0.001, 95%置信區間為-0.648到-0.437)。異齡小枝的截距差異使主軸向下漂移, 造成整體斜率的改變:整體斜率的絕對值極顯著小于1.0, 而趨近于0.5。因此, 在驗證出葉強度假說時, 應區分小枝年齡進行研究, 不能用總體代替部分, 也不能把部分簡單地累加起來作為整體, 混用不同年齡的小枝進行研究可能因為截距差異無法得到恒定的負等速關系。本研究結果表明當年生小枝上所發現的單葉質量-出葉強度的負等速權衡關系可推及多年生小枝, 但必須對小枝年齡進行區分。
本研究還可推測:當單葉質量與出葉強度間存在-1.0的比例關系時(即在y=bxa中,a恒為-1.0, 可變形為xy=b時), 截距b的生物學意義即為小枝中單位莖生物量上承載的葉生物量(xy=單葉質量×(葉數量/莖生物量)=葉生物量/莖生物量)。即隨著小枝年齡的增加, 單位莖生物量上承載的葉生物量將減小(表3)。岷江冷杉的2年生與3年生小枝上, 葉大小與數量已基本固定的, 且不存在葉片脫落的情況(岷江冷杉四年及以上枝條才有葉片脫落), 葉生物量可能隨外界環境而產生波動, 但莖生物量在不斷增加(表3); 這必然導致葉生物量/莖生物量不斷減小, 亦為圖1中截距隨年齡增加而減少的原因。
3.3 長期增溫對葉大小-數量權衡關系的影響
當以單葉質量或單葉面積表征葉大小時, 同一年齡小枝下對照組與增溫組的異速生長指數都沒有顯著差異, 即長期增溫并不影響異速生長指數, 這與楊冬梅等[9]、孫俊等[16]對不同海拔植物出葉策略的研究結果相似, 即海拔梯度上的溫度差異未改變單葉質量與出葉強度間的負等速生長關系。對植物性狀相互關聯構成的策略維度進行分析并理解它們之間的關系, 可以解釋為什么特定的性狀組合更優勢[4,35], 然而策略維度并非單獨存在, 單葉質量與出葉強度間的權衡關系可能建立在葉莖生長關系之上: WBE模型[1,12]以分型的分配網絡解釋異速生長現象, Niklas和Enquist[2]在此基礎上推導了種子植物當年生的葉與莖存在等速生長關系; 而當葉生物量與莖生物量間存在1.0的比例關系時(即在y=bxa中,a恒為1.0, 可變形為y/x=b時), 葉生物量與莖生物量間的異速生長常數b代表的生物學意義為小枝中單位莖生物量上承載的葉生物量(y/x=葉生物量/莖生物量), 這與單葉質量-出葉強度的異速生長常數的生物學意義相同。因此, 單葉質量與出葉強度的負生長關系可能是葉生物量與莖生物量生長關系的副產物[9]。如何基于植物功能性狀預測陸地生態系統生產力仍存在許多問題, 而長期增溫下岷江冷杉單葉質量與出葉強度間具有穩定的斜率, 將這一權衡關系納入全球碳循環模型可以更好地模擬生態系統生產力的變化[36]。
確定和理解這一負等速生長關系的主軸在增溫下的變化是另一個重要的生態學問題, 長期增溫對權衡關系的影響體現在當年生小枝的異速生長常數減小, 而二年生、三年生的異速生長常數無差異(表1)。在共同斜率下, OTC中1年生小枝的截距顯著小于對照(表1), 共同主軸向下漂移。這一差異可以理解為相同出葉強度下OTC中1年生的小枝單葉質量更小, 但OTC中1年生的單葉質量顯著大于CK中1年生, 而OTC中1年生的出葉強度顯著小于CK中1年生(表3), 因此在本研究中應解釋為相同單葉質量下增溫處理的當年生小枝出葉強度更低。這與出葉強度優勢[7]存在一定區別。“出葉強度優勢”假說認為高的出葉強度意味著每單位莖體積具有更多的芽, 提供了更大的表型可塑性, 以調整分生組織/芽的分配而適應營養和生殖功能[7,37]。但這忽略了一個問題, 出葉強度高并不意味著葉片數量高。在本研究中, OTC中1年生的出葉強度顯著小于CK中1年生, 而葉片數量卻顯著大于CK中1年生, 原因在于增溫也促進了莖生物量的增加, 并且超過了葉片數量的增加速度(表3)。這一現象符合李俊慧等對展葉效率的研究,即葉數量增加速度沒有小枝莖長度的增加速度快, 長枝相對短枝具有更低的出葉強度[25]。在本研究中, 增溫促進了當年生小枝莖長度的增加(表3), 而長枝在承載相同葉生物量時機械負載更高[38-39]: 由于機械結構的限制, 小枝在重力與風壓脅迫下, 長枝因力臂較長, 承載相同葉質量或相同葉面積時, 莖基部的機械負載將高于短枝[38]。因此, 增溫降低了當年生小枝的出葉強度, 促進了莖長度的增加, 從而使小枝上可以附著更多的葉片, 以獲得更多的適應性收益。此外, 增溫處理下2年生、3年生小枝的異速常數與對照組并無顯著差異, 莖長度與出葉強度也無顯著差異。這可能有兩方面原因, 一方面是小枝長度受到機械結構與發育的限制[38-39]; 另一方面, 可能是溫度的作用隨小枝年齡增加而降低: 當年生小枝更為活躍, 是重要的生長點, 隨著年齡增加小枝生長對溫度的敏感性降低。由此導致不同年齡小枝的出葉策略對長期增溫具有差異性響應。總體而言, 岷江冷杉葉片形態特征因長期增溫而表現出顯著的旱生傾向[23], 但當年生小枝上葉片數量的增加, 彌補了葉大小發育受限的損失, 進而促進植株生長。盡管這一促進作用將隨小枝年齡增加而減弱, 但整個植株仍將積累更多的生物量, 符合Silva等[40]關于野外樹木生長隨氣候變化而加速的結論。溫度增加被認為是樹木加速生長和森林擴張的原因之一[40], 出葉策略的改變可能是其內在機制,如果全球氣候進一步變化, 岷江冷杉將加速生長而在樹線交錯帶擴張。
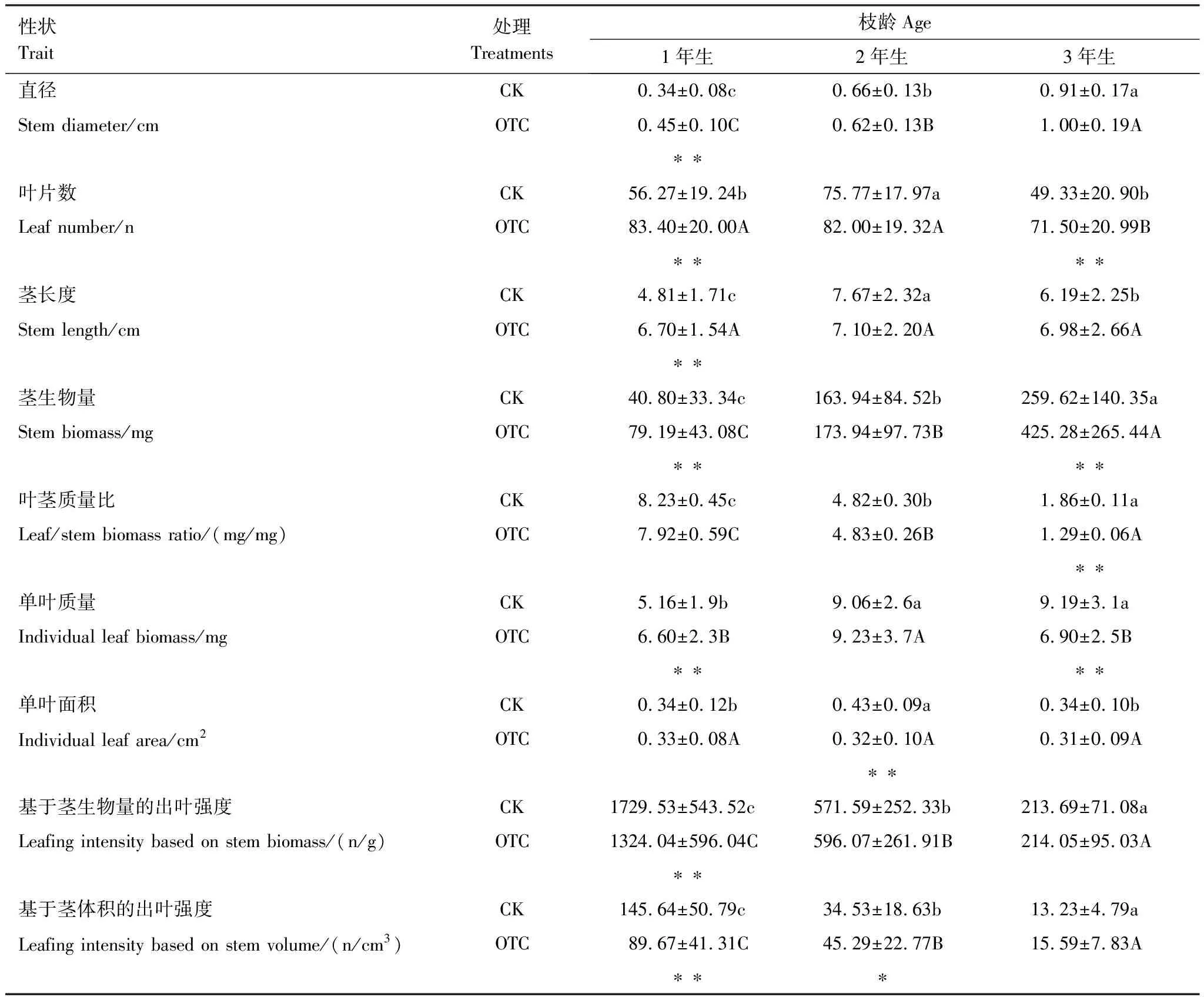
表3 岷江冷杉的莖、葉功能性狀(平均值±標準偏差)
4 結論
使用不同參數表征葉大小與出葉強度得到的結果存在差異; 多年生小枝上存在單葉質量-出葉強度的負等速權衡關系, 共同主軸隨小枝年齡增加而向下漂移; 長期增溫并不影響單葉質量與出葉強度的異速生長指數, 不同年齡小枝的異速生長常數對增溫具有差異性響應。增溫處理的當年生小枝在相同單葉質量下的出葉強度更低, 以換取葉片總數的增加, 使小枝具有更大的可塑性而適應增溫。今后的研究中, 可對策略維度的集合構成的混合策略進行分析, 全面反映植物的生態適應策略。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
